

Specific microRNA Profile Associated with Inflammation and Lipid Metabolism for Stratifying Allergic Asthma Severity
 , , , , ,
, , , , ,  , ,
, , 
Abstract

1. Introduction
2. Results
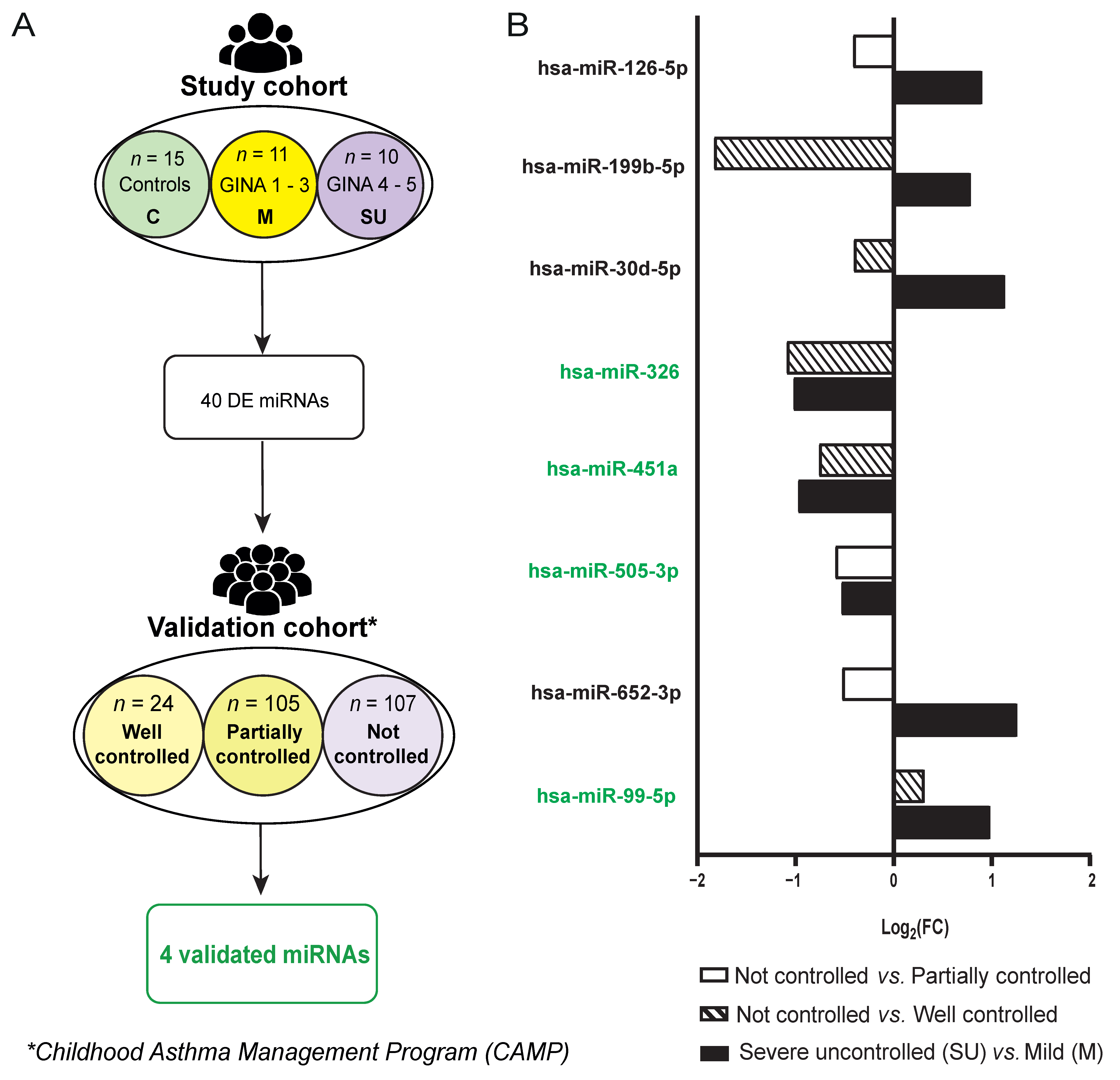
2.1. Subject Classification in Study Cohort
2.2. Severe Uncontrolled Allergic Asthmatic Patients Display a Unique miRNA Fingerprint in Serum
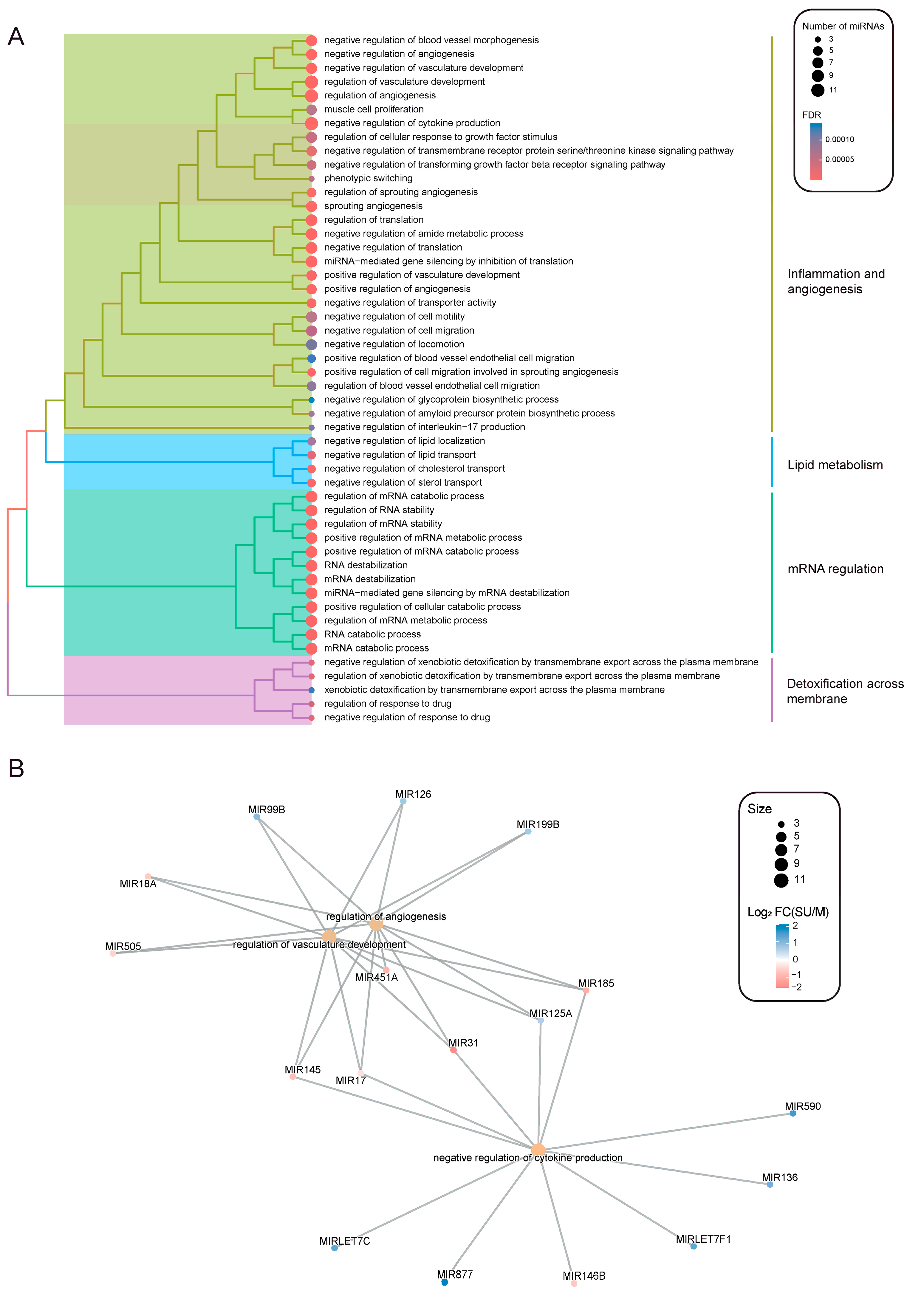
2.3. Inflammation, Angiogenesis and Lipid Metabolism Signatures Are Enriched in Severe Allergic Asthmatic Patients
2.4. 4 Severe Allergic Asthma miRNA Profile Correlates with Dysregulated Lipid Metabolism
2.5. DE miRNAs Can Accurately Classify Allergic Asthmatic Patients According to Their Severity
2.6. Validation of miRNAs as Biomarkers for Severity Degree in Allergic Asthma
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. miRNAs Extraction and Expression Quantification
4.3. Statistics
4.4. Functional Enrichment Analysis
4.5. Random Forest-Based Classifier
4.6. miRNA Validation Analysis in CAMP (Childhood Asthma Management Program) Cohort
4.7. Data Analysis of miRNA Validation in CAMP Cohort
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Villaseñor, A.; Rosace, D.; Obeso, D.; Pérez-Gordo, M.; Chivato, T.; Barbas, C.; Barber, D.; Escribese, M.M. Allergic Asthma: An Overview of Metabolomic Strategies Leading to the Identification of Biomarkers in the Field. Clin. Exp. Allergy 2017, 47, 442–456. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Asthma. Available online: https://www.who.int/news-room/fact-sheets/detail/asthma#:~:text=Asthma%20affected%20an%20estimated%20262,help%20to%20reduce%20asthma%20symptoms (accessed on 7 August 2024).
- Wang, Z.; Li, Y.; Gao, Y.; Fu, Y.; Lin, J.; Lei, X.; Zheng, J.; Jiang, M. Global, Regional, and National Burden of Asthma and Its Attributable Risk Factors from 1990 to 2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Respir. Res. 2023, 24, 169. [Google Scholar] [CrossRef]
- Loureiro, C.C.; Amaral, L.; Ferreira, J.A.; Lima, R.; Pardal, C.; Fernandes, I.; Semedo, L.; Arrobas, A. Omalizumab for Severe Asthma: Beyond Allergic Asthma. BioMed Res. Int. 2018, 2018, 3254094. [Google Scholar] [CrossRef]
- Schatz, M.; Rosenwasser, L. The Allergic Asthma Phenotype. J. Allergy Clin. Immunol. Pract. 2014, 2, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-Y. The Innate Immune Response in House Dust Mite-Induced Allergic Inflammation. Allergy Asthma Immunol. Res. 2013, 5, 68. [Google Scholar] [CrossRef]
- Akar-Ghibril, N.; Casale, T.; Custovic, A.; Phipatanakul, W. Allergic Endotypes and Phenotypes of Asthma. J. Allergy Clin. Immunol. Pract. 2020, 8, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Global Initiative for Asthma. Global Strategy for Asthma Management and Prevention. Available online: www.ginasthma.org (accessed on 17 April 2023).
- Global Initiative for Asthma. Difficult-To-Treat & Severe Asthma in Adolescent and Adult Patients. Available online: https://ginasthma.org/wp-content/uploads/2023/09/GINA-Severe-Asthma-Guide-2023-WEB-WMS.pdf (accessed on 26 October 2023).
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. MiRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef] [PubMed]
- Specjalski, K.; Jassem, E. MicroRNAs: Potential Biomarkers and Targets of Therapy in Allergic Diseases? Arch. Immunol. Ther. Exp. 2019, 67, 213–223. [Google Scholar] [CrossRef]
- Specjalski, K.; Niedoszytko, M. MicroRNAs: Future Biomarkers and Targets of Therapy in Asthma? Curr. Opin. Pulm. Med. 2020, 26, 285–292. [Google Scholar] [CrossRef]
- Weidner, J.; Bartel, S.; Kılıç, A.; Zissler, U.M.; Renz, H.; Schwarze, J.; Schmidt-Weber, C.B.; Maes, T.; Rebane, A.; Krauss-Etschmann, S.; et al. Spotlight on MicroRNAs in Allergy and Asthma. Allergy 2021, 76, 1661–1678. [Google Scholar] [CrossRef]
- Rodrigo-Muñoz, J.M.; Cañas, J.A.; Sastre, B.; Rego, N.; Greif, G.; Rial, M.; Mínguez, P.; Mahíllo-Fernández, I.; Fernández-Nieto, M.; Mora, I.; et al. Asthma Diagnosis Using Integrated Analysis of Eosinophil MicroRNAs. Allergy 2019, 74, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An Overview of MicroRNAs: Biology, Functions, Therapeutics, and Analysis Methods. J. Cell Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.; Chen, Y.; Huang, X.; Huang, Q.; Li, M.; Tan, X. MiR-19 Targets PTEN and Mediates High Mobility Group Protein B1(HMGB1)-Induced Proliferation and Migration of Human Airway Smooth Muscle Cells. PLoS ONE 2019, 14, e0219081. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lang, X.; Xia, S. Elevated Expression of MicroRNA-378 in Children with Asthma Aggravates Airway Remodeling by Promoting the Proliferation and Apoptosis Resistance of Airway Smooth Muscle Cells. Exp. Ther. Med. 2019, 17, 1529–1536. [Google Scholar] [CrossRef]
- Huo, X.; Zhang, K.; Yi, L.; Mo, Y.; Liang, Y.; Zhao, J.; Zhang, Z.; Xu, Y.; Zhen, G. Decreased Epithelial and Plasma MiR-181b-5p Expression Associates with Airway Eosinophilic Inflammation in Asthma. Clin. Exp. Allergy 2016, 46, 1281–1290. [Google Scholar] [CrossRef]
- Milger, K.; Götschke, J.; Krause, L.; Nathan, P.; Alessandrini, F.; Tufman, A.; Fischer, R.; Bartel, S.; Theis, F.J.; Behr, J.; et al. Identification of a Plasma MiRNA Biomarker Signature for Allergic Asthma: A Translational Approach. Allergy 2017, 72, 1962–1971. [Google Scholar] [CrossRef]
- Delgado-Dolset, M.I.; Obeso, D.; Rodríguez-Coira, J.; Tarin, C.; Tan, G.; Cumplido, J.A.; Cabrera, A.; Angulo, S.; Barbas, C.; Sokolowska, M.; et al. Understanding Uncontrolled Severe Allergic Asthma by Integration of Omic and Clinical Data. Allergy 2022, 77, 1772–1785. [Google Scholar] [CrossRef]
- Levänen, B.; Bhakta, N.R.; Torregrosa Paredes, P.; Barbeau, R.; Hiltbrunner, S.; Pollack, J.L.; Sköld, C.M.; Svartengren, M.; Grunewald, J.; Gabrielsson, S.; et al. Altered MicroRNA Profiles in Bronchoalveolar Lavage Fluid Exosomes in Asthmatic Patients. J. Allergy Clin. Immunol. 2013, 131, 894–903.e8. [Google Scholar] [CrossRef]
- Narisety, S.D.; Frischmeyer-Guerrerio, P.A.; Keet, C.A.; Gorelik, M.; Schroeder, J.; Hamilton, R.G.; Wood, R.A. A Randomized, Double-Blind, Placebo-Controlled Pilot Study of Sublingual versus Oral Immunotherapy for the Treatment of Peanut Allergy. J. Allergy Clin. Immunol. 2015, 135, 1275–1282.e6. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X. MiRDB: An Online Database for Prediction of Functional MicroRNA Targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef]
- Childhood Asthma Management Program Research Group; Szefler, S.; Weiss, S.; Tonascia, J.; Adkinson, N.F.; Bender, B.; Cherniack, R.; Donithan, M.; Kelly, H.W.; Reisman, J.; et al. Long-Term Effects of Budesonide or Nedocromil in Children with Asthma. N. Engl. J. Med. 2000, 343, 1054–1063. [Google Scholar] [CrossRef]
- Childhood Asthma Management Program Research Group. The Childhood Asthma Management Program (CAMP): Design, Rationale, and Methods. Control Clin. Trials 1999, 20, 91–120. [Google Scholar] [CrossRef]
- Ortea, I.; Rodríguez-Ariza, A.; Chicano-Gálvez, E.; Arenas Vacas, M.S.; Jurado Gámez, B. Discovery of Potential Protein Biomarkers of Lung Adenocarcinoma in Bronchoalveolar Lavage Fluid by SWATH MS Data-Independent Acquisition and Targeted Data Extraction. J. Proteom. 2016, 138, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Kyyaly, M.A.; Vorobeva, E.V.; Kothalawala, D.M.; Fong, W.C.G.; He, P.; Sones, C.L.; Al-Zahrani, M.; Sanchez-Elsner, T.; Arshad, S.H.; Kurukulaaratchy, R.J. MicroRNAs—A Promising Tool for Asthma Diagnosis and Severity Assessment: A Systematic Review. J. Pers. Med. 2022, 12, 543. [Google Scholar] [CrossRef]
- Gomez, J.L.; Chen, A.; Diaz, M.P.; Zirn, N.; Gupta, A.; Britto, C.; Sauler, M.; Yan, X.; Stewart, E.; Santerian, K.; et al. A Network of Sputum MicroRNAs Is Associated with Neutrophilic Airway Inflammation in Asthma. Am. J. Respir. Crit. Care Med. 2020, 202, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Piao, L.-Z.; Liu, L.; Zhang, X.-F. DNA Methylation and Gene Expression Profiles to Identify Childhood Atopic Asthma Associated Genes. BMC Pulm. Med. 2021, 21, 292. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.R.; Sutcliffe, A.; Kaur, D.; Gupta, S.; Desai, D.; Saunders, R.; Brightling, C.E. CCL2 Release by Airway Smooth Muscle Is Increased in Asthma and Promotes Fibrocyte Migration. Allergy 2014, 69, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo-Muñoz, J.M.; Cañas, J.A.; Sastre, B.; Gil-Martínez, M.; García Latorre, R.; Sastre, J.; Pozo, V. Role of MiR-185-5p as Modulator of Periostin Synthesis and Smooth Muscle Contraction in Asthma. J. Cell Physiol. 2022, 237, 1498–1508. [Google Scholar] [CrossRef]
- Izuhara, K.; Ohta, S.; Ono, J. Using Periostin as a Biomarker in the Treatment of Asthma. Allergy Asthma Immunol. Res. 2016, 8, 491. [Google Scholar] [CrossRef]
- Licari, A.; Brambilla, I.; Sacchi, L.; Marseglia, G.; Ciprandi, G. Periostin, Type 2 Biomarker, Is Not Associated with Asthma Control Grade in Asthmatic Allergic Children. Respir. Med. 2019, 151, 118–120. [Google Scholar] [CrossRef]
- Haj-Salem, I.; Fakhfakh, R.; Bérubé, J.-C.; Jacques, E.; Plante, S.; Simard, M.J.; Bossé, Y.; Chakir, J. MicroRNA-19a Enhances Proliferation of Bronchial Epithelial Cells by Targeting TGF β R2 Gene in Severe Asthma. Allergy 2015, 70, 212–219. [Google Scholar] [CrossRef]
- Rijavec, M.; Korošec, P.; Žavbi, M.; Kern, I.; Malovrh, M.M. Let-7a Is Differentially Expressed in Bronchial Biopsies of Patients with Severe Asthma. Sci. Rep. 2014, 4, 6103. [Google Scholar] [CrossRef] [PubMed]
- Sturgill, J.L. Sphingolipids and Their Enigmatic Role in Asthma. Adv. Biol. Regul. 2018, 70, 74–81. [Google Scholar] [CrossRef] [PubMed]
- McGeachie, M.J.; Dahlin, A.; Qiu, W.; Croteau-Chonka, D.C.; Savage, J.; Wu, A.C.; Wan, E.S.; Sordillo, J.E.; Al-Garawi, A.; Martinez, F.D.; et al. The Metabolomics of Asthma Control: A Promising Link between Genetics and Disease. Immun. Inflamm. Dis. 2015, 3, 224–238. [Google Scholar] [CrossRef] [PubMed]
- Reinke, S.N.; Gallart-Ayala, H.; Gómez, C.; Checa, A.; Fauland, A.; Naz, S.; Kamleh, M.A.; Djukanović, R.; Hinks, T.S.C.; Wheelock, C.E. Metabolomics Analysis Identifies Different Metabotypes of Asthma Severity. Eur. Respir. J. 2017, 49, 1601740. [Google Scholar] [CrossRef] [PubMed]
- Perzanowski, M.S.; Ono, J.G.; Acosta, L.M.; Kim, B.I.; Divjan, A.; Miller, R.; Rundle, A.; Worgall, S.; Worgall, T.S. Distinct Serum Sphingolipid Profiles among School-Aged Children with Exercise-Induced Wheeze and Asthma Persistence. Am. J. Respir. Crit. Care Med. 2017, 195, 1068–1070. [Google Scholar] [CrossRef]
- Trinh, H.K.T.; Kim, S.-C.; Cho, K.; Kim, S.-J.; Ban, G.-Y.; Yoo, H.-J.; Cho, J.-Y.; Park, H.-S.; Kim, S.-H. Exploration of the Sphingolipid Metabolite, Sphingosine-1-Phosphate and Sphingosine, as Novel Biomarkers for Aspirin-Exacerbated Respiratory Disease. Sci. Rep. 2016, 6, 36599. [Google Scholar] [CrossRef]
- Duan, X.-J.; Zhang, X.; Li, L.-R.; Zhang, J.-Y.; Chen, Y.-P. MiR-200a and MiR-200b Restrain Inflammation by Targeting ORMDL3 to Regulate the ERK/MMP-9 Pathway in Asthma. Exp. Lung Res. 2020, 46, 321–331. [Google Scholar] [CrossRef]
- Luthers, C.R.; Dunn, T.M.; Snow, A.L. ORMDL3 and Asthma: Linking Sphingolipid Regulation to Altered T Cell Function. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Miles, E.A.; Childs, C.E.; Calder, P.C. Long-Chain Polyunsaturated Fatty Acids (LCPUFAs) and the Developing Immune System: A Narrative Review. Nutrients 2021, 13, 247. [Google Scholar] [CrossRef]
- Gomez-Casado, C.; Villaseñor, A.; Rodriguez-Nogales, A.; Bueno, J.; Barber, D.; Escribese, M. Understanding Platelets in Infectious and Allergic Lung Diseases. Int. J. Mol. Sci. 2019, 20, 1730. [Google Scholar] [CrossRef] [PubMed]
- D’haene, B.; Mestdagh, P.; Hellemans, J.; Vandesompele, J. MiRNA Expression Profiling: From Reference Genes to Global Mean Normalization. Methods Mol. Biol. 2012, 822, 261–272. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 21 August 2024).
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. ClusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. ClusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Pagès, H.; Carlson, M.; Falcon, S.; Li, N. AnnotationDbi: Manipulation of SQLite-based annotations in Bioconductor. Available online: https://bioconductor.org/packages/AnnotationDbi (accessed on 21 August 2024).
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Li, S.; Xia, J. MetaboAnalystR 3.0: Toward an Optimized Workflow for Global Metabolomics. Metabolites 2020, 10, 186. [Google Scholar] [CrossRef]
- Li, J.; Panganiban, R.; Kho, A.T.; McGeachie, M.J.; Farnam, L.; Chase, R.P.; Weiss, S.T.; Lu, Q.; Tantisira, K.G. Circulating MicroRNAs and Treatment Response in Childhood Asthma. Am. J. Respir. Crit. Care Med. 2020, 202, 65–72. [Google Scholar] [CrossRef]
- Gu, Z. Complex heatmap visualization. iMeta 2022, 1. [Google Scholar] [CrossRef]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Kassambara, A. ggcorrplot: Visualization of a Correlation Matrix Using ‘ggplot2’. 2023. Available online: https://CRAN.R-project.org/package=ggcorrplot (accessed on 21 August 2024).
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. Circlize implements and enhances circular visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Coira, J.; Dolores-Dolset, M.; Obeso, D.; Dolores-Hernández, M.; Quintás, G.; Angulo, S.; Barber, D.; Carrillo, T.; Escribese, M.M.; Villaseñor, A. Troubleshooting in Large-Scale LC-ToF-MS Metabolomics Analysis: Solving Complex Issues in Big Cohorts. Metabolites 2019, 9, 247. [Google Scholar] [CrossRef]
- Gil-de-la-Fuente, A.; Godzien, J.; Saugar, S.; Garcia-Carmona, R.; Badran, H.; Wishart, D.S.; Barbas, C.; Otero, A. CEU Mass Mediator 3.0: A Metabolite Annotation Tool. J. Proteome Res. 2019, 18, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Oestvang, J.; Anthonsen, M.W.; Johansen, B. LysoPC and PAF Trigger Arachidonic Acid Release by Divergent Signaling Mechanisms in Monocytes. J. Lipids 2011, 2011, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sokolowska, M.; Chen, L.-Y.; Liu, Y.; Martinez-Anton, A.; Logun, C.; Alsaaty, S.; Cuento, R.A.; Cai, R.; Sun, J.; Quehenberger, O.; et al. Dysregulation of lipidomic profile and antiviral immunity in response to hyaluronan in patients with severe asthma. J. Allergy Clin. Immunol. 2017, 139, 1379–1383. [Google Scholar] [CrossRef]
- Nie, X.; Wei, J.; Hao, Y.; Tao, J.; Li, Y.; Liu, M.; Xu, B.; Li, B. Consistent Biomarkers and Related Pathogenesis Underlying Asthma Revealed by Systems Biology Approach. Int. J. Mol. Sci. 2019, 20, 4037. [Google Scholar] [CrossRef]
- Ananieva, E.A.; Powell, J.D.; Hutson, S.M. Leucine Metabolism in T Cell Activation: mTOR Signaling and Beyond. Adv. Nutr. 2016, 7, 798S–805S. [Google Scholar] [CrossRef]
- Petrache, I.; Berdyshev, E.V. Ceramide Signaling and Metabolism in Pathophysiological States of the Lung. Annu. Rev. Physiol. 2016, 78, 463–480. [Google Scholar] [CrossRef]
- Luo, Y.; Jin, M.; Lou, L.; Yang, S.; Li, C.; Li, X. Role of arachidonic acid lipoxygenase pathway in Asthma. Prostaglandins Other Lipid Mediat. 2022, 158, 106609. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Mild | Severe Uncontrolled | |
|---|---|---|---|
| n | 15 | 11 | 10 |
| Age (years) | 37.4 ± 13.95 * | 36 ± 6.53 * | 50.9 ± 10.62 |
| Onset (years) | - | 14.55 ± 10.21 | 10.7 ± 6.11 |
| Gender (%F/%M) | 60/40 | 63.64/36.36 | 60/40 |
| BMI | 28.71 ± 5.65 | 27.52 ± 5.80 | 28.35 ± 4.00 |
| Smoker (%) | 20 | 9.09 | 0 |
| Ex-smoker (%) | 0 | 0 | 20 |
| Total IgE (U) | - | 551.09 ± 733.8 | 462.1 ± 587.92 |
| AC (%) | 0 | 0 * | 60 |
| AH (%) | 6.67 | 100 * | 30 |
| LTRA (%) | 0 | 18 * | 90 |
| ICS/LABA (%) | 0 | 90.91 | 100 |
| OCS (%) | 0 | 0 | 10 |
| TCS (%) | 6.67 | 100 * | 30 |
| SABA (%) | 0 | 9.09 * | 100 |
| T (%) | 0 | 0 | 10 |
| D. pteronyssinus (%) | 0 | 90.91 | 80 |
| D. farinae (%) | 0 | 90.91 | 80 |
| L. destructor (%) | 0 | 54.55 | 50 |
| B. tropicalis (%) | 0 | 81.82 | 40 |
| A. siro (%) | 0 | 36.36 | 10 |
| T. putrescentiae (%) | 0 | 45.45 | 50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escolar-Peña, A.; Delgado-Dolset, M.I.; Pablo-Torres, C.; Tarin, C.; Mera-Berriatua, L.; Cuesta Apausa, M.d.P.; González Cuervo, H.; Sharma, R.; Kho, A.T.; Tantisira, K.G.; et al. Specific microRNA Profile Associated with Inflammation and Lipid Metabolism for Stratifying Allergic Asthma Severity. Int. J. Mol. Sci. 2024, 25, 9425. https://doi.org/10.3390/ijms25179425
Escolar-Peña A, Delgado-Dolset MI, Pablo-Torres C, Tarin C, Mera-Berriatua L, Cuesta Apausa MdP, González Cuervo H, Sharma R, Kho AT, Tantisira KG, et al. Specific microRNA Profile Associated with Inflammation and Lipid Metabolism for Stratifying Allergic Asthma Severity. International Journal of Molecular Sciences. 2024; 25(17):9425. https://doi.org/10.3390/ijms25179425
Chicago/Turabian StyleEscolar-Peña, Andrea, María Isabel Delgado-Dolset, Carmela Pablo-Torres, Carlos Tarin, Leticia Mera-Berriatua, María del Pilar Cuesta Apausa, Heleia González Cuervo, Rinku Sharma, Alvin T. Kho, Kelan G. Tantisira, and et al. 2024. "Specific microRNA Profile Associated with Inflammation and Lipid Metabolism for Stratifying Allergic Asthma Severity" International Journal of Molecular Sciences 25, no. 17: 9425. https://doi.org/10.3390/ijms25179425
APA StyleEscolar-Peña, A., Delgado-Dolset, M. I., Pablo-Torres, C., Tarin, C., Mera-Berriatua, L., Cuesta Apausa, M. d. P., González Cuervo, H., Sharma, R., Kho, A. T., Tantisira, K. G., McGeachie, M. J., Rebollido-Rios, R., Barber, D., Carrillo, T., Izquierdo, E., & Escribese, M. M. (2024). Specific microRNA Profile Associated with Inflammation and Lipid Metabolism for Stratifying Allergic Asthma Severity. International Journal of Molecular Sciences, 25(17), 9425. https://doi.org/10.3390/ijms25179425