
The Impact of Liver Failure on the Immune System
 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
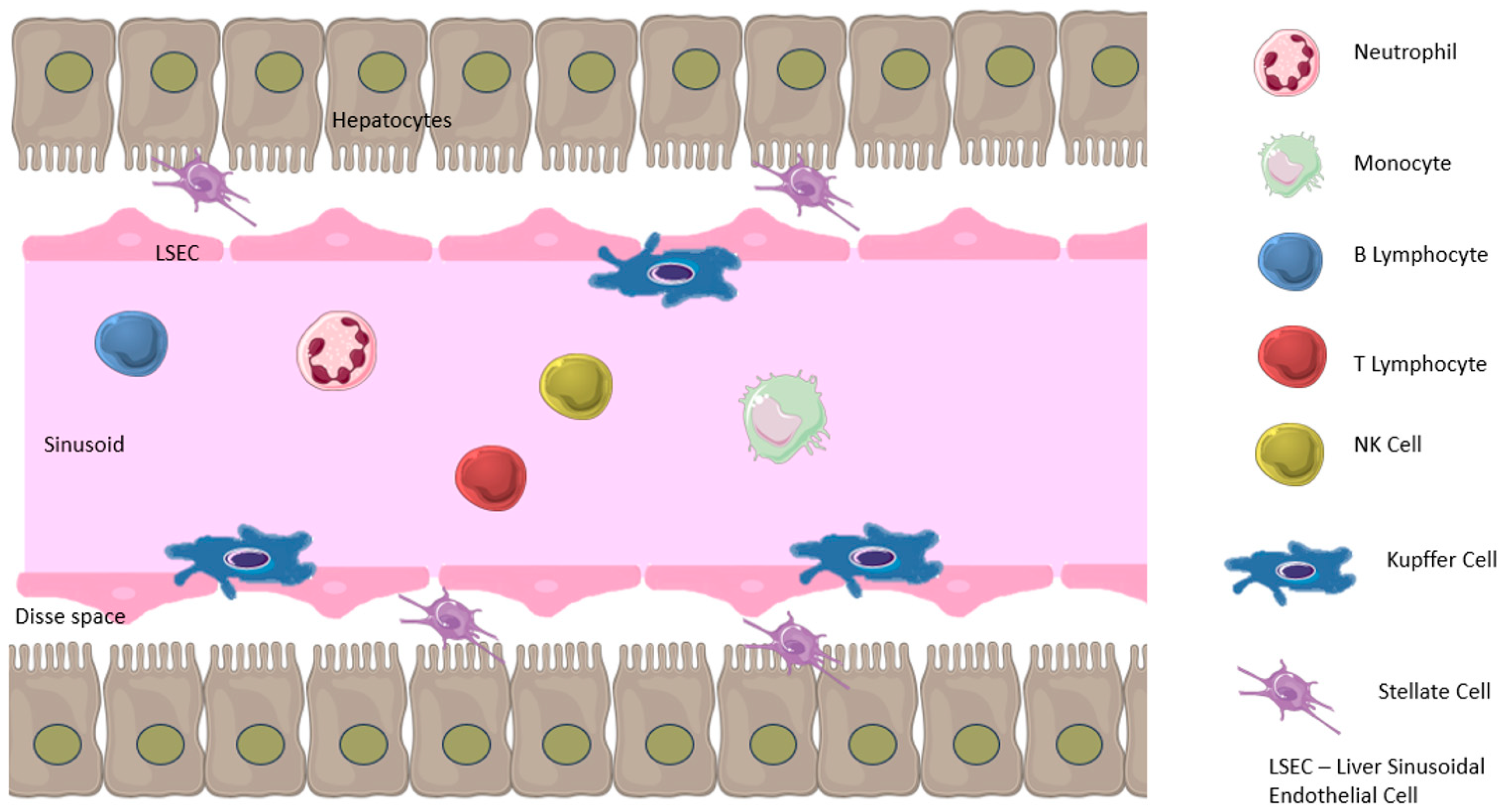
2. Role of the Liver in Immune Response
3. Immunometabolism and Mitochondrial Dysfunction in ACLF
4. The Gut–Liver Axis and Initiation of Hepatic Inflammation
5. Systemic Inflammation
6. Mediators of Inflammation
6.1. Cytokines
6.1.1. Cytokines as Prognostic Biomarkers
6.1.2. Cytokines as Potential Therapeutic Targets
6.2. Chemokines
6.3. Growth Factors
6.4. Lipid Mediators
7. Immunosuppression
7.1. Innate Immune Response
7.1.1. Neutrophils

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neutrophils | |||
|---|---|---|---|
| Author (Year) | Article Type | Number of patients | Conclusions |
| Mookerjee R. P. et al. (2007) [86] | Research Article | 63 patients with alcoholic cirrhosis and cirrhosis superimposed on AH | Full activation of neutrophils resulting from the presence of humoral factors such as endotoxin is associated with reversible impairment of their immune response to infection |
| Fiuza C. et al. (2002) [87] | Research Article | 14 patients with liver cirrhosis and 14 healthy controls | Increased neutrophil adhesion to microvascular endothelium and deficient transendothelial migration in patients with liver cirrhosis. G-CSF increases neutrophil transendothelial migration in patients with cirrhosis |
| Kedarisetty C. K. et al. (2015) [88] | Randomized Controlled Trial | 55 patients with decompensated liver cirrhosis | Reduced mortality and reduced symptom severity during 12 months of follow-up in patients treated with G-CSF and darbepoetin α |
| Konstantis G. et al. (2023) [90] | Systematic Review and Meta-Analysis of Randomized Controlled Trials | 421 patients with ACLF | Reduced mortality and improved MELD scores in patients treated with G-CSF. There is no correlation between improved Child–Pugh score or complication rates. |
| Hou X. et al. (2021) [91] | Systematic Review and Meta-Analysis of Randomized Controlled Trials | 479 patients with ACLF | Overall, there is no association between G-CSF treatment and reduced risk of death or complications. Correlation with better survival in the Asian population. |
7.1.2. Macrophages
7.1.3. Natural Killer Cells (NK Cells)
| Natural Killer Cells (NK Cells) | |||
|---|---|---|---|
| Author (Year) | Article Type | Number of Patients | Conclusions |
| Weiss E. et al. (2021) [99] | Research Article | 67 patients with decompensated cirrhosis (including 35 critically ill patients with ACLF in the intensive care unit) and 12 healthy subjects | Increased numbers of neutrophils and macrophage M0-like monocytes and decreased numbers of several lymphocyte subsets (including memory lymphocytes) in patients with ACLF |
| Radaeva S. et al. (2006) [101] | Research Article | Not applicable (mouse model) | NK cells killed activated hepatic stellate cells (HSCs), alleviating liver fibrosis. This process depended on retinoic acid early inducible 1/NKG2D and tumor necrosis factor-related apoptosis-inducing ligand. |
| Jeong W. Il et al. (2011) [100] | Research Article | Not applicable (mouse model) | Transforming growth factor-β (TGF-β), produced by indirectly activated HSCs, reduced NK cell cytotoxicity, including that targeting HSCs. It has been suggested that retinol/SOCS1/TGF-b metabolites may be therapeutic targets and improve treatment efficacy with IFN-c and NK cell therapy. |
| Li H. J. et al. (2022) [102] | Research Article | 37 HBV-ACLF patients and 13 control subjects | Reduced number of NK cells in the blood of ACLF patients and altered phenotypic and functional profile of NK. The observed association of these disorders with excessive CXCL-10 production |
| Macrophages | |||
|---|---|---|---|
| Author (Year) | Article Type | Number of Patients | Conclusions |
| Bernsmeier C. et al. (2015) [95] | Research article | 41 patients with ACLF, 9 patients with acute decompensation of cirrhosis without ACLF, 17 patients with cirrhosis without decompensation, 23 patients with acute liver failure, 29 healthy individuals | Patients with ACLF have higher numbers of immunoregulatory monocytes and macrophages expressing MERTK, suppressing the innate immune response to microbes. MERTK inhibitors can restore the production of inflammatory cytokines by immune cells from ACLF patients and may be developed to enhance the innate immune response in these individuals |
| Kou K. et al. (2022) [96] | Review article | Not applicable (number of patients not mentioned) | Monocytes and macrophages show decreased HLA-DR expression and increased MERTK expression. |
| Li Z.-H. et al. (2023) [97] | Research article | Not applicable (mouse model) | In ACLF mice, MSCs enhanced liver function and 48-h survival while reducing inflammatory injury by promoting M2 macrophage polarization and increasing Mertk expression levels in macrophages. |
7.2. Adaptive Immune Response
7.2.1. B Cells
7.2.2. T Cells
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACLF | Acute, chronic liver failure |
| ALD | Alcoholic liver disease |
| APCs | Antigen-presenting cells |
| bFGF | Basic fibroblast growth factor |
| CAID | Cirrhosis-associated immune dysfunction |
| CCL5 | C-C motif chemokine ligand 5 |
| CLIF-C | Chronic Liver Failure Consortium |
| COX | Cyclooxygenase |
| DAMPs | Danger-associated molecular patterns |
| EP2 | Prostaglandin E receptor 2 |
| FGF21 | Fibroblast growth factor 21 |
| FoxO | Forkhead box O |
| G-CSF | Granulocyte colony-stimulating factor |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| GPCRs | G-protein-coupled receptors |
| HBV-ACLF | Hepatitis B virus-related acute-on-chronic liver failure |
| HMG1 | High mobility protein group 1 |
| IFN-γ | Interferon-gamma |
| iNKT | Invariant killer T cells |
| KC | Kupffer cells |
| LTE4 | Leukotriene E 4 |
| LSEC | Liver sinusoidal endothelial cells |
| MELD | Model of End-Stage Liver Disease |
| MERTK | Mer receptor tyrosine kinase |
| NAFLD | Non-alcoholic fatty liver disease |
| NK | Natural killer |
| NLRs | NOD-like receptors |
| PAMPs | Pathogen-associated molecular patterns |
| PG | Prostaglandin |
| PGE2 | Derived eicosanoid prostaglandin E2 |
| PHLF | Post-hepatectomy liver failure |
| PRRs | Pattern recognition receptors |
| SCF | Stem cell factor |
| TLRs | Toll-like receptors |
| TIPS | Transjugular intrahepatic portal-systemic shunt |
| TGF-β1/IL-31 | Transforming growth factor β1/interleukin-31 |
| 12-HHT | 12-Hydroxyheptadecatrienoic acid |
References
- Kalaitzakis, E. Gastrointestinal dysfunction in liver cirrhosis. World J. Gastroenterol. 2014, 20, 14686–14695. [Google Scholar] [CrossRef]
- Noor, M.T.; Manoria, P. Immune Dysfunction in Cirrhosis. J. Clin. Transl. Hepatol. 2017, 5, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Hernaez, R.; Kramer, J.R.; Liu, Y.; Tansel, A.; Natarajan, Y.; Hussain, K.B.; Ginès, P.; Solà, E.; Moreau, R.; Gerbes, A.; et al. Prevalence and short-term mortality of acute-on-chronic liver failure: A national cohort study from the USA. J. Hepatol. 2019, 70, 639–647. [Google Scholar] [CrossRef]
- Leão, G.S.; Lunardi, F.L.; Picon, R.V.; Tovo, C.V.; de Mattos, A.A. Acute-on-chronic liver failure: A comparison of three different diagnostic criteria. Ann. Hepatol. 2019, 18, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Sarin, S.K.; Choudhury, A. Acute-on-chronic liver failure: Terminology, mechanisms and management. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.W.; Harmon, C.; O’farrelly, C. Liver immunology and its role in inflammation and homeostasis. Cell. Mol. Immunol. 2016, 13, 267–276. [Google Scholar] [CrossRef]
- Albillos, A.; Martin-Mateos, R.; Van der Merwe, S.; Wiest, R.; Jalan, R.; Álvarez-Mon, M. Cirrhosis-associated immune dysfunction. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 112–134. [Google Scholar] [CrossRef]
- Brandl, K.; Kumar, V.; Eckmann, L. MINI-REVIEW Microbiome and Host Interactions Gut-liver axis at the frontier of host-microbial interactions. Am. J. Physiol. Gastrointest. Liver. Physiol. 2017, 312, G413–G419. [Google Scholar] [CrossRef] [PubMed]
- Bilzer, M.; Roggel, F.; Gerbes, A.L. Role of Kupffer cells in host defense and liver disease. Liver Int. 2006, 26, 1175–1186. [Google Scholar] [CrossRef]
- Sana, G.; Lombard, C.; Vosters, O.; Jazouli, N.; Andre, F.; Stephenne, X.; Smets, F.; Najimi, M.; Sokal, E.M. Adult Human Hepatocytes Promote CD4+ T-Cell Hyporesponsiveness Via Interleukin-10-Producing Allogeneic Dendritic Cells. Cell Transplant. 2014, 23, 1127–1142. [Google Scholar] [CrossRef]
- Wen, Y.; Lambrecht, J.; Ju, C.; Tacke, F. Hepatic macrophages in liver homeostasis and diseases-diversity, plasticity and therapeutic opportunities. Cell Mol. Immunol. 2021, 18, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Furze, R.C.; Rankin, S.M. Neutrophil mobilization and clearance in the bone marrow. Immunology 2008, 125, 281–288. [Google Scholar] [CrossRef]
- Gu, X.; Chu, Q.; Ma, X.; Wang, J.; Chen, C.; Guan, J.; Ren, Y.; Wu, S.; Zhu, H. New insights into iNKT cells and their roles in liver diseases. Front. Immunol. 2022, 13, 1035950. [Google Scholar] [CrossRef] [PubMed]
- Zhang, I.W.; López-Vicario, C.; Duran-Güell, M.; Clària, J. Mitochondrial Dysfunction in Advanced Liver Disease: Emerging Concepts. Front. Mol. Biosci. 2021, 8, 772174. [Google Scholar] [CrossRef]
- Zaccherini, G.; Weiss, E.; Moreau, R. Acute-on-chronic liver failure: Definitions, pathophysiology and principles of treatment. JHEP Rep. 2021, 3, 100176. [Google Scholar] [CrossRef]
- Moreau, R.; Gao, B.; Papp, M.; Bañares, R.; Kamath, P.S. Acute-on-chronic liver failure: A distinct clinical syndrome. J. Hepatol. 2021, 75, S27–S35. [Google Scholar] [CrossRef]
- Bhatti, G.K.; Gupta, A.; Pahwa, P.; Khullar, N.; Singh, S.; Navik, U.; Kumar, S.; Mastana, S.S.; Reddy, A.P.; Reddy, P.H.; et al. Targeting mitochondrial bioenergetics as a promising therapeutic strategy in metabolic and neurodegenerative diseases. Biomed. J. 2022, 45, 733–748. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Peiseler, M.; Schwabe, R.; Hampe, J.; Kubes, P.; Heikenwälder, M.; Tacke, F. Immune mechanisms linking metabolic injury to inflammation and fibrosis in fatty liver disease—Novel insights into cellular communication circuits. J. Hepatol. 2022, 77, 1136–1160. [Google Scholar] [CrossRef]
- Zhang, I.W.; Curto, A.; López-Vicario, C.; Casulleras, M.; Duran-Güell, M.; Flores-Costa, R.; Colsch, B.; Aguilar, F.; Aransay, A.M.; Lozano, J.J.; et al. Mitochondrial dysfunction governs immunometabolism in leukocytes of patients with acute-on-chronic liver failure. J. Hepatol. 2022, 76, 93–106. [Google Scholar] [CrossRef]
- Yang, X.; Lu, D.; Zhuo, J.; Lin, Z.; Yang, M.; Xu, X. The Gut-liver Axis in Immune Remodeling: New insight into Liver Diseases. Int. J. Biol. Sci. 2020, 16, 2357–2366. [Google Scholar] [CrossRef]
- Guan, H.; Zhang, X.; Kuang, M.; Yu, J. The gut–liver axis in immune remodeling of hepatic cirrhosis. Front. Immunol. 2022, 13, 946628. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.C.; Harsch, I.A.; Konturek, K.; Schink, M.; Konturek, T.; Neurath, M.F.; Zopf, Y. Gut–Liver Axis: How Do Gut Bacteria Influence the Liver? Med. Sci. 2018, 6, 79. [Google Scholar] [CrossRef] [PubMed]
- Martin-Mateos, R.; Alvarez-Mon, M.; Albillos, A. Dysfunctional Immune Response in Acute-on-Chronic Liver Failure: It Takes Two to Tango. Front. Immunol. 2019, 10, 973. [Google Scholar] [CrossRef]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 polarization. Eur. J. Pharmacol. 2020, 877, 173090. [Google Scholar] [CrossRef] [PubMed]
- Clària, J.; Arroyo, V.; Moreau, R. Roles of systemic inflammatory and metabolic responses in the pathophysiology of acute-on-chronic liver failure. JHEP Rep. 2023, 5, 100807. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hu, J.; Gou, C.; Jin, H.; Zhang, C.; Liu, Y.; Wang, Y.; Wang, X. Serum Interleukins as Potential Prognostic Biomarkers in HBV-Related Acute-on-Chronic Liver Failure. Mediat. Inflamm. 2022, 2022, 7794890. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Dinarello, C.A.; Molgora, M.; Garlanda, C. Interleukin-1 and Related Cytokines in the Regulation of Inflammation and Immunity. Immunity 2019, 50, 778–795. [Google Scholar] [CrossRef]
- Clària, J.; Arroyo, V.; Moreau, R. The Acute-on-Chronic Liver Failure Syndrome, or When the Innate Immune System Goes Astray. J. Immunol. 2016, 197, 3755–3761. [Google Scholar] [CrossRef]
- Hirano, T. IL-6 in inflammation, autoimmunity and cancer. Int. Immunol. 2019, 33, 127–148. [Google Scholar] [CrossRef]
- Karatayli, E.; Hall, R.A.; Weber, S.N.; Dooley, S.; Lammert, F. Effect of alcohol on the interleukin 6-mediated inflammatory response in a new mouse model of acute-on-chronic liver injury. Biochim. Et Biophys. Acta Mol. Basis Dis. 2019, 1865, 298–307. [Google Scholar] [CrossRef]
- Clària, J.; Stauber, R.E.; Coenraad, M.J.; Moreau, R.; Jalan, R.; Pavesi, M.; Amorós; Titos, E.; Alcaraz-Quiles, J.; Oettl, K.; et al. Systemic inflammation in decompensated cirrhosis: Characterization and role in acute-on-chronic liver failure. Hepatology 2016, 64, 1249–1264. [Google Scholar] [CrossRef]
- Dirchwolf, M.; Podhorzer, A.; Marino, M.; Shulman, C.; Cartier, M.; Zunino, M.; Paz, S.; Muñoz, A.; Bocassi, A.; Gimenez, J.; et al. Immune dysfunction in cirrhosis: Distinct cytokines phenotypes according to cirrhosis severity. Cytokine 2016, 77, 14–25. [Google Scholar] [CrossRef]
- Zanza, C.; Romenskaya, T.; Manetti, A.C.; Franceschi, F.; La Russa, R.; Bertozzi, G.; Maiese, A.; Savioli, G.; Volonnino, G.; Longhitano, Y. Cytokine Storm in COVID-19: Immunopathogenesis and Therapy. Medicina 2022, 58, 144. [Google Scholar] [CrossRef]
- Xiang, X.; Hwang, S.; Feng, D.; Shah, V.H.; Gao, B. Interleukin-22 in alcoholic hepatitis and beyond. Hepatol. Int. 2020, 14, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Cron, R.Q.; Goyal, G.; Chatham, W.W. Cytokine Storm Syndrome. Annu. Rev. Med. 2023, 74, 321–337. [Google Scholar] [CrossRef]
- Remmler, J.; Schneider, C.; Treuner-Kaueroff, T.; Bartels, M.; Seehofer, D.; Scholz, M.; Berg, T.; Kaiser, T. Increased Level of Interleukin 6 Associates with Increased 90-Day and 1-Year Mortality in Patients with End-Stage Liver Disease. Clin. Gastroenterol. Hepatol. 2018, 16, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Xiao, N.; Liu, L.; Zhang, Y.; Nie, Y.; Zhu, X. A practical nomogram based on serum interleukin-6 for the prognosis of liver failure. Front. Med. 2022, 9, 1035699. [Google Scholar] [CrossRef] [PubMed]
- Labenz, C.; Toenges, G.; Huber, Y.; Nagel, M.; Marquardt, J.U.; Schattenberg, J.M.; Galle, P.R.; Labenz, J.; Wörns, M. Raised serum Interleukin-6 identifies patients with liver cirrhosis at high risk for overt hepatic encephalopathy. Aliment. Pharmacol. Ther. 2019, 50, 1112–1119. [Google Scholar] [CrossRef]
- Tornai, D.; Mitchell, M.; McClain, C.J.; Dasarathy, S.; McCullough, A.; Radaeva, S.; Kroll-Desrosiers, A.; Lee, J.; Barton, B.; Szabo, G. A novel score of IL-13 and age predicts 90-day mortality in severe alcohol-associated hepatitis: A multicenter plasma biomarker analysis. Hepatol. Commun. 2023, 7, e0296. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, H.; Pan, L.; Zhang, M.; Wan, X.; Xu, H.; Hua, R.; Zhu, M.; Gao, P. Systemic inflammatory regulators and risk of acute-on-chronic liver failure: A bidirectional mendelian-randomization study. Front. Cell Dev. Biol. 2023, 11, 1125233. [Google Scholar] [CrossRef]
- Choy, E.H.; De Benedetti, F.; Takeuchi, T.; Hashizume, M.; John, M.R.; Kishimoto, T. Translating IL-6 biology into effective treatments. Nat. Rev. Rheumatol. 2020, 16, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Feng, D.; Hwang, S.; Ren, T.; Wang, X.; Trojnar, E.; Matyas, C.; Mo, R.; Shang, D.; He, Y.; et al. Interleukin-22 ameliorates acute-on-chronic liver failure by reprogramming impaired regeneration pathways in mice. J. Hepatol. 2020, 72, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.; Hicks, A.; Hoo, C.Z.; Kwon, Y.S.; Cho, Y.E.; Moore, J.; Gao, B. Novel treatment of acute and acute-on-chronic liver failure: Interleukin-22. Liver Int. 2023. [Google Scholar] [CrossRef] [PubMed]
- Arab, J.P.; Sehrawat, T.S.; Simonetto, D.A.; Verma, V.K.; Feng, D.; Tang, T.; Dreyer, K.; Yan, X.; Daley, W.L.; Sanyal, A.; et al. An Open-Label, Dose-Escalation Study to Assess the Safety and Efficacy of IL-22 Agonist F-652 in Patients with Alcohol-associated Hepatitis. Hepatology 2020, 72, 441–453. [Google Scholar] [CrossRef]
- Cook, S.A. The Pathobiology of Interleukin 11 in Mammalian Disease is Likely Explained by its Essential Evolutionary Role for Fin Regeneration. J. Cardiovasc. Transl. Res. 2023, 16, 755–757. [Google Scholar] [CrossRef]
- Ng, B.; Xie, C.; Su, L.; Kuthubudeen, F.F.; Kwek, X.-Y.; Yeong, D.; Pua, C.J.; Cook, S.A.; Lim, W.-W. IL11 (Interleukin-11) Causes Emphysematous Lung Disease in a Mouse Model of Marfan Syndrome. Arter. Thromb. Vasc. Biol. 2023, 43, 739–754. [Google Scholar] [CrossRef] [PubMed]
- Widjaja, A.A.; Singh, B.K.; Adami, E.; Viswanathan, S.; Dong, J.; D’agostino, G.A.; Ng, B.; Lim, W.W.; Tan, J.; Paleja, B.S.; et al. Inhibiting Interleukin 11 Signaling Reduces Hepatocyte Death and Liver Fibrosis, Inflammation, and Steatosis in Mouse Models of Nonalcoholic Steatohepatitis. Gastroenterology 2019, 157, 777–792.e14. [Google Scholar] [CrossRef]
- Widjaja, A.A.; Viswanathan, S.; Shekeran, S.G.; Adami, E.; Lim, W.-W.; Chothani, S.; Tan, J.; Goh, J.W.T.; Chen, H.M.; Lim, S.Y.; et al. Targeting endogenous kidney regeneration using anti-IL11 therapy in acute and chronic models of kidney disease. Nat. Commun. 2022, 13, 7497. [Google Scholar] [CrossRef]
- Gustavsson, M. New insights into the structure and function of chemokine receptor: Chemokine complexes from an experimental perspective. J. Leukoc. Biol. 2020, 107, 1115–1122. [Google Scholar] [CrossRef]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef]
- Lei, W.; Jia, L.; Wang, Z.; Liang, Z.; Zhao, A.; Liu, Y.; Tian, Y.; Zhao, L.; Chen, Y.; Shi, G.; et al. CC chemokines family in fibrosis and aging: From mechanisms to therapy. Ageing Res. Rev. 2023, 87, 101900. [Google Scholar] [CrossRef]
- Khanam, A.; Trehanpati, N.; Riese, P.; Rastogi, A.; Guzman, C.A.; Sarin, S.K. Blockade of Neutrophil’s Chemokine Receptors CXCR1/2 Abrogate Liver Damage in Acute-on-Chronic Liver Failure. Front. Immunol. 2017, 8, 464. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, K.; Yang, D.; Oppenheim, J.J. Interleukin-8: An evolving chemokine. Cytokine 2022, 153, 155828. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-H.; Wang, M.-L.; Tao, Y.-C.; Wu, D.-B.; Chen, E.-Q.; Tang, H. The high level of IL-1β in the serum of ACLF patients induces increased IL-8 expression in hUC-MSCs and reduces the efficacy of hUC-MSCs in liver failure. Stem Cell Res. Ther. 2023, 14, 231. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Suzuki, Y.; Kakisaka, K.; Wang, T.; Ishida, K.; Suzuki, A.; Abe, H.; Sugai, T.; Takikawa, Y. IL-8 induces transdifferentiation of mature hepatocytes toward the cholangiocyte phenotype. FEBS Open Bio 2019, 9, 2105–2116. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, P.; Xiao, N.; Liu, Q.; Zhu, X. Interleukin-8 predicts short-term mortality in acute-on-chronic liver failure patients with hepatitis B-related-related cirrhosis background. Ann. Med. 2023, 55, 2287708. [Google Scholar] [CrossRef]
- Zhu, B.; Gao, F.; Li, Y.; Shi, K.; Hou, Y.; Chen, J.; Zhang, Q.; Wang, X. Serum cytokine and chemokine profiles and disease prognosis in hepatitis B virus-related acute-on-chronic liver failure. Front. Immunol. 2023, 14, 1133656. [Google Scholar] [CrossRef]
- Xiao, L.; Tang, S.; Zhang, L.; Ma, S.; Zhao, Y.; Zhang, F.; Xie, Z.; Li, L. Serum CXCL1 Is a Prognostic Factor for Patients with Hepatitis B Virus–Related Acute-On-Chronic Liver Failure. Front. Med. 2021, 8, 657076. [Google Scholar] [CrossRef]
- Tang, S.; Zhang, J.; Zhang, L.; Zhao, Y.; Xiao, L.; Zhang, F.; Li, Q.; Yang, Y.; Liu, Q.; Xu, J.; et al. Knockdown of CXCL1 improves ACLF by reducing neutrophil recruitment to attenuate ROS production and hepatocyte apoptosis. Hepatol. Commun. 2023, 7, e0257. [Google Scholar] [CrossRef]
- Liu, G.; Wang, X.; Yang, T.; Yan, Y.; Xiang, T.; Yang, L.; Luo, X. High Interleukin-8 Levels Associated with Decreased Survival in Patients with Cirrhosis Following Transjugular Intrahepatic Portosystemic Shunt. Front. Med. 2022, 9, 829245. [Google Scholar] [CrossRef]
- Berres, M.-L.; Asmacher, S.; Lehmann, J.; Jansen, C.; Görtzen, J.; Klein, S.; Meyer, C.; Strunk, H.M.; Fimmers, R.; Tacke, F.; et al. CXCL9 is a prognostic marker in patients with liver cirrhosis receiving transjugular intrahepatic portosystemic shunt. J. Hepatol. 2015, 62, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.M.; Claus, K.; Jansen, C.; Pohlmann, A.; Schierwagen, R.; Meyer, C.; Thomas, D.; Manekeller, S.; Claria, J.; Strassburg, C.P.; et al. Circulating CXCL10 in cirrhotic portal hypertension might reflect systemic inflammation and predict ACLF and mortality. Liver Int. 2018, 38, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Jiao, J.; Cai, H.; Zhang, Y.; Xia, Y.; Lin, J.; Shang, Z.; Qian, Y.; Wang, F.; Wu, H.; et al. C-C motif chemokine ligand 5 confines liver regeneration by down-regulating reparative macrophage-derived hepatocyte growth factor in a forkhead box O 3a–dependent manner. Hepatology 2022, 76, 1706–1722. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, H.M.; Pitts, K.; Wang, T.; Lee, E.; Buchbinder, E.; Dougan, M.; Armstrong, D.G.; Paine, R.; Ragsdale, C.E.; Boyd, T.; et al. Recombinant GM-CSF for diseases of GM-CSF insufficiency: Correcting dysfunctional mononuclear phagocyte disorders. Front. Immunol. 2023, 13, 1069444. [Google Scholar] [CrossRef]
- Lu, W.-Y.; Bird, T.G.; Boulter, L.; Tsuchiya, A.; Cole, A.M.; Hay, T.; Guest, R.V.; Wojtacha, D.; Man, T.Y.; Mackinnon, A.; et al. Hepatic progenitor cells of biliary origin with liver repopulation capacity. Nat. Cell Biol. 2015, 17, 971–983. [Google Scholar] [CrossRef]
- Simonetto, D.A.; Shah, V.H.; Kamath, P.S. Improving survival in ACLF: Growing evidence for use of G-CSF. Hepatol. Int. 2017, 11, 473–475. [Google Scholar] [CrossRef]
- Engelmann, C.; Habtesion, A.; Hassan, M.; Kerbert, A.J.; Hammerich, L.; Novelli, S.; Fidaleo, M.; Philips, A.; Davies, N.; Ferreira-Gonzalez, S.; et al. Combination of G-CSF and a TLR4 inhibitor reduce inflammation and promote regeneration in a mouse model of ACLF. J. Hepatol. 2022, 77, 1325–1338. [Google Scholar] [CrossRef]
- Yu, X.; Guo, R.; Ming, D.; Deng, Y.; Su, M.; Lin, C.; Li, J.; Lin, Z.; Su, Z. The Transforming Growth Factor β1/Interleukin-31 Pathway Is Upregulated in Patients with Hepatitis B Virus-Related Acute-on-Chronic Liver Failure and Is Associated with Disease Severity and Survival. Clin. Vaccine Immunol. 2015, 22, 484–492. [Google Scholar] [CrossRef]
- Ruiz-Margáin, A.; Pohlmann, A.; Ryan, P.; Schierwagen, R.; Chi-Cervera, L.A.; Jansen, C.; Mendez-Guerrero, O.; Flores-García, N.C.; Lehmann, J.; Torre, A.; et al. Fibroblast growth factor 21 is an early predictor of acute-on-chronic liver failure in critically ill patients with cirrhosis. Liver Transplant. 2018, 24, 595–605. [Google Scholar] [CrossRef]
- Narumiya, S.; Yokomizo, T.; Aoki, J. Lipid Mediators in Inflammation. In Inflammation—From Molecular and Cellular Mechanisms to the Clinic; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2017. [Google Scholar] [CrossRef]
- Artru, F.; McPhail, M.J.W.; Triantafyllou, E.; Trovato, F.M. Lipids in Liver Failure Syndromes: A Focus on Eicosanoids, Specialized Pro-Resolving Lipid Mediators and Lysophospholipids. Front. Immunol. 2022, 13, 867261. [Google Scholar] [CrossRef]
- López-Vicario, C.; Checa, A.; Urdangarin, A.; Aguilar, F.; Alcaraz-Quiles, J.; Caraceni, P.; Amorós, A.; Pavesi, M.; Gómez-Cabrero, D.; Trebicka, J.; et al. Targeted lipidomics reveals extensive changes in circulating lipid mediators in patients with acutely decompensated cirrhosis. J. Hepatol. 2020, 73, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, F.; Yokomizo, T. The leukotriene receptors as therapeutic targets of inflammatory diseases. Int. Immunol. 2019, 31, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Casulleras, M.; Flores-Costa, R.; Duran-Güell, M.; Zhang, I.W.; López-Vicario, C.; Curto, A.; Fernández, J.; Arroyo, V.; Clària, J. Albumin Lipidomics Reveals Meaningful Compositional Changes in Advanced Cirrhosis and Its Potential to Promote Inflammation Resolution. Hepatol. Commun. 2022, 6, 1443–1456. [Google Scholar] [CrossRef] [PubMed]
- Becares, N.; Härmälä, S.; China, L.; Colas, R.A.; Maini, A.A.; Bennet, K.; Skene, S.S.; Shabir, Z.; Dalli, J.; O’brien, A. Immune Regulatory Mediators in Plasma from Patients With Acute Decompensation Are Associated with 3-Month Mortality. Clin. Gastroenterol. Hepatol. 2020, 18, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, A.J.; Fullerton, J.N.; A Massey, K.; Auld, G.; Sewell, G.; James, S.; Newson, J.; Karra, E.; Winstanley, A.; Alazawi, W.; et al. Immunosuppression in acutely decompensated cirrhosis is mediated by prostaglandin E2. Nat. Med. 2014, 20, 518–523. [Google Scholar] [CrossRef]
- Clària, J.; Curto, A.; Moreau, R.; Colsch, B.; López-Vicario, C.; Lozano, J.J.; Aguilar, F.; Castelli, F.A.; Fenaille, F.; Junot, C.; et al. Untargeted lipidomics uncovers lipid signatures that distinguish severe from moderate forms of acutely decompensated cirrhosis. J. Hepatol. 2021, 75, 1116–1127. [Google Scholar] [CrossRef]
- Huang, X.-P.; Wang, Y.; Chen, L.; Sun, W.; Huang, Y.; Xu, Y.; Feng, T.-T.; Luo, E.-P.; Qin, A.-L.; Zhao, W.-F.; et al. Elevated serum prostaglandin E2 predicts the risk of infection in hepatitis B virus-related acute-on-chronic liver failure patients. Asian Pac. J. Trop. Med. 2017, 10, 916–920. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, C.; Qi, J.; Wu, F.; Guan, J.; Chen, Z.; Zhu, H. Altered PGE2-EP2 is associated with an excessive immune response in HBV-related acute-on-chronic liver failure. J. Transl. Med. 2019, 17, 93. [Google Scholar] [CrossRef]
- China, L.; Maini, A.; Skene, S.S.; Shabir, Z.; Sylvestre, Y.; Colas, R.A.; Ly, L.; Salles, N.B.; Belloti, V.; Dalli, J.; et al. Albumin Counteracts Immune-Suppressive Effects of Lipid Mediators in Patients with Advanced Liver Disease. Clin. Gastroenterol. Hepatol. 2018, 16, 738–747.e7. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Ding, H.; Shi, X.; Ren, H. Mesenchymal stem cell-secreted prostaglandin E2 ameliorates acute liver failure via attenuation of cell death and regulation of macrophage polarization. Stem Cell Res. Ther. 2021, 12, 338. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.; Dronfield, M.; Toghill, P.; Reeves, W. Neutrophil function in chronic liver disease. Clin. Exp. Immunol. 1981, 45, 81–89. [Google Scholar]
- Tritto, G.; Bechlis, Z.; Stadlbauer, V.; Davies, N.; Francés, R.; Shah, N.; Mookerjee, R.P.; Such, J.; Jalan, R. Evidence of neutrophil functional defect despite inflammation in stable cirrhosis. J. Hepatol. 2011, 55, 574–581. [Google Scholar] [CrossRef] [PubMed]
- DeMeo, A.N.; Andersen, B.R.; English, D.K.; Peterson, J. Defective Chemotaxis Associated with a Serum Inhibitor in Cirrhotic Patients. J. Lab. Clin. Med. 1971, 286, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Van Epps, D.E.; Strickland, R.G.; Williams, R.C. Inhibitors of leukocyte chemotaxis in alcoholic liver disease. Am. J. Med. 1975, 59, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Mookerjee, R.P.; Stadlbauer, V.; Lidder, S.; Wright, G.A.; Hodges, S.J.; Davies, N.A.; Jalan, R. Neutrophil dysfunction in alcoholic hepatitis superimposed on cirrhosis is reversible and predicts the outcome. Hepatology 2007, 46, 831–840. [Google Scholar] [CrossRef]
- Fiuza, C.; Salcedo, M.; Clemente, G.; Tellado, J.M. Granulocyte Colony-Stimulating Factor Improves Deficient In Vitro Neutrophil Transendothelial Migration in Patients with Advanced Liver Disease. Clin. Vaccine Immunol. 2002, 9, 433–439. [Google Scholar] [CrossRef]
- Kedarisetty, C.K.; Anand, L.; Bhardwaj, A.; Bhadoria, A.S.; Kumar, G.; Vyas, A.K.; David, P.; Trehanpati, N.; Rastogi, A.; Bihari, C.; et al. Combination of Granulocyte Colony-Stimulating Factor and Erythropoietin Improves Outcomes of Patients with Decompensated Cirrhosis. Gastroenterology 2015, 148, 1362–1370.e7. [Google Scholar] [CrossRef]
- Rolando, N.; Wade, J.; Davalos, M.; Wendon, J.; Philpott-Howard, J.; Williams, R. The Systemic Inflammatory Response Syndrome in Acute Liver Failure. Hepatology 2000, 32, 734–739. [Google Scholar] [CrossRef]
- Konstantis, G.; Tsaousi, G.; Pourzitaki, C.; Kitsikidou, E.; Magouliotis, D.E.; Wiener, S.; Zeller, A.C.; Willuweit, K.; Schmidt, H.H.; Rashidi-Alavijeh, J. Efficacy of Granulocyte Colony-Stimulating Factor in Acute on Chronic Liver Failure: A Systematic Review and Survival Meta-Analysis. J. Clin. Med. 2023, 12, 6541. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Li, Y.; Yuan, H.; Cai, J.; Liu, R.; Li, J.; Zhu, C. Therapeutic Effect and Safety of Granulocyte Colony-Stimulating Factor Therapy for Acute-On-Chronic Liver Failure: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Front. Med. 2021, 8, 784240. [Google Scholar] [CrossRef]
- Choe, W.H.; Baik, S.K. Prostaglandin E2-mediated immunosuppression and the role of albumin as its modulator. Hepatology 2015, 61, 1080–1082. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, V.; Moreau, R. Tying up PGE2 with albumin to relieve immunosuppression in cirrhosis. Nat. Med. 2014, 20, 467–469. [Google Scholar] [CrossRef]
- Bernsmeier, C.; Pop, O.T.; Singanayagam, A.; Triantafyllou, E.; Patel, V.C.; Weston, C.J.; Curbishley, S.; Sadiq, F.; Vergis, N.; Khamri, W.; et al. Patients With Acute-on-Chronic Liver Failure Have Increased Numbers of Regulatory Immune Cells Expressing the Receptor Tyrosine Kinase MERTK. Gastroenterology 2015, 148, 603–615.e14. [Google Scholar] [CrossRef]
- Kou, K.; Sun, X.; Tian, G.; Zhi, Y.; Fan, Z.; Lv, G. The Mechanisms of Systemic Inflammatory and Immunosuppressive Acute-on-Chronic Liver Failure and Application Prospect of Single-Cell Sequencing. J. Immunol. Res. 2022, 2022, 5091275. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-H.; Chen, J.-F.; Zhang, J.; Lei, Z.-Y.; Wu, L.-L.; Meng, S.-B.; Wang, J.-L.; Xiong, J.; Lin, D.-N.; Wang, J.-Y.; et al. Mesenchymal Stem Cells Promote Polarization of M2 Macrophages in Mice with Acute-On-Chronic Liver Failure via Mertk/JAK1/STAT6 Signaling. Stem Cells 2023, 41, 1171–1184. [Google Scholar] [CrossRef]
- Lian, Z.X.; Li, L. The Liver as a Lymphoid Organ. In Liver Immunology: Principles and Practice; Springer: Berlin/Heidelberg, Germany, 2020; pp. 17–23. [Google Scholar] [CrossRef]
- Weiss, E.; de la Grange, P.; Defaye, M.; Lozano, J.J.; Aguilar, F.; Hegde, P.; Jolly, A.; Moga, L.; Sukriti, S.; Agarwal, B.; et al. Characterization of Blood Immune Cells in Patients With Decompensated Cirrhosis Including ACLF. Front. Immunol. 2021, 11, 619039. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.-I.; Park, O.; Suh, Y.-G.; Byun, J.-S.; Park, S.-Y.; Choi, E.; Kim, J.-K.; Ko, H.; Wang, H.; Miller, A.M.; et al. Suppression of innate immunity (natural killer cell/interferon-γ) in the advanced stages of liver fibrosis in mice. Hepatology 2011, 53, 1342–1351. [Google Scholar] [CrossRef] [PubMed]
- Radaeva, S.; Sun, R.; Jaruga, B.; Nguyen, V.T.; Tian, Z.; Gao, B. Natural Killer Cells Ameliorate Liver Fibrosis by Killing Activated Stellate Cells in NKG2D-Dependent and Tumor Necrosis Factor–Related Apoptosis-Inducing Ligand–Dependent Manners. Gastroenterology 2006, 130, 435–452. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-J.; Yang, N.; Mu, X.; Tang, L.; Wang, S.-S.; Zhou, C.-B.; Yuan, J.-H.; Wang, H.-Y.; Yu, Y.-Y.; Li, J.; et al. Reduction of natural killer cells is associated with poor outcomes in patients with hepatitis B virus-related acute-on-chronic liver failure. Hepatol. Int. 2022, 16, 1398–1411. [Google Scholar] [CrossRef]
- Br, V.K.; Sarin, S.K. Acute-on-chronic liver failure: Terminology, mechanisms and management. Clin. Mol. Hepatol. 2023, 29, 670–689. [Google Scholar] [CrossRef]
- Wang, N.; He, S.; Zheng, Y.; Wang, L. The value of NLR versus MLR in the short-term prognostic assessment of HBV-related acute-on-chronic liver failure. Int. Immunopharmacol. 2023, 121, 110489. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, X.; Saredy, J.; Yuan, Z.; Yang, X.; Wang, H. Innate-adaptive immunity interplay and redox regulation in immune response. Redox Biol. 2020, 37, 101759. [Google Scholar] [CrossRef] [PubMed]
- Hensley, M.K.; Deng, J.C. Acute on Chronic Liver Failure and Immune Dysfunction: A Mimic of Sepsis. Semin. Respir. Crit. Care Med. 2018, 39, 588–597. [Google Scholar] [CrossRef]
- Mehta, A.S.; Long, R.E.; Comunale, M.A.; Wang, M.; Rodemich, L.; Krakover, J.; Philip, R.; Marrero, J.A.; Dwek, R.A.; Block, T.M. Increased Levels of Galactose-Deficient Anti-Gal Immunoglobulin G in the Sera of Hepatitis C Virus-Infected Individuals with Fibrosis and Cirrhosis. J. Virol. 2008, 82, 1259–1270. [Google Scholar] [CrossRef] [PubMed]
- Tangye, S.G.; Good, K.L. Human IgM+CD27+ B Cells: Memory B Cells or “Memory” B Cells? J. Immunol. 2007, 179, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Cook, R.T.; Waldschmidt, T.J.; Cook, B.L.; Labrecque, D.R.; Mclatchie, K. Loss of the CD5+ and CD45RAhi B cell subsets in alcoholics. Clin. Exp. Immunol. 1996, 103, 304–310. [Google Scholar] [CrossRef]
- Hassnine, A.A.; Saber, M.A.; Fouad, Y.M.; Sarhan, H.; Elsayed, M.M.; Zaki, Z.M.; Abdelraheem, E.M.; Abdelhalim, S.M.; Elsayed, A.M. Clinical study on the efficacy of hepatitis B vaccination in hepatitis C virus related chronic liver diseases in Egypt. Virus Res. 2023, 323, 198953. [Google Scholar] [CrossRef] [PubMed]
- Ashhab, A.A.; Rodin, H.; Campos, M.; Abu-Sulb, A.; Hall, J.A.; Powell, J.; Debes, J.D. Response to hepatitis B virus vaccination in individuals with chronic hepatitis C virus infection. PLoS ONE 2020, 15, e0237398. [Google Scholar] [CrossRef]
- Morishima, C.; Di Bisceglie, A.M.; Rothman, A.L.; Bonkovsky, H.L.; Lindsay, K.L.; Lee, W.M.; Koziel, M.J.; Fontana, R.J.; Kim, H.; Wright, E.C.; et al. Antigen-specific T lymphocyte proliferation decreases over time in advanced chronic hepatitis C. J. Viral Hepat. 2012, 19, 404–413. [Google Scholar] [CrossRef]
- Bernsmeier, C.; Triantafyllou, E.; Brenig, R.; Lebosse, F.J.; Singanayagam, A.; Patel, V.C.; Pop, O.T.; Khamri, W.; Nathwani, R.; Tidswell, R.; et al. CD14+ CD15− HLA-DR− myeloid-derived suppressor cells impair antimicrobial responses in patients with acute-on-chronic liver failure. Gut 2017, 67, 1155–1167. [Google Scholar] [CrossRef]
- Shen, C.; Yan, W.-Z.; Zhao, C.-Y.; Che, H.-H.; Liu, X.-Y.; Liu, Z.-Z.; Wang, Y.-D.; Wang, W.; Li, M.; Gao, J. Increased CD4+CD25+ regulatory T cells correlate with poor short-term outcomes in hepatitis B virus-related acute-on-chronic liver failure patients. J. Microbiol. Immunol. Infect. 2015, 48, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Shi, H.; Zhang, L.; Shi, H.; Miao, X.; Chen, L.; Chen, Y.; Ma, Y. Comparative analysis of monocyte-derived dendritic cell phenotype and T cell stimulatory function in patients with acute-on-chronic liver failure with different clinical parameters. Front. Immunol. 2023, 14, 1290445. [Google Scholar] [CrossRef] [PubMed]
- Adenote, A.; Dumic, I.; Madrid, C.; Barusya, C.; Nordstrom, C.W.; Prada, L.R. NAFLD and Infection, a Nuanced Relationship. Can. J. Gastroenterol. Hepatol. 2021, 2021, 5556354. [Google Scholar] [CrossRef]
- Li, B.; Zhang, C.; Zhan, Y.-T. Nonalcoholic Fatty Liver Disease Cirrhosis: A Review of Its Epidemiology, Risk Factors, Clinical Presentation, Diagnosis, Management, and Prognosis. Can. J. Gastroenterol. Hepatol. 2018, 2018, 2784537. [Google Scholar] [CrossRef]
- Widjaja, A.A.; Chothani, S.P.; Cook, S.A. Different roles of interleukin 6 and interleukin 11 in the liver: Implications for therapy. Hum. Vaccines Immunother. 2020, 16, 2357–2362. [Google Scholar] [CrossRef] [PubMed]



| Impaired Function of Immune System | Treatment Possibilities | Treatment Effects |
|---|---|---|
| Activation of IFN-Î3/STAT1 anti-regenerative pathway | F-652 (IL-22 Fc) | Reduction in Lille and MELD scores, decrease in serum cytokine and chemokine levels. |
| Increased levels of IL-11 | Anti IL-11 antibodies | Reduced fibrosis, anti-inflammatory effect, reduced hepatocyte death |
| Increased levels of PGE2 | Human albumin solution | Improved macrophage function, Reduced infection risk |
| Impaired neutrophil phagocytosis, chemotaxis, superoxide production, and attenuated degranulation | G-CSF | Improvement in Child–Pugh and MELD scores reduced short-term mortality |
| Immune Cells | Effector Molecules | Action |
|---|---|---|
| Plasma and Memory B cells | IgA, IgG, IgE | Eliminate antigen |
| Th1 | IFNγ, TNF, IL-2, Lymphotoxin | Pro-inflammatory |
| Th2 | IL-4, IL-5, IL-6, IL-10, IL-13 | Pro-inflammatory, Allergy |
| Th9 | IL-9, IL-10, IL-21 | Pro-inflammatory |
| Th17 | IL-17, IL-21, IL-22, IL-25, IL-26 | Pro-inflammatory |
| Th22 | IL-22 | Pro-inflammatory |
| Treg | TGFβ, IL-10, IL-35 | Anti-inflammatory |
| Tfh | IFNγ, STAT3, IL-4, IL-10, IL-21 | Help B cell activation |
| CTL | Perforin, Granzyme, INFγ, TNFα, TNFβ | Tumor cell/ Lyse infected |
| MC | TNFα, IL-1β, IL-6, IL-12, IL-23, MCP1, ROS | Pro-inflammatory |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dąbrowska, A.; Wilczyński, B.; Mastalerz, J.; Kucharczyk, J.; Kulbacka, J.; Szewczyk, A.; Rembiałkowska, N. The Impact of Liver Failure on the Immune System. Int. J. Mol. Sci. 2024, 25, 9522. https://doi.org/10.3390/ijms25179522
Dąbrowska A, Wilczyński B, Mastalerz J, Kucharczyk J, Kulbacka J, Szewczyk A, Rembiałkowska N. The Impact of Liver Failure on the Immune System. International Journal of Molecular Sciences. 2024; 25(17):9522. https://doi.org/10.3390/ijms25179522
Chicago/Turabian StyleDąbrowska, Alicja, Bartosz Wilczyński, Jakub Mastalerz, Julia Kucharczyk, Julita Kulbacka, Anna Szewczyk, and Nina Rembiałkowska. 2024. "The Impact of Liver Failure on the Immune System" International Journal of Molecular Sciences 25, no. 17: 9522. https://doi.org/10.3390/ijms25179522