
Opioid and Cannabinoid Systems in Pain: Emerging Molecular Mechanisms and Use in Clinical Practice, Health, and Fitness

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Genesis and Modulation of Nociception in the Nervous System
2.1. Modulation of the Signal in the Central Nervous System
2.2. The Role of Brain Structures
2.3. Descending Pathways, Bidirectional Control, and DNIC
3. Opioid and Cannabinoid Receptors in Pain: Emerging Molecular Mechanisms
3.1. Opioid Receptors
3.2. Cannabinoid Receptors
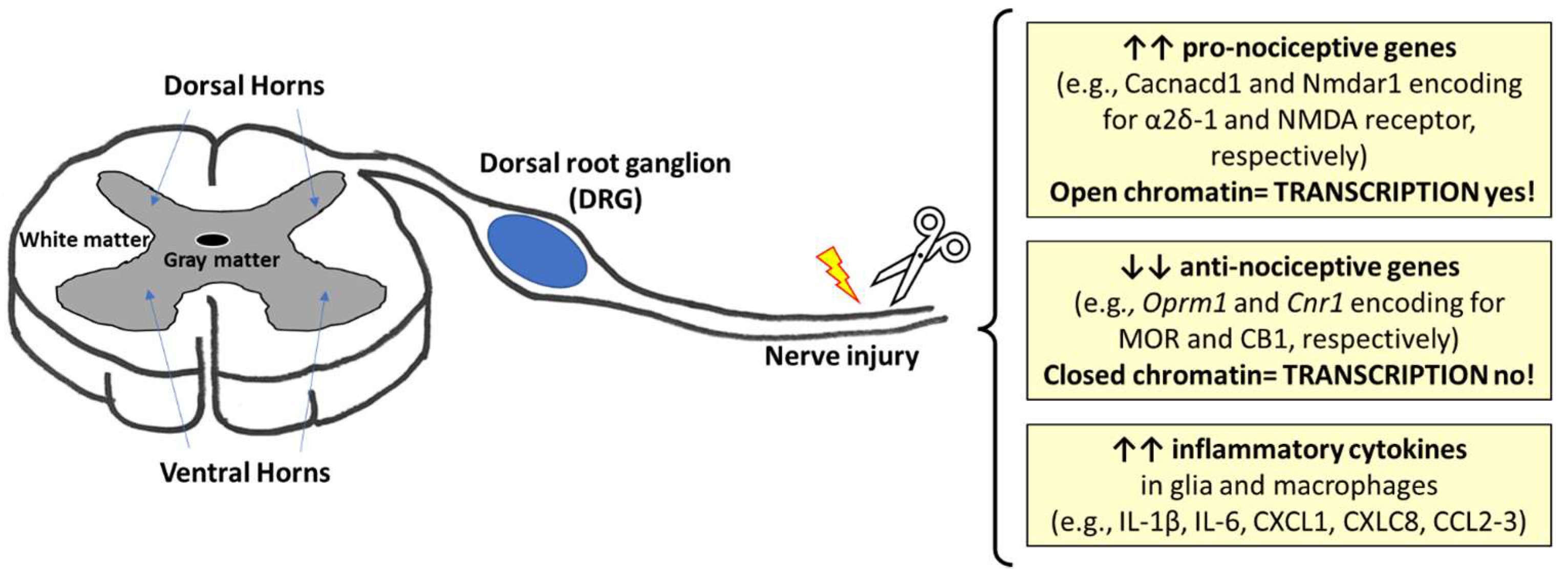
3.3. The Epigenetic Modulation of Genes Encoding for Opioid and Cannabinoid Receptors in Pain
4. Emerging Applications of CBD in Pain Management
4.1. Clinical Applications of CBD in Disease
4.2. CBD in Health and Fitness, and Sport
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- IASP Taxonomy. Available online: http://www.iasp-pain.org/Taxonomy (accessed on 6 January 2024).
- Ossipov, M.H. The Perception and Endogenous Modulation of Pain. Scientifica 2012, 2012, 561761. [Google Scholar] [CrossRef] [PubMed]
- Fields, H.L. Pain: An unpleasant topic. Pain 1999, 82 (Suppl. 6), S61–S69. [Google Scholar] [CrossRef]
- Welch, S.P. Interaction of the cannabinoid and opioid systems in the modulation of nociception. Int. Rev. Psychiatry 2009, 21, 143–151. [Google Scholar] [CrossRef]
- Ochandarena, N.E.; Niehaus, J.K.; Tassou, A.; Scherrer, G. Cell-type specific molecular architecture for mu opioid receptor function in pain and addiction circuits. Neuropharmacology 2023, 238, 109597. [Google Scholar] [CrossRef] [PubMed]
- Adarsh, A.T.; Rahmat, S.; Sangle, P.; Sandhu, O.; Khan, S. Cannabinoid Receptors and Their Relationship with Chronic Pain: A Narrative Review. Cureus 2020, 12, e10436. [Google Scholar] [CrossRef]
- Finn, D.P.; Haroutounian, S.; Hohmann, A.G.; Krane, E.; Soliman, N.; Rice, A.S.C. Cannabinoids, the endocannabinoid system, and pain: A review of preclinical studies. Pain 2021, 162 (Suppl. 1), S5–S25. [Google Scholar] [CrossRef]
- Mercadante, S.; Arcuri, E.; Santoni, A. Opioid-Induced Tolerance and Hyperalgesia. CNS Drugs 2019, 33, 943–955. [Google Scholar] [CrossRef] [PubMed]
- Mazzeo, F.; Meccariello, R.; Guatteo, E. Molecular and Epigenetic Aspects of Opioid Receptors in Drug Addiction and Pain Management in Sport. Int. J. Mol. Sci. 2023, 24, 7831. [Google Scholar] [CrossRef]
- Spanagel, R. Cannabinoids and the endocannabinoid system in reward processing and addiction: From mechanisms to interventions. Dialogues Clin. Neurosci. 2020, 22, 241–250. [Google Scholar] [CrossRef]
- Zvonarev, V.; Fatuki, T.A.; Tregubenko, P. The Public Health Concerns of Marijuana Legalization: An Overview of Current Trends. Cureus 2019, 11, e5806. [Google Scholar] [CrossRef]
- Meccariello, R.; Santoro, A.; D’Angelo, S.; Morrone, R.; Fasano, S.; Viggiano, A.; Pierantoni, R. The Epigenetics of the Endocannabinoid System. Int. J. Mol. Sci. 2020, 21, 1113. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.X.; Malik, A.; Xiong, M.; Bekker, A.; Tao, Y.X. Nerve trauma-caused downregulation of opioid receptors in primary afferent neurons: Molecular mechanisms and potential managements. Exp. Neurol. 2021, 337, 113572. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K.; Pan, H.L. Epigenetic Mechanisms of Neural Plasticity in Chronic Neuropathic Pain. ACS Chem. Neurosci. 2022, 13, 432–441. [Google Scholar] [CrossRef]
- Luz-Veiga, M.; Azevedo-Silva, J.; Fernandes, J.C. Beyond Pain Relief: A Review on Cannabidiol Potential in Medical Therapies. Pharmaceuticals 2023, 16, 155. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, M.R.B.; Joshaghani, N.; Villa, N.; Badla, O.; Goit, R.; Saddik, S.E.; Dawood, S.N.; Rabih, A.M.; Niaj, A.; Raman, A.; et al. Efficacy, Safety, and Regulation of Cannabidiol on Chronic Pain: A Systematic Review. Cureus 2022, 14, e26913. [Google Scholar] [CrossRef]
- McCartney, D.; Benson, M.J.; Desbrow, B.; Irwin, C.; Suraev, A.; McGregor, I.S. Cannabidiol and Sports Performance: A Narrative Review of Relevant Evidence and Recommendations for Future Research. Sports Med. Open 2020, 6, 27. [Google Scholar] [CrossRef]
- Rojas-Valverde, D. Potential Role of Cannabidiol on Sports Recovery: A Narrative Review. Front. Physiol. 2021, 12, 722550. [Google Scholar] [CrossRef]
- Flores, V.A.; Kisiolek, J.N.; Ramani, A.; Townsend, R.; Rodriguez, E.; Butler, B.; Stewart, L.K. Effects of Oral Cannabidiol on Health and Fitness in Healthy Adults: An 8-Week Randomized Trial. Nutrients 2023, 15, 2664. [Google Scholar] [CrossRef]
- Berta, T.; Qadri, Y.; Tan, P.-H.; Ji, R.-R. Targeting dorsal root ganglia and primary sensory neurons for the treatment of chronic pain. Expert Opin. Ther. Targets 2017, 21, 695–703. [Google Scholar] [CrossRef]
- Sexton, J.E.; Cox, J.J.; Zhao, J.; Wood, J.N. The Genetics of Pain: Implications for Therapeutics. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 123–142. [Google Scholar] [CrossRef]
- Dubin, A.E.; Patapoutian, A. Nociceptors: The sensors of the pain pathway. J. Clin. Investig. 2010, 120, 3760–3772. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Bautista, D.M.; Scherrer, G.; Julius, D. Cellular and Molecular Mechanisms of Pain. Cell 2009, 139, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Costigan, M.; Scholz, J.; Woolf, C.J. Neuropathic Pain: A Maladaptive Response of the Nervous System to Damage. Ann. Rev. Neurosci. 2009, 32, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Bruno, O.; Schenone, S.; Ranise, A.; Bondavalli, F.; Filippelli, W.; Falcone, G.; Motola, G.; Mazzeo, F. Antiinflammatory agents: New series of N-substituted amino acids with complex pyrimidine structures endowed with antiphlogistic activity. Farmaco 1999, 54, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, K.; Matsuda, H. Nerve growth factor and wound healing. Prog. Brain Res. 2004, 146, 369–384. [Google Scholar]
- Matsuda, H.; Koyama, H.; Sato, H.; Sawada, J.; Itakura, A.; Tanaka, A.; Matsumoto, M.; Konno, K.; Ushio, H.; Matsuda, K. Role of nerve growth factor in cutaneous wound healing: Accelerating effects in normal and healing-impaired diabetic mice. J. Exp. Med. 1998, 187, 297–306. [Google Scholar] [CrossRef]
- Heumann, R.; Korsching, S.; Bandtlow, C.; Thoenen, H. Changes of nerve growth factor synthesis in nonneuronal cells in response to sciatic nerve transection. J. Cell Biol. 1987, 104, 1623–1631. [Google Scholar] [CrossRef]
- Lewin, G.R.; Mendell, L.M. Nerve growth factor and nociception. Trends Neurosci. 1993, 16, 353–359. [Google Scholar] [CrossRef]
- McMahon, S.B. (Ed.) Wall and Melzack’s Textbook of Pain, 6th ed.; Elsevier/Saunders: Philadelphia, PA, USA, 2013; ISBN 978-0-7020-4059-7. [Google Scholar]
- Testa, G.; Cattaneo, A.; Capsoni, S. Understanding pain perception through genetic painlessness diseases: The role of NGF and proNGF. Pharmacol. Res. 2021, 169, 105662. [Google Scholar] [CrossRef]
- Standring, S. (Ed.) Gray’s Anatomy: The Anatomical Basis of Clinical Practice, 41st ed.; Elsevier Limited: New York, NY, USA, 2016; ISBN 978-0-7020-5230-9. [Google Scholar]
- Iyengar, S.; Ossipov, M.H.; Johnson, K.W. The role of calcitonin gene–related peptide in peripheral and central pain mechanisms including migraine. Pain 2017, 158, 543–559. [Google Scholar] [CrossRef]
- Woolf, C.J.; Ma, Q. Nociceptors--noxious stimulus detectors. Neuron 2007, 55, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Basbaum, A.I.; Julius, D. Toward better pain control. Sci. Am. 2006, 294, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; Morcuende, S.; Webber, M.; Hunt, S.P.; Dickenson, A.H. Superficial NK1-expressing neurons control spinal excitability through activation of descending pathways. Nat. Neurosci. 2003, 5, 1319–1326. [Google Scholar] [CrossRef]
- Latremoliere, A.; Woolf, C.J. Central sensitization: A generator of pain hypersensitivity by central neural plasticity. J. Pain 2009, 10, 895–926. [Google Scholar] [CrossRef] [PubMed]
- Ruscheweyh, R.; Wilder-Smith, O.; Drdla, R.; Liu, X.-G.; Sandkühler, J. Long-term potentiation in spinal nociceptive pathways as a novel target for pain therapy. Mol. Pain 2011, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Beecher, H.K. Pain in Men Wounded in Battle. Ann. Surg. 1946, 123, 96–105. [Google Scholar] [CrossRef]
- Treede, R.D.; Kenshalo, D.R.; Gracely, R.H.; Jones, A.K. The cortical representation of pain. Pain 1999, 79, 105–111. [Google Scholar] [CrossRef]
- Ploner, M.; Freund, H.J.; Schnitzler, A. Pain affect without pain sensation in a patient with a postcentral lesion. Pain 1999, 81, 211–214. [Google Scholar] [CrossRef]
- Foltz, E.L.; White, L.E. Pain ‘relief’ by frontal cingulumotomy. J. Neurosurg. 1962, 19, 89–100. [Google Scholar] [CrossRef]
- Tsou, K.; Jang, C.S. Studies on the site of analgesic action of morphine by intracerebral micro-injection. Sci. Sin. 1964, 13, 1099–1109. [Google Scholar] [CrossRef]
- Pereira, E.A.; Green, A.L.; Bradley, K.M.; Soper, N.; Moir, L.; Stein, J.F.; Aziz, T.Z. Regional cerebral perfusion differences between periventricular grey, thalamic and dual target deep brain stimulation for chronic neuropathic pain. Stereotact. Funct. Neurosurg. 2007, 85, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Heinricher, M.M.; Tavares, I.; Leith, J.L.; Lumb, B.M. Descending control of nociception: Specificity, recruitment and plasticity. Brain Res. Rev. 2009, 60, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Bajic, D.; Proudfit, H.K. Projections of neurons in the periaqueductal gray to pontine and medullary catecholamine cell groups involved in the modulation of nociception. J. Comp. Neurol. 1999, 405, 359–379. [Google Scholar] [CrossRef]
- Odeh, F.; Antal, M. The projections of the midbrain periaqueductal grey to the pons and medulla oblongata in rats. Eur. J. Neurosci. 2001, 14, 1275–1286. [Google Scholar] [CrossRef]
- Osborne, P.B.; Vaughan, C.W.; Wilson, H.I.; Christie, M.J. Opioid inhibition of rat periaqueductal grey neurones with identified projections to rostral ventromedial medulla in vitro. J. Physiol. 1996, 490 Pt 2, 383–389. [Google Scholar] [CrossRef]
- Morgan, M.M.; Whittier, K.L.; Hegarty, D.M.; Aicher, S.A. Periaqueductal gray neurons project to spinally projecting GABAergic neurons in the rostral ventromedial medulla. Pain 2008, 140, 376–386. [Google Scholar] [CrossRef]
- Aicher, S.A.; Hermes, S.M.; Whittier, K.L.; Hegarty, D.M. Descending projections from the rostral ventromedial medulla (RVM) to trigeminal and spinal dorsal horns are morphologically and neurochemically distinct. J. Chem. Neuroanat. 2012, 43, 103–111. [Google Scholar] [CrossRef]
- Le Bars, D.; Dickenson, A.H.; Besson, J.M. Diffuse noxious inhibitory controls (DNIC). I. Effects on dorsal horn convergent neurones in the rat. Pain 1979, 6, 283–304. [Google Scholar] [CrossRef]
- Le Bars, D.; Dickenson, A.H.; Besson, J.M. Diffuse noxious inhibitory controls (DNIC). II. Lack of effect on non-convergent neurones, supraspinal involvement and theoretical implications. Pain 1979, 6, 305–327. [Google Scholar] [CrossRef]
- Villanueva, L.; Le Bars, D. The activation of bulbo-spinal controls by peripheral nociceptive inputs: Diffuse noxious inhibitory controls. Biol. Res. 1995, 28, 113–125. [Google Scholar]
- Lima, D.; Almeida, A. The medullary dorsal reticular nucleus as a pronociceptive centre of the pain control system. Prog. Neurobiol. 2002, 66, 81–108. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.; Tavares, I.; Lima, D.; Coimbra, A. Descending projections from the medullary dorsal reticular nucleus make synaptic contacts with spinal cord lamina I cells projecting to that nucleus: An electron microscopic tracer study in the rat. Neuroscience 1993, 55, 1093–1106. [Google Scholar] [CrossRef]
- Le Bars, D. The whole body receptive field of dorsal horn multireceptive neurones. Brain Res. Brain Res. Rev. 2002, 40, 29–44. [Google Scholar] [CrossRef]
- Jeske, N.A. Dynamic Opioid Receptor Regulation in the Periphery. Mol. Pharmacol. 2019, 95, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Darcq, E.; Kieffer, B.L. Opioid receptors: Drivers to addiction? Nat. Rev. Neurosci. 2018, 19, 499–514. [Google Scholar] [CrossRef]
- Bodnar, R.J. Endogenous Opiates and Behavior: 2015. Peptides 2017, 88, 126–188. [Google Scholar] [PubMed]
- Pfeiffer, A.; Brantl, V.; Herz, A.; Emrich, H.M. Psychotomimesis mediated by kappa opiate receptors. Science 1986, 233, 774–776. [Google Scholar] [CrossRef]
- Filliol, D.; Ghozland, S.; Chluba, J.; Martin, M.; Matthes, H.W.; Simonin, F.; Befort, K.; Gavériaux-Ruff, C.; Dierich, A.; LeMeur, M.; et al. Mice deficient for delta- and mu-opioid receptors exhibit opposing alterations of emotional responses. Nat. Genet. 2000, 25, 195–200. [Google Scholar] [CrossRef]
- Matthes, H.W.; Maldonado, R.; Simonin, F.; Valverde, O.; Slowe, S.; Kitchen, I.; Befort, K.; Dierich, A.; Le Meur, M.; Dollé, P.; et al. Loss of morphine-induced analgesia, reward effect and withdrawal symptoms in mice lacking the mu-opioid-receptor gene. Nature 1996, 383, 819–823. [Google Scholar] [CrossRef]
- Stein, C. New concepts in opioid analgesia. Expert Opin. Investig. Drugs 2018, 27, 765–775. [Google Scholar] [CrossRef]
- Yekkirala, A.S.; Roberson, D.P.; Bean, B.P.; Woolf, C.J. Breaking barriers to novel analgesic drug development. Nat. Rev. Drug Discov. 2017, 16, 545–564. [Google Scholar] [CrossRef] [PubMed]
- Spahn, V.; Del Vecchio, G.; Labuz, D.; Rodriguez-Gaztelumendi, A.; Massaly, N.; Temp, J.; Durmaz, V.; Sabri, P.; Reidelbach, M.; Machelska, H.; et al. A nontoxic pain killer designed by modeling of pathological receptor conformations. Science 2017, 355, 966–969. [Google Scholar] [CrossRef] [PubMed]
- Machelska, H.; Celik, M.Ö. Advances in Achieving Opioid Analgesia without Side Effects. Front. Pharmacol. 2018, 9, 1388. [Google Scholar] [CrossRef]
- Meccariello, R. Molecular Advances on Cannabinoid and Endocannabinoid Research. Int. J. Mol. Sci. 2023, 24, 12760. [Google Scholar] [CrossRef] [PubMed]
- Mackie, K. Cannabinoid receptors: Where they are and what they do. J. Neuroendocrinol. 2008, 20 (Suppl. 1), 10–14. [Google Scholar] [CrossRef]
- Xu, K.; Wu, Y.; Tian, Z.; Xu, Y.; Wu, C.; Wang, Z. Microglial Cannabinoid CB(2) Receptors in Pain Modulation. Int. J. Mol. Sci. 2023, 24, 2348. [Google Scholar] [CrossRef] [PubMed]
- Kleberg, K.; Hassing, H.A.; Hansen, H.S. Classical endocannabinoid-like compounds and their regulation by nutrients. Biofactors 2014, 40, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.C.; Mackie, K. Review of the Endocannabinoid System. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2021, 6, 607–615. [Google Scholar] [CrossRef]
- Oddi, S.; Scipioni, L.; Maccarrone, M. Endocannabinoid system and adult neurogenesis: A focused review. Prog. Lipid Res. 2023, 91, 101239. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J. The biomedical challenge of neurodegenerative disorders: An opportunity for cannabinoid-based therapies to improve on the poor current therapeutic outcomes. Br. J. Pharmacol. 2019, 176, 1370–1383. [Google Scholar] [CrossRef]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Chianese, R.; Coccurello, R.; Viggiano, A.; Scafuro, M.; Fiore, M.; Coppola, G.; Operto, F.F.; Fasano, S.; Laye, S.; Pierantoni, R.; et al. Impact of Dietary Fats on Brain Functions. Curr. Neuropharmacol. 2018, 16, 1059–1085. [Google Scholar] [CrossRef]
- Santoro, A.; Mele, E.; Marino, M.; Viggiano, A.; Nori, S.L.; Meccariello, R. The Complex Interplay between Endocannabinoid System and the Estrogen System in Central Nervous System and Periphery. Int. J. Mol. Sci. 2021, 22, 972. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, R.W.; Oliveira, C.L.; Guimarães, F.S.; Campos, A.C. Cannabinoid signalling in embryonic and adult neurogenesis: Possible implications for psychiatric and neurological disorders. Acta Neuropsychiatr. 2019, 31, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Kumar, U. Cannabinoid Receptors and the Endocannabinoid System: Signaling and Function in the Central Nervous System. Int. J. Mol. Sci. 2018, 19, 833. [Google Scholar] [CrossRef]
- Wang, J.; Lu, H.X.; Wang, J. Cannabinoid receptors in osteoporosis and osteoporotic pain: A narrative update of review. J. Pharm. Pharmacol. 2019, 71, 1469–1474. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, Q.; Chang, C. Epigenetics in Health and Disease. Adv. Exp. Med. Biol. 2020, 1253, 3–55. [Google Scholar] [CrossRef]
- Zhang, G.; Pradhan, S. Mammalian epigenetic mechanisms. IUBMB Life 2014, 66, 240–256. [Google Scholar] [CrossRef]
- Gölzenleuchter, M.; Kanwar, R.; Zaibak, M.; Al Saiegh, F.; Hartung, T.; Klukas, J.; Smalley, R.L.; Cunningham, J.M.; Figueroa, M.E.; Schroth, G.P.; et al. Plasticity of DNA methylation in a nerve injury model of pain. Epigenetics 2015, 10, 200–212. [Google Scholar] [CrossRef]
- Garriga, J.; Laumet, G.; Chen, S.R.; Zhang, Y.; Madzo, J.; Issa, J.J.; Pan, H.L.; Jelinek, J. Nerve Injury-Induced Chronic Pain Is Associated with Persistent DNA Methylation Reprogramming in Dorsal Root Ganglion. J. Neurosci. 2018, 38, 6090–6101. [Google Scholar] [CrossRef]
- Mo, K.; Wu, S.; Gu, X.; Xiong, M.; Cai, W.; Atianjoh, F.E.; Jobe, E.E.; Zhao, X.; Tu, W.F.; Tao, Y.X. MBD1 Contributes to the Genesis of Acute Pain and Neuropathic Pain by Epigenetic Silencing of Oprm1 and Kcna2 Genes in Primary Sensory Neurons. J. Neurosci. 2018, 38, 9883–9899. [Google Scholar] [CrossRef]
- Hwang, C.K.; Wagley, Y.; Law, P.Y.; Wei, L.N.; Loh, H.H. Analysis of epigenetic mechanisms regulating opioid receptor gene transcription. Methods Mol. Biol. 2015, 1230, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Zhou, W.; Wang, Y.; Sun, T.; Wang, H.; Zhang, Z.; Jin, Y. Histone deacetylase inhibitor-induced emergence of synaptic δ-opioid receptors and behavioral antinociception in persistent neuropathic pain. Neuroscience 2016, 339, 54–63. [Google Scholar] [CrossRef]
- Sun, Y.; Sahbaie, P.; Liang, D.; Li, W.; Shi, X.; Kingery, P.; Clark, J.D. DNA Methylation Modulates Nociceptive Sensitization after Incision. PLoS ONE 2015, 10, e0142046. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, S.R.; Laumet, G.; Chen, H.; Pan, H.L. Nerve Injury Diminishes Opioid Analgesia through Lysine Methyltransferase-mediated Transcriptional Repression of μ-Opioid Receptors in Primary Sensory Neurons. J. Biol. Chem. 2016, 291, 8475–8485. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhang, J.; Chen, L.; Chen, S.R.; Chen, H.; Zhang, G.; Pan, H.L. Histone methyltransferase G9a diminishes expression of cannabinoid CB1 receptors in primary sensory neurons in neuropathic pain. J. Biol. Chem. 2020, 295, 3553–3562. [Google Scholar] [CrossRef] [PubMed]
- Viet, C.T.; Dang, D.; Aouizerat, B.E.; Miaskowski, C.; Ye, Y.; Viet, D.T.; Ono, K.; Schmidt, B.L. OPRM1 Methylation Contributes to Opioid Tolerance in Cancer Patients. J. Pain 2017, 18, 1046–1059. [Google Scholar] [CrossRef]
- Meccariello, R.; Bellenchi, G.C.; Pulcrano, S.; D’Addario, S.L.; Tafuri, D.; Mercuri, N.B.; Guatteo, E. Neuronal dysfunction and gene modulation by non-coding RNA in Parkinson’s disease and synucleinopathies. Front. Cell. Neurosci. 2024, 17, 1328269. [Google Scholar] [CrossRef]
- Wu, W.; Ji, X.; Zhao, Y. Emerging Roles of Long Non-coding RNAs in Chronic Neuropathic Pain. Front. Neurosci. 2019, 13, 1097. [Google Scholar] [CrossRef]
- He, Y.; Wang, Z.J. Let-7 microRNAs and Opioid Tolerance. Front. Genet. 2012, 3, 110. [Google Scholar] [CrossRef]
- Chen, Y.; Han, Y.; Liu, L.; Liu, M.; Lin, J.; Tang, Y.; Guo, S.; He, R.; Wu, Q. N6-Methyladenosine methylase METTL3 contributes to neuropathic pain by epigenetic silencing of mu opioid receptor. Behav. Brain Res. 2023, 452, 114592. [Google Scholar] [CrossRef] [PubMed]
- Uchida, H.; Ma, L.; Ueda, H. Epigenetic gene silencing underlies C-fiber dysfunctions in neuropathic pain. J. Neurosci. 2010, 30, 4806–4814. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhao, J.Y.; Gu, X.; Liang, L.; Wu, S.; Mo, K.; Feng, J.; Guo, W.; Zhang, J.; Bekker, A.; et al. Nerve injury-induced epigenetic silencing of opioid receptors controlled by DNMT3a in primary afferent neurons. Pain 2017, 158, 1153–1165. [Google Scholar] [CrossRef]
- Ghosh, K.; Zhang, G.; Chen, H.; Chen, S.R.; Pan, H.L. Cannabinoid CB2 receptors are upregulated via bivalent histone modifications and control primary afferent input to the spinal cord in neuropathic pain. J. Biol. Chem. 2022, 298, 101999. [Google Scholar] [CrossRef]
- Sideris, A.; Doan, L.V. An Overview of Cannabidiol. Anesth. Analg. 2024, 138, 54–68. [Google Scholar] [CrossRef]
- Pellati, F.; Borgonetti, V.; Brighenti, V.; Biagi, M.; Benvenuti, S.; Corsi, L. Cannabis sativa L. and Nonpsychoactive Cannabinoids: Their Chemistry and Role against Oxidative Stress, Inflammation, and Cancer. BioMed Res. Int. 2018, 2018, 1691428. [Google Scholar] [CrossRef] [PubMed]
- WHO. Cannabidiol (CBD) Critical Review Report 2018. Available online: https://cdn.who.int/media/docs/default-source/controlled-substances/whocbdreportmay2018-2.pdf?sfvrsn=f78db177_2 (accessed on 22 September 2019).
- Larsen, C.; Shahinas, J. Dosage, efficacy and safety of cannabidiol administration in adults: A systematic review of human trials. J. Clin. Med. Res. 2020, 12, 129–141. [Google Scholar] [CrossRef]
- Krcevski-Skvarc, N.; Wells, C.; Häuser, W. Availability and approval of cannabis-based medicines for chronic pain management and palliative/supportive care in Europe: A survey of the status in the chapters of the European Pain Federation. Eur. J. Pain 2018, 22, 440–454. [Google Scholar] [CrossRef]
- Maldonado, R.; Baños, J.E.; Cabañero, D. The endocannabinoid system and neuropathic pain. Pain 2016, 157 (Suppl. 1), S23–S32. [Google Scholar] [CrossRef]
- Mohammed, S.Y.M.; Leis, K.; Mercado, R.E.; Castillo, M.M.S.; Miranda, K.J.; Carandang, R.R. Effectiveness of Cannabidiol to Manage Chronic Pain: A Systematic Review. Pain Manag. Nurs. 2024, 25, e76–e86. [Google Scholar] [CrossRef]
- Hameed, M.; Prasad, S.; Jain, E.; Dogrul, B.N.; Al-Oleimat, A.; Pokhrel, B.; Chowdhury, S.; Co, E.L.; Mitra, S.; Quinonez, J.; et al. Medical Cannabis for Chronic Nonmalignant Pain Management. Curr. Pain Headache Rep. 2023, 27, 57–63. [Google Scholar] [CrossRef]
- Capano, A.; Weaver, R.; Burkman, E. Evaluation of the effects of CBD hemp extract on opioid use and quality of life indicators in chronic pain patients: A prospective cohort study. Postgrad. Med. 2020, 132, 56–61. [Google Scholar] [CrossRef]
- Aviram, J.; Samuelly-Leichtag, G. Efficacy of Cannabis-Based Medicines for Pain Management: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Pain Physician 2017, 20, E755–E796. [Google Scholar] [CrossRef]
- Mücke, M.; Phillips, T.; Radbruch, L.; Petzke, F.; Häuser, W. Cannabis-based medicines for chronic neuropathic pain in adults. Cochrane Database Syst. Rev. 2018, 3, CD012182. [Google Scholar] [CrossRef] [PubMed]
- Petzke, F.; Tölle, T.; Fitzcharles, M.A.; Häuser, W. Cannabis-Based Medicines and Medical Cannabis for Chronic Neuropathic Pain. CNS Drugs 2022, 36, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Boehnke, K.F.; Häuser, W.; Fitzcharles, M.A. Cannabidiol (CBD) in Rheumatic Diseases (Musculoskeletal Pain). Curr. Rheumatol. Rep. 2022, 24, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Frane, N.; Stapleton, E.; Iturriaga, C.; Ganz, M.; Rasquinha, V.; Duarte, R. Cannabidiol as a treatment for arthritis and joint pain: An exploratory cross-sectional study. J. Cannabis Res. 2022, 4, 47. [Google Scholar] [CrossRef]
- Mistry, M.; Simpson, P.; Morris, E.; Fritz, A.K.; Karavadra, B.; Lennox, C.; Prosser-Snelling, E. Cannabidiol for the Management of Endometriosis and Chronic Pelvic Pain. J. Minim. Invasive Gynecol. 2022, 29, 169–176. [Google Scholar] [CrossRef]
- Chandwani, B.; Bradley, B.A.; Pace, A.; Buse, D.C.; Singh, R.; Kuruvilla, D. The Exploration of Cannabis and Cannabinoid Therapies for Migraine. Curr. Pain Headache Rep. 2023, 27, 339–350. [Google Scholar] [CrossRef]
- Fiani, B.; Sarhadi, K.J.; Soula, M.; Zafar, A.; Quadri, S.A. Current application of cannabidiol (CBD) in the management and treatment of neurological disorders. Neurol. Sci. 2020, 41, 3085–3098. [Google Scholar] [CrossRef]
- Chen, L.; Sun, Y.; Li, J.; Liu, S.; Ding, H.; Wang, G.; Li, X. Assessing Cannabidiol as a Therapeutic Agent for Preventing and Alleviating Alzheimer’s Disease Neurodegeneration. Cells 2023, 12, 2672. [Google Scholar] [CrossRef]
- Kapoor, G.; Chauhan, P.; Singh, G.; Malhotra, N.; Chahal, A. Physical Activity for Health and Fitness: Past, Present and Future. J. Lifestyle Med. 2022, 12, 9–14. [Google Scholar] [CrossRef]
- Lesnak, J.B.; Sluka, K.A. Mechanism of exercise-induced analgesia: What we can learn from physically active animals. Pain Rep. 2020, 5, e850. [Google Scholar] [CrossRef] [PubMed]
- Harber, V.J.; Sutton, J.R. Endorphins and exercise. Sports Med. 1984, 1, 154–171. [Google Scholar] [CrossRef] [PubMed]
- Docherty, S.; Harley, R.; McAuley, J.J.; Crowe, L.A.N.; Pedret, C.; Kirwan, P.D.; Siebert, S.; Millar, N.L. The effect of exercise on cytokines: Implications for musculoskeletal health: A narrative review. BMC Sports Sci. Med. Rehabil. 2022, 14, 5. [Google Scholar] [CrossRef]
- Hearon, B.A.; Calkins, A.W.; Halperin, D.M.; McHugh, R.K.; Murray, H.W.; Otto, M.W. Anxiety sensitivity and illicit sedative use among opiate-dependent women and men. Am. J. Drug Alcohol. Abus. 2011, 37, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Mazzeo, F.; Santamaria, S.; Montesano, P. Gender difference, nutritional supplements and drug use in sport to enhancing performance: An Italian revision over the last decade. Sport Mont. 2019, 17, 69–73. [Google Scholar] [CrossRef]
- Mazzeo, F.; Volpe, R.A. From gene doping to athlete biological passport. Sport Sci. 2016, 9, 97–103. [Google Scholar]
- Pergolizzi, J.V., Jr.; Raffa, R.B.; Taylor, R., Jr.; Rodriguez, G.; Nalamachu, S.; Langley, P. A review of duloxetine 60 mg once-daily dosing for the management of diabetic peripheral neuropathic pain, fibromyalgia, and chronic musculoskeletal pain due to chronic osteoarthritis pain and low back pain. Pain Pract. 2013, 13, 239–252. [Google Scholar] [CrossRef]
- Mazzeo, F.; Raiola, G. An investigation of drugs abuse in sport performance. J. Hum. Sport Exercise 2018, 13, 309–319. [Google Scholar]
- Nijs, J.; Kosek, E.; Van Oosterwijck, J.; Meeus, M. Dysfunctional endogenous analgesia during exercise in patients with chronic pain: To exercise or not to exercise? Pain Physician 2012, 15 (Suppl. 3), ES205–ES213. [Google Scholar] [CrossRef]
- Mazzeo, F. Anabolic steroid use in sports and in physical activity: Overview and analysis. Sport Mont. 2018, 16, 113–118. [Google Scholar] [CrossRef]
- Peters, E.N.; Yardley, H.; Harrison, A.; Eglit, G.M.L.; Antonio, J.; Turcotte, C.; Bonn-Miller, M.O. A randomized, double-blind, placebo-controlled, repeated-dose pilot study of the safety, tolerability, and preliminary effects of a cannabidiol (CBD)- and cannabigerol (CBG)-based beverage powder to support recovery from delayed onset muscle soreness (DOMS). J. Int. Soc. Sports Nutr. 2023, 20, 2280113. [Google Scholar] [CrossRef]
- Cochrane-Snyman, K.C.; Cruz, C.; Morales, J.; Coles, M. The effects of cannabidiol oil on noninvasive measures of muscle damage in men. Med. Sci. Sport Exerc. 2021, 53, 1460–1472. [Google Scholar] [CrossRef]
- Motola, G.; Russo, F.; Mazzeo, F.; Rinaldi, B.; Capuano, A.; Rossi, F.; Filippelli, A. Over-the-counter oral nonsteroidal anti-inflammatory drugs: A pharmacoepidemiologic study in southern Italy. Adv. Ther. 2001, 18, 216–222. [Google Scholar] [CrossRef]
- Antunes, F.J.R. Musculoskeletal traumatic pain. DOLOR 2022, 37, 86–90. [Google Scholar]
- Marques Azzini, G.O.; Marques Azzini, V.O.; Santos, G.S.; Visoni, S.; Fusco, M.A.; Beker, N.S.; Mahmood, A.; Bizinotto Lana, J.V.; Jeyaraman, M.; Nallakumarasamy, A.; et al. Cannabidiol for musculoskeletal regenerative medicine. Exp. Biol. Med. 2023, 248, 445–455. [Google Scholar] [CrossRef]
- Isenmann, E.; Veit, S.; Starke, L.; Flenker, U.; Diel, P. Effects of Cannabidiol Supplementation on Skeletal Muscle Regeneration after Intensive Resistance Training. Nutrients 2021, 13, 3028. [Google Scholar] [CrossRef] [PubMed]
- Burr, J.F.; Christian, P.; Cheung, C.P.; Kasper, A.M.; Gillham, S.H.; Close, G.L. Cannabis and Athletic Performance. Sports Med. 2021, 51 (Suppl. 1), S75–S87. [Google Scholar] [CrossRef]
- Aguiar, A.S., Jr. Cannabis Is Not Doping. Cannabis Cannabinoid Res. 2023, 8, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Ross, B.; Trojian, T.; Cushman, D.M. Physician Perceptions of Cannabidiol (CBD) and Cannabis in Sports Medicine and Performance. Transl. Sports Med. 2023, 2023, 8824466. [Google Scholar] [CrossRef] [PubMed]
- Mazzeo, F. Attitude and practice of substance misuse and dietary supplements to improve performance in sport. J. Sub. Use 2019, 24, 581–586. [Google Scholar] [CrossRef]
- Hitchcock, L.N.; Tracy, B.L.; Bryan, A.D.; Hutchison, K.E.; Bidwell, L.C. Acute Effects of Cannabis Concentrate on Motor Control and Speed: Smartphone-Based Mobile Assessment. Front. Psychiatry 2021, 11, 623672. [Google Scholar] [CrossRef]
- Boyaji, S.; Merkow, J.; Elman, R.N.M.; Kaye, A.D.; Yong, R.J.; Urman, R.D. The Role of Cannabidiol (CBD) in Chronic Pain Management: An Assessment of Current Evidence. Curr. Pain Headache Rep. 2020, 24, 4. [Google Scholar] [CrossRef]
- Britch, S.C.; Babalonis, S.; Walsh, S.L. Cannabidiol: Pharmacology and therapeutic targets. Psychopharmacology 2021, 238, 9–28. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Gene | Effects on Transcription | Epigenetic Modification | Reference |
|---|---|---|---|
| Oprm1 | ↓ | ↑ H3K9me2 ↑ H3K27me3 ↓ H3K4me3 | [88] |
| Oprm1 | ↓ | H3/H4 hypoacethylation | [95] |
| Oprm1 | ↓ | ↑ promoter DNA methylation via MBD1-dependent recruitment of DNMT3a | [84] |
| Oprm1 and Oprk1 | ↓ | ↑DNA methylation via MBD1-dependent recruitment of DNMT3a for Oprm1 | [96] |
| Cnr1 | ↓ | ↑ H3K9me2 at promoter region | [89] |
| Cnr2 | ↑ | ↑ H3K4me3 ↑ H3K9ac ↓ H3K9me2 ↓ H3K27me3 No effect on DNA methylation | [97] |
| Feature | Opioids | Cannabinoids |
|---|---|---|
| Main effects | Pain relief | Varies depending on the specific cannabinoid (THC can be psychoactive, CBD is not) |
| Mechanism of action | Binds to opioid receptors in the central nervous system and body | Binds to cannabinoid receptors in the brain and body |
| Risks | Addiction, overdose, death | Impaired coordination and reaction time, potential for dependence |
| Use in sports | Pain management | Pain management, inflammation, anxiety (though prohibited by most anti-doping agencies) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Secondulfo, C.; Mazzeo, F.; Pastorino, G.M.G.; Vicidomini, A.; Meccariello, R.; Operto, F.F. Opioid and Cannabinoid Systems in Pain: Emerging Molecular Mechanisms and Use in Clinical Practice, Health, and Fitness. Int. J. Mol. Sci. 2024, 25, 9407. https://doi.org/10.3390/ijms25179407
Secondulfo C, Mazzeo F, Pastorino GMG, Vicidomini A, Meccariello R, Operto FF. Opioid and Cannabinoid Systems in Pain: Emerging Molecular Mechanisms and Use in Clinical Practice, Health, and Fitness. International Journal of Molecular Sciences. 2024; 25(17):9407. https://doi.org/10.3390/ijms25179407
Chicago/Turabian StyleSecondulfo, Carmine, Filomena Mazzeo, Grazia Maria Giovanna Pastorino, Antonella Vicidomini, Rosaria Meccariello, and Francesca Felicia Operto. 2024. "Opioid and Cannabinoid Systems in Pain: Emerging Molecular Mechanisms and Use in Clinical Practice, Health, and Fitness" International Journal of Molecular Sciences 25, no. 17: 9407. https://doi.org/10.3390/ijms25179407
APA StyleSecondulfo, C., Mazzeo, F., Pastorino, G. M. G., Vicidomini, A., Meccariello, R., & Operto, F. F. (2024). Opioid and Cannabinoid Systems in Pain: Emerging Molecular Mechanisms and Use in Clinical Practice, Health, and Fitness. International Journal of Molecular Sciences, 25(17), 9407. https://doi.org/10.3390/ijms25179407