
Downregulation of IL-8 and IL-10 by LRRC8A Inhibition through the NOX2–Nrf2–CEBPB Transcriptional Axis in THP-1-Derived M2 Macrophages
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
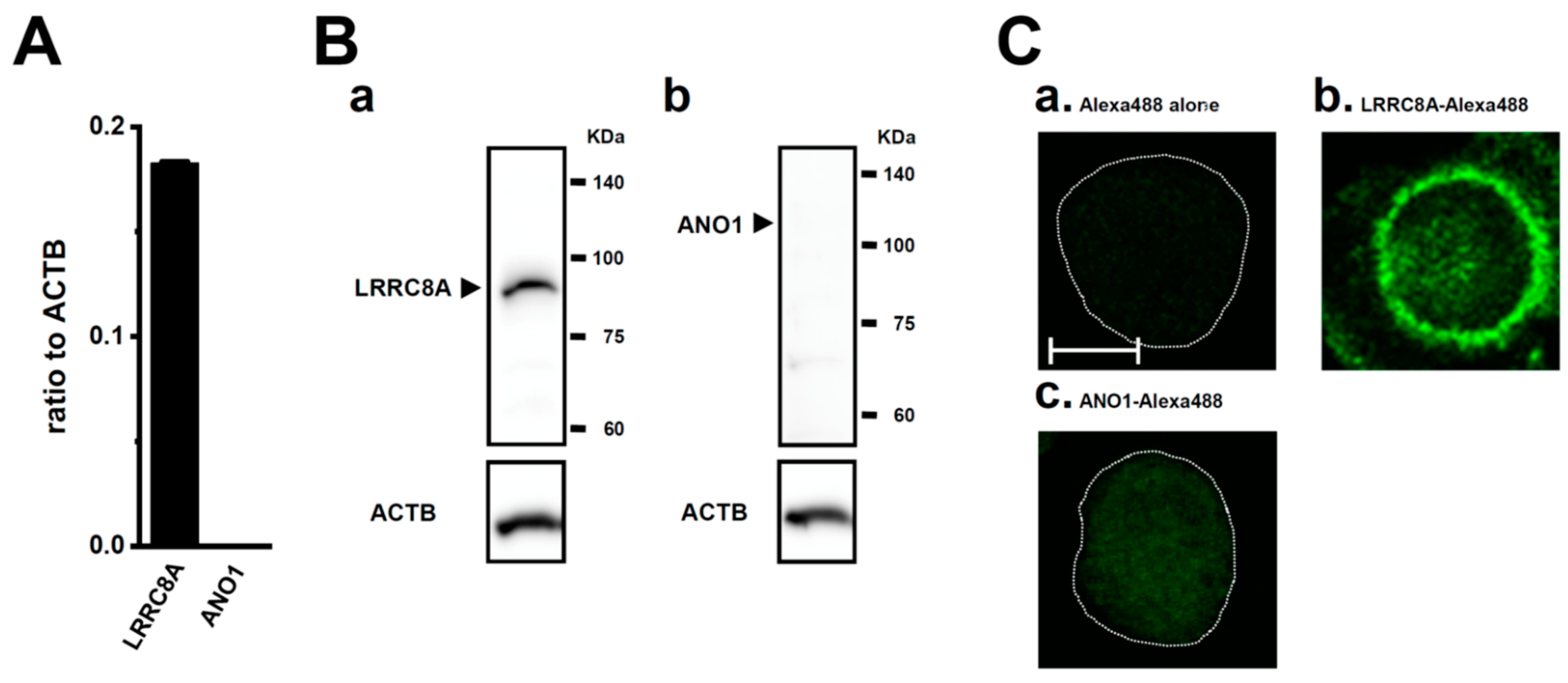
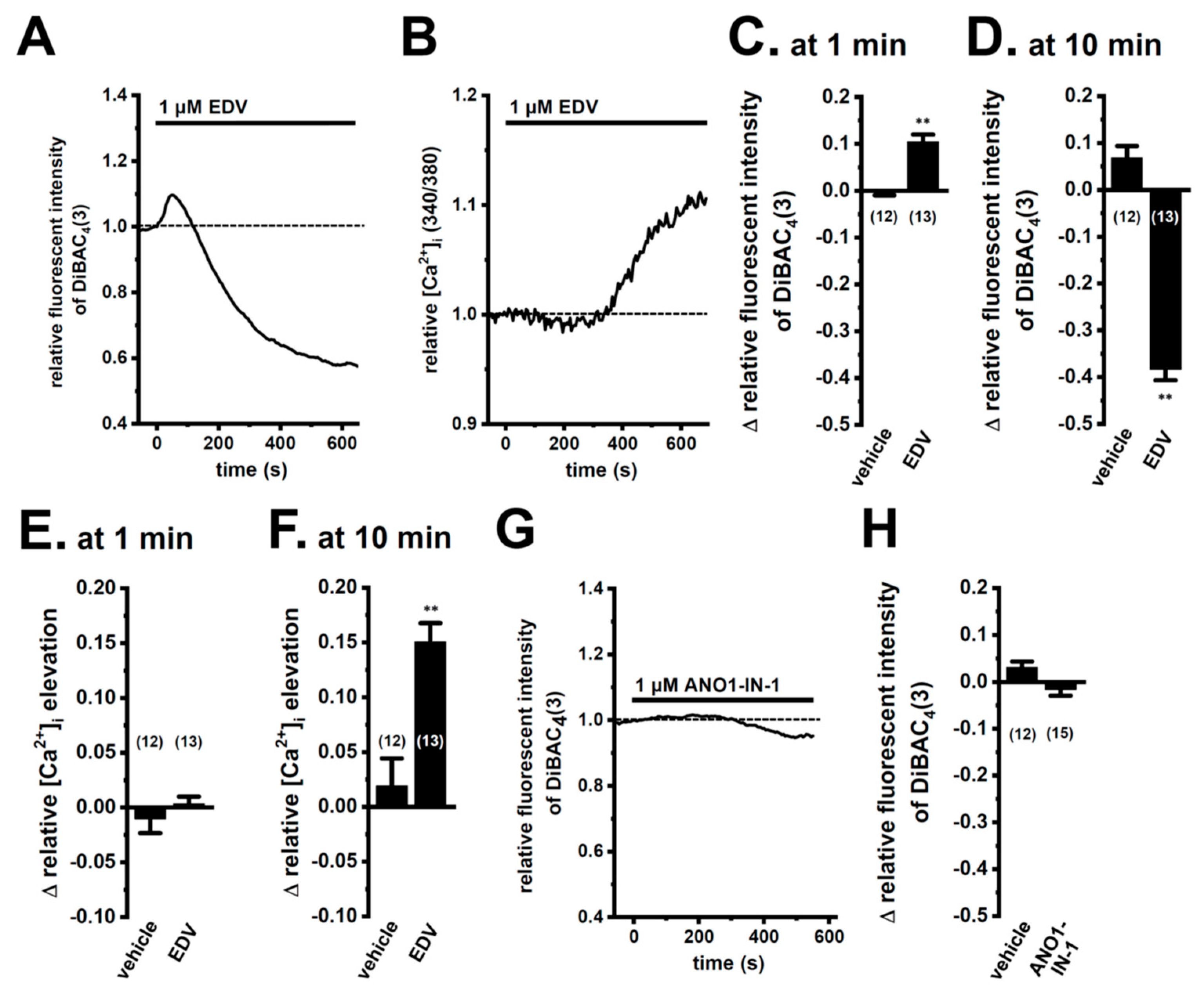
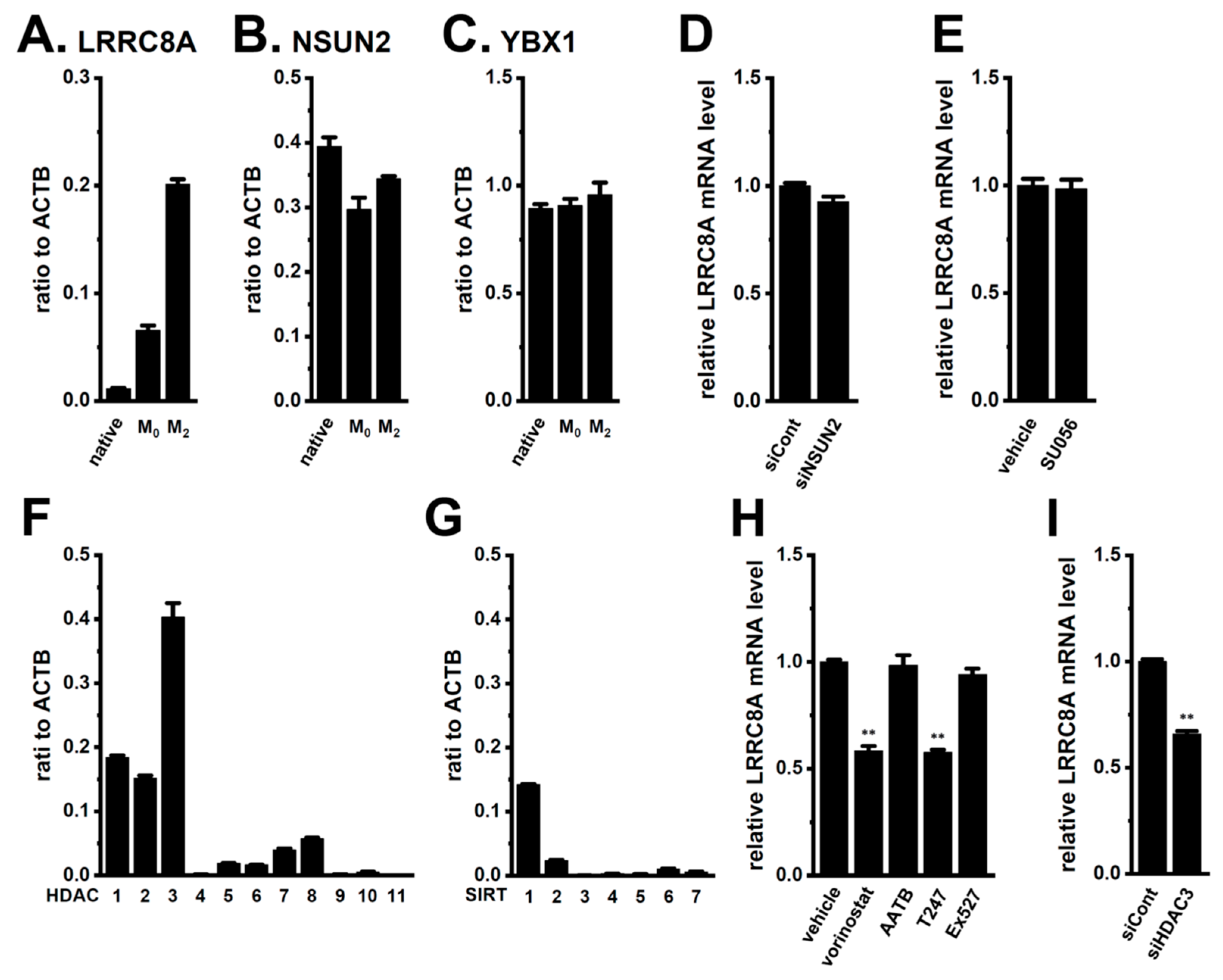
2.1. Functional Expression of LRRC8A in M2-MACs
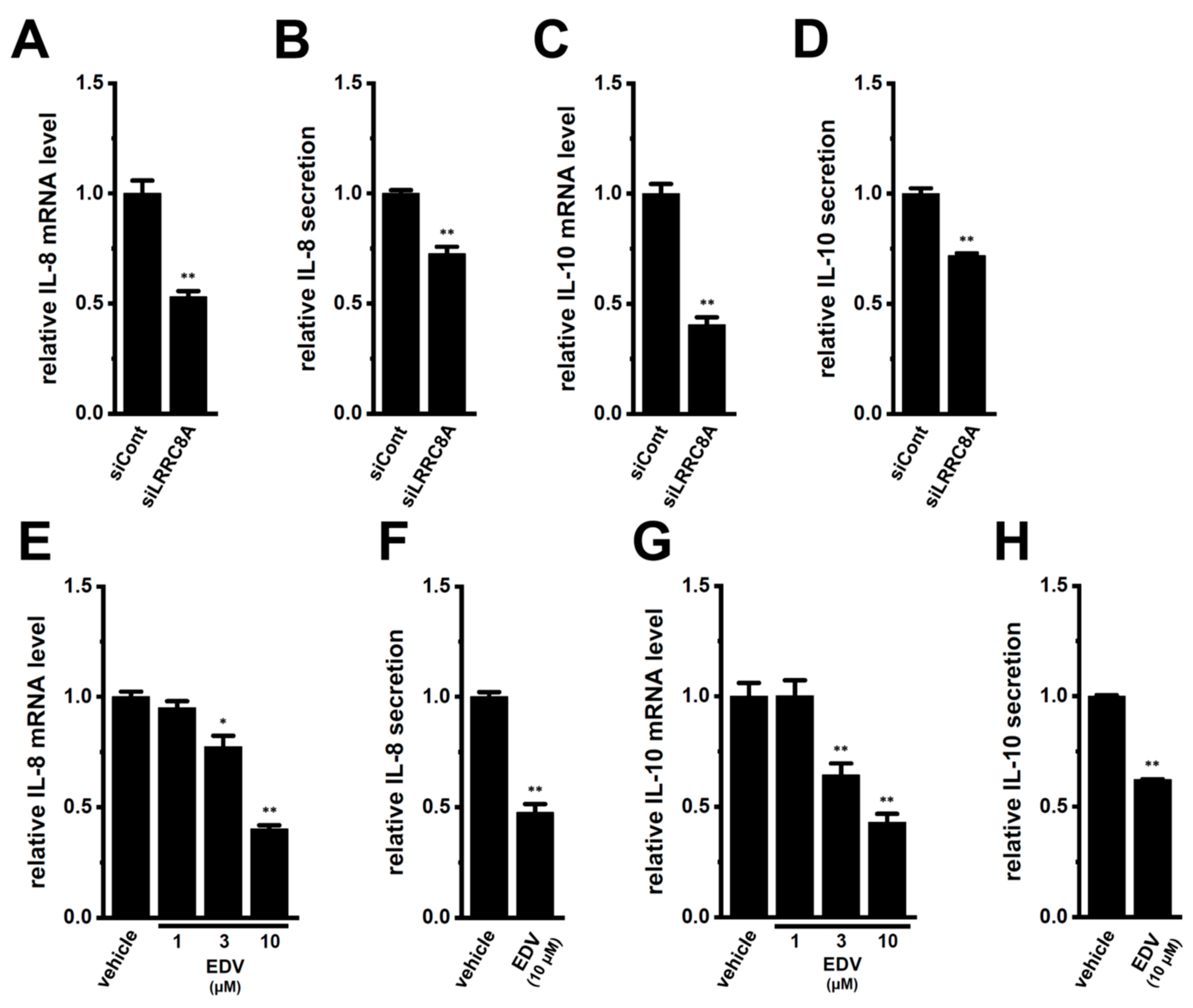
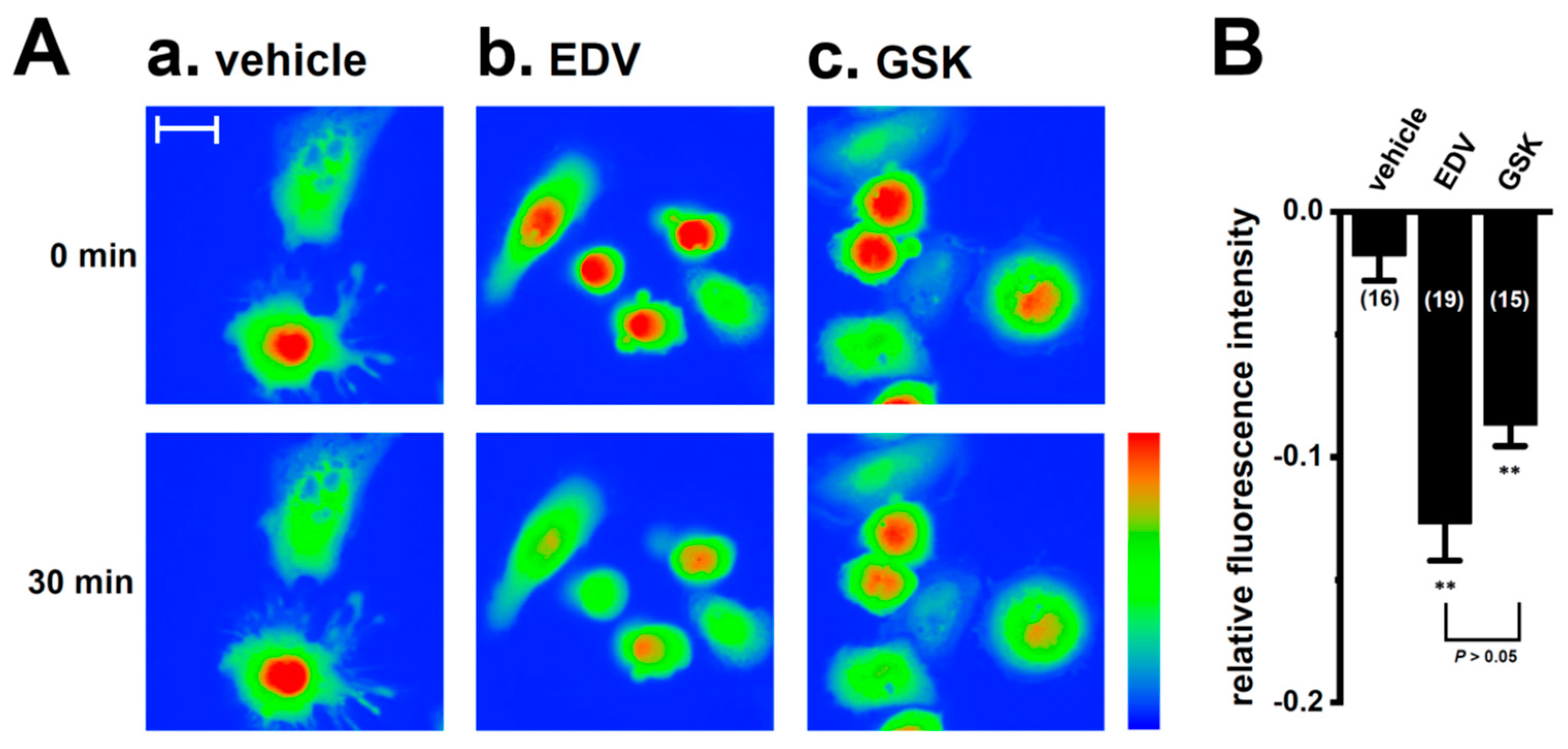
2.2. Decreased Expression and Secretion of IL-8 and IL-10 by the Small Interfering RNA (siRNA)-Mediated and Pharmacological Inhibition of LRRC8A in M2-MACs
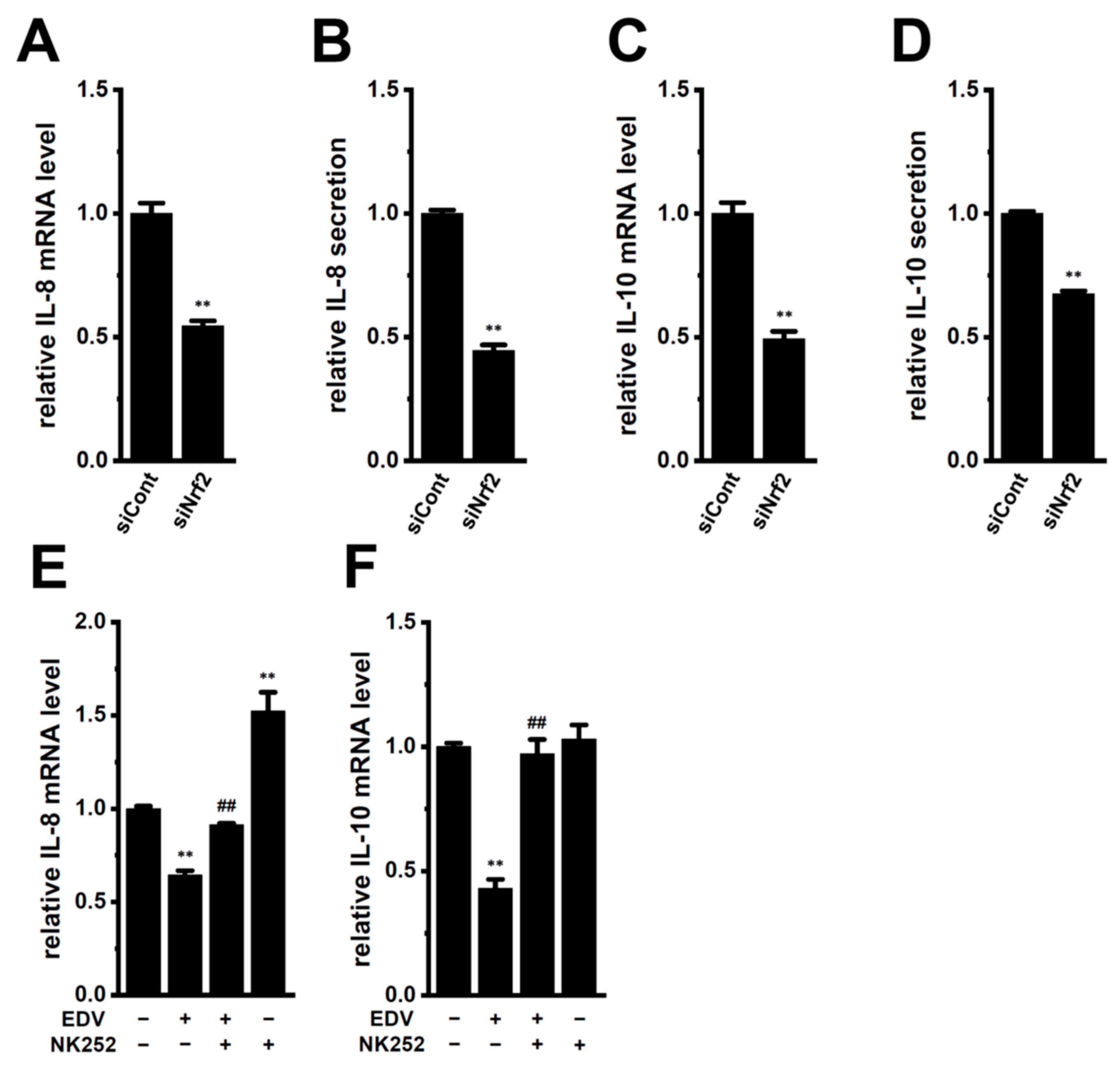
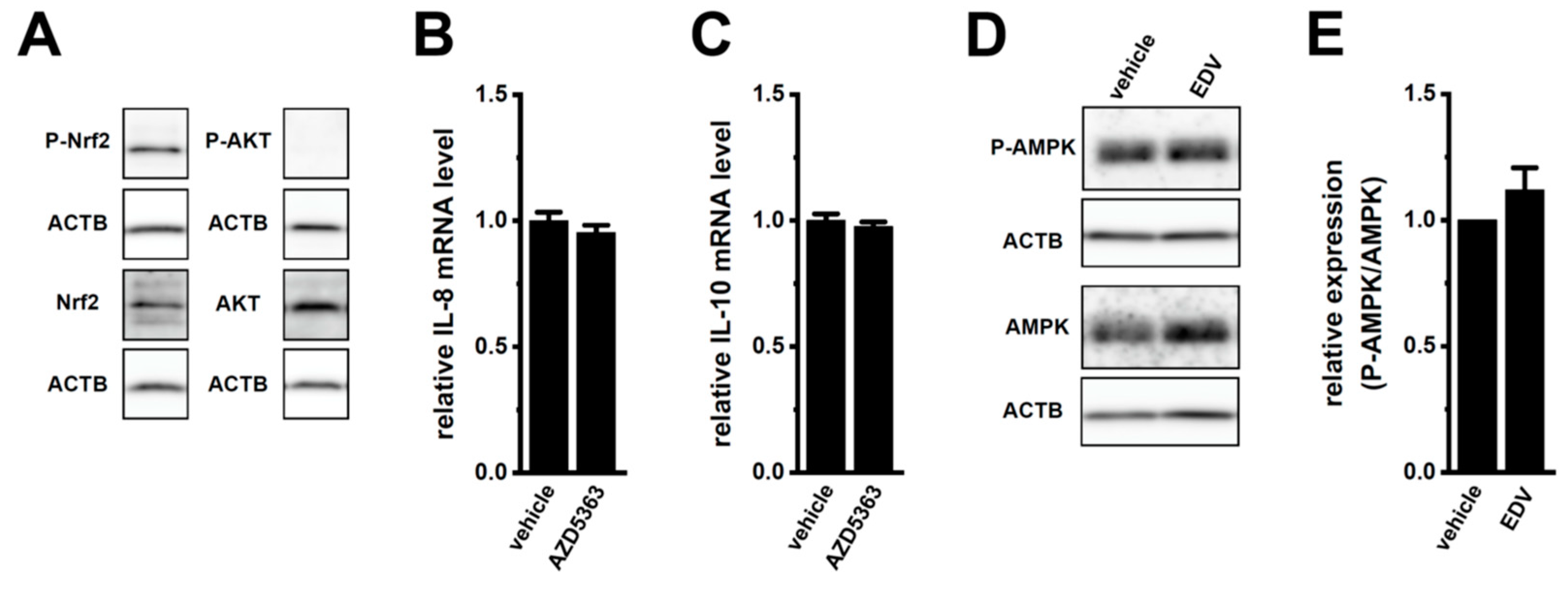
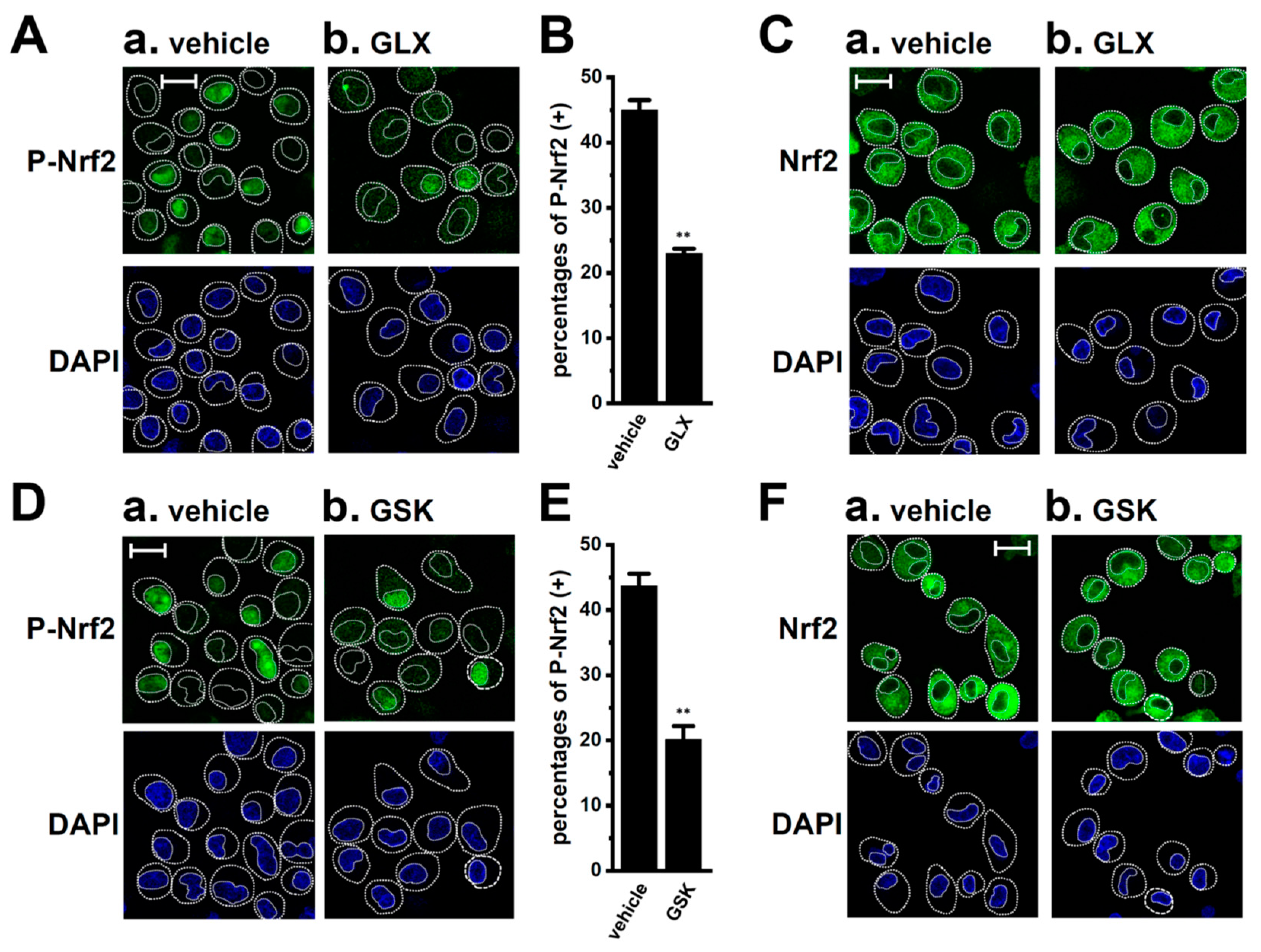
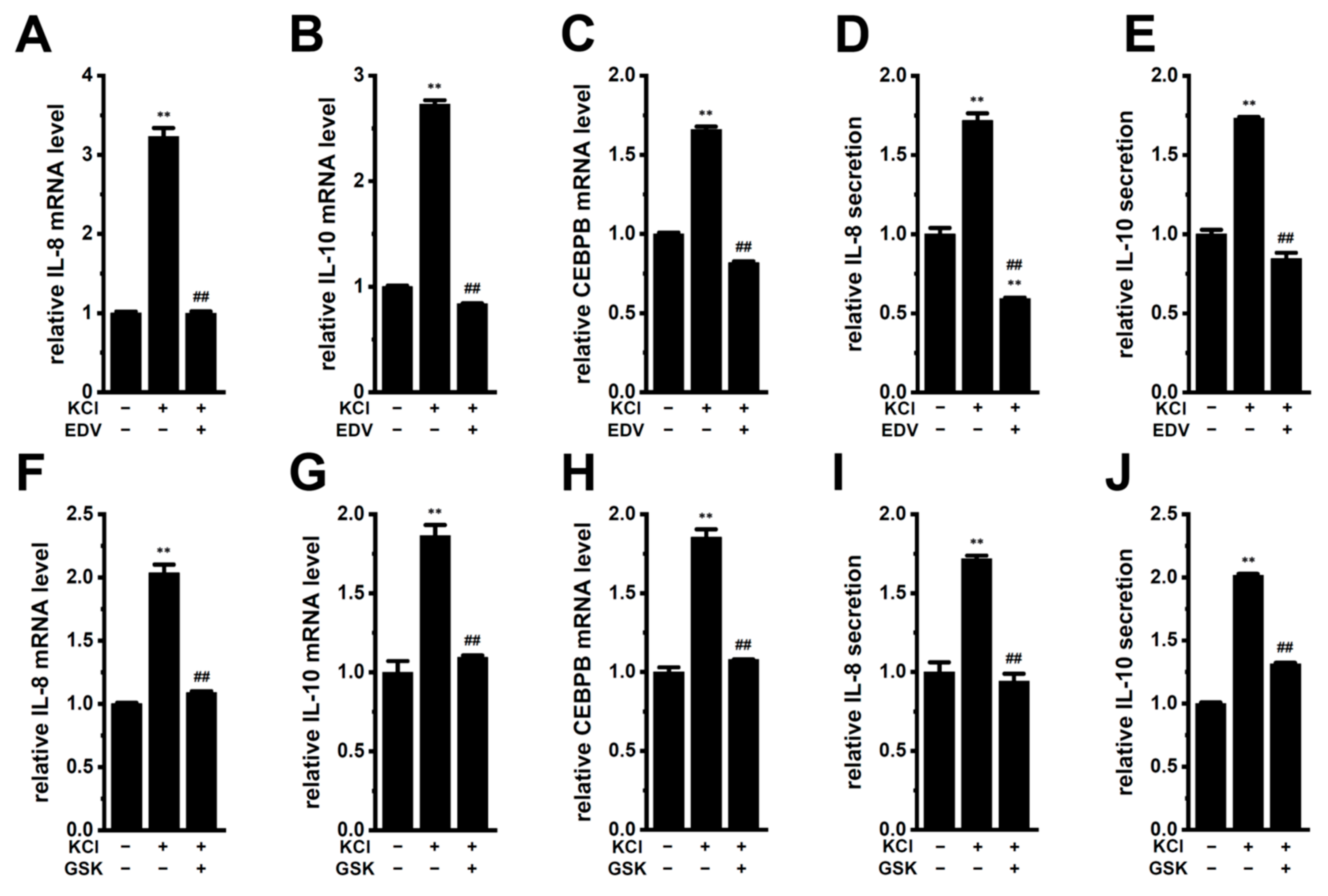
2.3. LRRC8A Inhibition-Induced Transcriptional Repression of IL-8 and IL-10 through the Nrf2 Signaling Pathway in M2-MACs
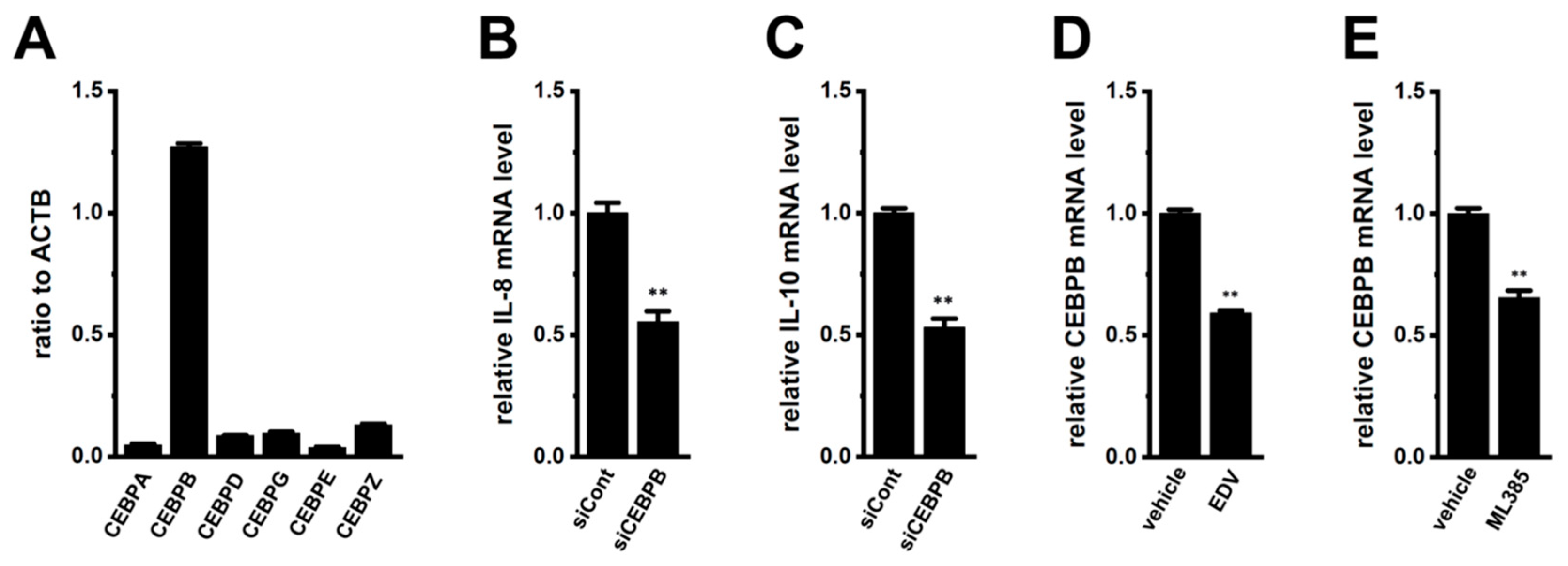
2.4. Involvement of CEBPB in the LRRC8A Inhibition-Induced Transcriptional Repression of IL-8 and IL-10 in M2-MACs
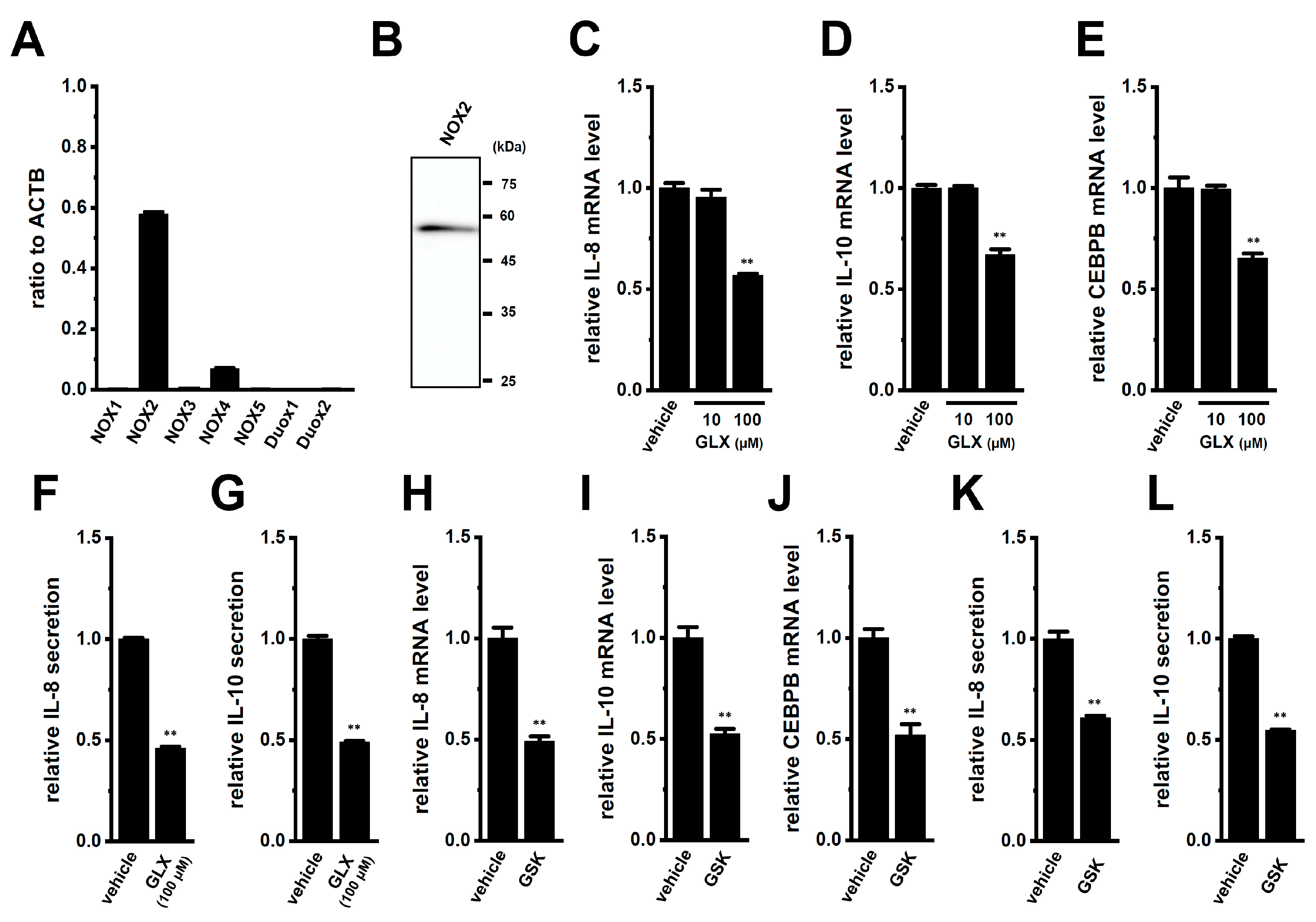
2.5. Involvement of the NOX2-ROS Axis in the LRRC8A Inhibition-Induced Transcriptional Repression of IL-8 and IL-10 in M2-MACs
2.6. Decreased ROS Production by LRRC8A Inhibition in M2-MACs
2.7. Suppression of the High Extracellular K+-Induced Upregulation of IL-8 and IL-10 by LRRC8A and NOX2 Inhibition in M2-MACs
2.8. Mechanisms Underlying the Transcriptional Upregulation of LRRC8A in M2-MACs
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture and Differentiation into M2 Macrophages
4.3. Measurements of Membrane Potential and Intracellular Concentration of Ca2+ ([Ca2+]i)
4.4. Real-Time PCR Assay
4.5. Western Blots
4.6. Measurement of Cytokine Production by ELISA and ROS by a ROS Assay Kit
4.7. Confocal Imaging of the Nuclear and Cytosolic Distribution of Nrf2 and AMPK and the Plasma Membrane Localization of LRRC8A and NOX2
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jentsch, T.J.; Lutter, D.; Planells-Cases, R.; Ullrich, F.; Voss, F.K. VRAC: Molecular identification as LRRC8 heteromers with differential functions. Pflugers Arch. Eur. J. Physiol. 2015, 468, 385–393. [Google Scholar] [CrossRef]
- Okada, Y.; Sabirov, R.Z.; Merzlyak, P.G.; Numata, T.; Sato-Numata, K. Properties, structures, and physiological roles of three types of anion channels molecularly identified in the 2010’s. Front. Physiol. 2021, 12, 805148. [Google Scholar] [CrossRef] [PubMed]
- Prevarskaya, N.; Skryma, R.; Shuba, Y. Ion Channels in Cancer: Are Cancer Hallmarks Oncochannelopathies? Physiol. Rev. 2018, 98, 559–621. [Google Scholar] [CrossRef] [PubMed]
- Vig, M.; Kinet, J.-P. Calcium signaling in immune cells. Nat. Immunol. 2009, 10, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Cersosimo, F.; Lonardi, S.; Bernardini, G.; Telfer, B.; Mandelli, G.E.; Santucci, A.; Vermi, W.; Giurisato, E. Tumor-Associated Macrophages in Osteosarcoma: From Mechanisms to Therapy. Int. J. Mol. Sci. 2020, 21, 5207. [Google Scholar] [CrossRef]
- Lin, Y.; Xu, J.; Lan, H. Tumor-associated macrophages in tumor metastasis: Biological roles and clinical therapeutic applications. J. Hematol. Oncol. 2019, 12, 1–16. [Google Scholar] [CrossRef]
- Mantovani, A.; Allavena, P.; Marchesi, F.; Garlanda, C. Macrophages as tools and targets in cancer therapy. Nat. Rev. Drug Discov. 2022, 21, 799–820. [Google Scholar] [CrossRef]
- DeNardo, D.G.; Ruffell, B. Macrophages as regulators of tumour immunity and immunotherapy. Nat. Rev. Immunol. 2019, 19, 369–382. [Google Scholar] [CrossRef]
- Genin, M.; Clement, F.; Fattaccioli, A.; Raes, M.; Michiels, C. M1 and M2 macrophages derived from THP-1 cells differentially modulate the response of cancer cells to etoposide. BMC Cancer 2015, 15, 577. [Google Scholar] [CrossRef]
- Salmaninejad, A.; Valilou, S.F.; Soltani, A.; Ahmadi, S.; Abarghan, Y.J.; Rosengren, R.J.; Sahebkar, A. Tumor-associated macrophages: Role in cancer development and therapeutic implications. Cell. Oncol. 2019, 42, 591–608. [Google Scholar] [CrossRef]
- Molgora, M.; Colonna, M. Turning enemies into allies—reprogramming tumor-associated macrophages for cancer therapy. Med 2021, 2, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Fousek, K.; Horn, L.A.; Palena, C. Interleukin-8: A chemokine at the intersection of cancer plasticity, angiogenesis, and immune suppression. Pharmacol. Ther. 2020, 219, 107692. [Google Scholar] [CrossRef]
- Han, Z.-J.; Li, Y.-B.; Yang, L.-X.; Cheng, H.-J.; Liu, X.; Chen, H. Roles of the CXCL8-CXCR1/2 Axis in the Tumor Microenvironment and Immunotherapy. Molecules 2021, 27, 137. [Google Scholar] [CrossRef] [PubMed]
- Ohya, S.; Matsui, M.; Kajikuri, J.; Kito, H.; Endo, K. Downregulation of IL-8 and IL-10 by the Activation of Ca2+-Activated K+ Channel KCa3.1 in THP-1-Derived M2 Macrophages. Int. J. Mol. Sci. 2022, 23, 8603. [Google Scholar] [CrossRef]
- Tossetta, G.; Fantone, S.; Marzioni, D.; Mazzucchelli, R. Cellular Modulators of the NRF2/KEAP1 Signaling Pathway in Prostate Cancer. Front. Biosci. 2023, 28, 143. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Zhang, F.; Meng, D.; Zhang, X.; Feng, Y.; Yin, G.; Liang, P.; Chen, S.; Liu, H. Mechanism of Action and Related Natural Regulators of Nrf2 in Nonalcoholic Fatty Liver Disease. Curr. Drug Deliv. 2024, 21, 1300–1319. [Google Scholar] [CrossRef]
- Wang, L.; He, C. Nrf2-mediated anti-inflammatory polarization of macrophages as therapeutic targets for osteoarthritis. Front. Immunol. 2022, 13, 967193. [Google Scholar] [CrossRef]
- Wang, Y.-R.; Zhang, X.-N.; Meng, F.-G.; Zeng, T. Targeting macrophage polarization by Nrf2 agonists for treating various xenobiotics-induced toxic responses. Toxicol. Mech. Methods 2021, 31, 334–342. [Google Scholar] [CrossRef]
- Feng, R.; Morine, Y.; Ikemoto, T.; Imura, S.; Iwahashi, S.; Saito, Y.; Shimada, M. Nrf2 activation drive macrophages polarization and cancer cell epithelial-mesenchymal transition during interaction. Cell Commun. Signal. 2018, 16, 1–12. [Google Scholar] [CrossRef]
- Liu, J.; Shen, D.; Wei, C.; Wu, W.; Luo, Z.; Hu, L.; Xiao, Z.; Hu, T.; Sun, Q.; Wang, X.; et al. Inhibition of the LRRC8A channel promotes microglia/macrophage phagocytosis and improves outcomes after intracerebral hemorrhagic stroke. iScience 2022, 25, 105527. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, X.; Song, H.; Chen, H.-Z.; Rovin, B.H. Activation of the Nrf2/antioxidant response pathway increases IL-8 expression. Eur. J. Immunol. 2005, 35, 3258–3267. [Google Scholar] [CrossRef]
- Härkönen, J.; Pölönen, P.; Deen, A.J.; Selvarajan, I.; Teppo, H.-R.; Dimova, E.Y.; Kietzmann, T.; Ahtiainen, M.; Väyrynen, J.P.; Väyrynen, S.A.; et al. A pan-cancer analysis shows immunoevasive characteristics in NRF2 hyperactive squamous malignancies. Redox Biol. 2023, 61, 102644. [Google Scholar] [CrossRef]
- Kyung, S.; Lim, J.W.; Kim, H. α-Lipoic Acid Inhibits IL-8 Expression by Activating Nrf2 Signaling in Helicobacter pylori-infected Gastric Epithelial Cells. Nutrients 2019, 11, 2524. [Google Scholar] [CrossRef]
- Subedi, L.; Lee, J.H.; Yumnam, S.; Ji, E.; Kim, S.Y. Anti-Inflammatory Effect of Sulforaphane on LPS-Activated Microglia Potentially through JNK/AP-1/NF-κB Inhibition and Nrf2/HO-1 Activation. Cells 2019, 8, 194. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Yuan, X.; Yan, J.; Huang, Y.; Xu, M.; Yang, Z.; Yang, N.; Wang, M.; Zhang, C.; Zhang, L. Nrf2 exerts mixed inflammation and glucose metabolism regulatory effects on murine RAW264.7 macrophages. Int. Immunopharmacol. 2019, 71, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, L.; Naik, I.; Braunstein, Z.; Zhong, J.; Ren, B. Transcription Factor C/EBP Homologous Protein in Health and Diseases. Front. Immunol. 2017, 8, 1612. [Google Scholar] [CrossRef] [PubMed]
- Ruffell, D.; Mourkioti, F.; Gambardella, A.; Kirstetter, P.; Lopez, R.G.; Rosenthal, N.; Nerlov, C. A CREB-C/EBPβ cascade induces M2 macrophage-specific gene expression and promotes muscle injury repair. Proc. Natl. Acad. Sci. USA 2009, 106, 17475–17480. [Google Scholar] [CrossRef] [PubMed]
- Venza, M.; Visalli, M.; Alafaci, C.; Caffo, M.; Caruso, G.; Salpietro, F.M.; Tomasello, F.; Teti, D. Interleukin-8 Overexpression in Astrocytomas Is Induced by Prostaglandin E2 and Is Associated With the Transcription Factors CCAAT/Enhancer-Binding Protein-β and CCAAT/Enhancer-Binding Homologous Protein. Neurosurgery 2011, 69, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S.; Prösch, S.; Schenke-Layland, K.; Riese, U.; Gausmann, U.; Platzer, C. cAMP-induced Interleukin-10 Promoter Activation Depends on CCAAT/Enhancer-binding Protein Expression and Monocytic Differentiation. J. Biol. Chem. 2003, 278, 5597–5604. [Google Scholar] [CrossRef]
- Zhou, J.; Li, H.; Xia, X.; Herrera, A.; Pollock, N.; Reebye, V.; Sodergren, M.H.; Dorman, S.; Littman, B.H.; Doogan, D.; et al. Anti-inflammatory Activity of MTL-CEBPA, a Small Activating RNA Drug, in LPS-Stimulated Monocytes and Humanized Mice. Mol. Ther. 2019, 27, 999–1016. [Google Scholar] [CrossRef]
- Huo, C.; Liu, Y.; Li, X.; Xu, R.; Jia, X.; Hou, L.; Wang, X. LRRC8A contributes to angiotensin II-induced cardiac hypertrophy by interacting with NADPH oxidases via the C-terminal leucine-rich repeat domain. Free. Radic. Biol. Med. 2021, 165, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.-Y.; Kim, S.; Park, H.-M.; Lim, C.-M.; Kim, J.; Park, J.-Y.; Jeon, K.-B.; Poudel, A.; Lee, H.P.; Oh, S.-R.; et al. Cinnamomum verum extract inhibits NOX2/ROS and PKCδ/JNK/AP-1/NF-κB pathway-mediated inflammatory response in PMA-stimulated THP-1 monocytes. Phytomedicine 2023, 112, 154685. [Google Scholar] [CrossRef]
- Hu, C.-F.; Wu, S.-P.; Lin, G.-J.; Shieh, C.-C.; Hsu, C.-S.; Chen, J.-W.; Chen, S.-H.; Hong, J.-S.; Chen, S.-J. Microglial Nox2 Plays a Key Role in the Pathogenesis of Experimental Autoimmune Encephalomyelitis. Front. Immunol. 2021, 12, 638381. [Google Scholar] [CrossRef] [PubMed]
- Virág, L.; Jaén, R.I.; Regdon, Z.; Boscá, L.; Prieto, P. Self-defense of macrophages against oxidative injury: Fighting for their own survival. Redox Biol. 2019, 26, 101261. [Google Scholar] [CrossRef]
- Lee, G.H.; Kim, J.Y.; Jin, S.W.; Pham, T.H.; Park, J.S.; Kim, C.Y.; Choi, J.H.; Han, E.H.; Kim, Y.H.; Jeong, H.G. Impressic Acid Attenuates the Lipopolysaccharide-Induced Inflammatory Response by Activating the AMPK/GSK3β/Nrf2 Axis in RAW264.7 Macrophages. Int. J. Mol. Sci. 2021, 22, 762. [Google Scholar] [CrossRef]
- Li, S.-T.; Dai, Q.; Zhang, S.-X.; Liu, Y.-J.; Yu, Q.-Q.; Tan, F.; Lu, S.-H.; Wang, Q.; Chen, J.-W.; Huang, H.-Q.; et al. Ulinastatin attenuates LPS-induced inflammation in mouse macrophage RAW264.7 cells by inhibiting the JNK/NF-κB signaling pathway and activating the PI3K/Akt/Nrf2 pathway. Acta Pharmacol. Sin. 2018, 39, 1294–1304. [Google Scholar] [CrossRef]
- Cui, Y.; Chen, J.; Zhang, Z.; Shi, H.; Sun, W.; Yi, Q. The role of AMPK in macrophage metabolism, function and polarisation. J. Transl. Med. 2023, 21, 1–16. [Google Scholar] [CrossRef]
- Liu, B.; Deng, X.; Jiang, Q.; Li, G.; Zhang, J.; Zhang, N.; Xin, S.; Xu, K. Scoparone improves hepatic inflammation and autophagy in mice with nonalcoholic steatohepatitis by regulating the ROS/P38/Nrf2 axis and PI3K/AKT/mTOR pathway in macrophages. Biomed. Pharmacother. 2020, 125, 109895. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Rohrbough, J.C.; Nguyen, H.N.; Dikalova, A.; Lamb, F.S. Oxidant-resistant LRRC8A/C anion channels support superoxide production by NADPH oxidase. J. Physiol. 2021, 599, 3013–3036. [Google Scholar] [CrossRef]
- Zhen, J.; Chen, X.; Mao, Y.; Xie, X.; Chen, X.; Xu, W.; Zhang, S. GLX351322, a Novel NADPH Oxidase 4 Inhibitor, Attenuates TMJ Osteoarthritis by Inhibiting the ROS/MAPK/NF-κB Signaling Pathways. Oxidative Med. Cell. Longev. 2023, 2023, 1–13. [Google Scholar] [CrossRef]
- Hirano, K.; Chen, W.S.; Chueng, A.L.; Dunne, A.A.; Seredenina, T.; Filippova, A.; Ramachandran, S.; Bridges, A.; Chaudry, L.; Pettman, G.; et al. Discovery of GSK2795039, a Novel Small Molecule NADPH Oxidase 2 Inhibitor. Antioxidants Redox Signal. 2015, 23, 358–374. [Google Scholar] [CrossRef] [PubMed]
- Eil, R.; Vodnala, S.K.; Clever, D.; Klebanoff, C.A.; Sukumar, M.; Pan, J.H.; Palmer, D.C.; Gros, A.; Yamamoto, T.N.; Patel, S.J.; et al. Ionic immune suppression within the tumour microenvironment limits T cell effector function. Nature 2016, 537, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.T.; Ng, A.S.; Ng, X.R.; Zhuang, Z.; Wong, B.H.S.; Prasannan, P.; Kok, Y.J.; Bi, X.; Shim, H.; Wulff, H.; et al. Extracellular K+Dampens T Cell Functions: Implications for Immune Suppression in the Tumor Microenvironment. Bioelectricity 2019, 1, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zuo, X.; Wei, Q.; Xu, J.; Liu, X.; Liu, S.; Wang, H.; Luo, Q.; Wang, Y.; Yang, Y.; et al. Upregulation of LRRC8A by m5C modification-mediated mRNA stability suppresses apoptosis and facilitates tumorigenesis in cervical cancer. Int. J. Biol. Sci. 2023, 19, 691–704. [Google Scholar] [CrossRef]
- Yao, Y.; Liu, Q.; Adrianto, I.; Wu, X.; Glassbrook, J.; Khalasawi, N.; Yin, C.; Yi, Q.; Dong, Z.; Geissmann, F.; et al. Histone deacetylase 3 controls lung alveolar macrophage development and homeostasis. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Matsuba, S.; Niwa, S.; Muraki, K.; Kanatsuka, S.; Nakazono, Y.; Hatano, N.; Fujii, M.; Zhan, P.; Suzuki, T.; Ohya, S. Downregulation of Ca2+-Activated Cl− Channel TMEM16A by the Inhibition of Histone Deacetylase in TMEM16A-Expressing Cancer Cells. J. Pharmacol. Exp. Ther. 2014, 351, 510–518. [Google Scholar] [CrossRef]
- Ohya, S.; Kanatsuka, S.; Hatano, N.; Kito, H.; Matsui, A.; Fujimoto, M.; Matsuba, S.; Niwa, S.; Zhan, P.; Suzuki, T.; et al. Downregulation of the Ca2+-activated K+ channel KCa3.1 by histone deacetylase inhibition in human breast cancer cells. Pharmacol. Res. Perspect. 2016, 4, e00228. [Google Scholar] [CrossRef]
- Choi, H.; Miller, M.R.; Nguyen, H.; Rohrbough, J.C.; Koch, S.R.; Boatwright, N.; Yarboro, M.T.; Sah, R.; McDonald, W.H.; Reese, J.J.; et al. LRRC8A anion channels modulate vascular reactivity via association with myosin phosphatase rho interacting protein. FASEB J. 2023, 37, e23028. [Google Scholar] [CrossRef]
- Pérez, S.; Rius-Pérez, S. Macrophage Polarization and Reprogramming in Acute Inflammation: A Redox Perspective. Antioxidants 2022, 11, 1394. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Becker, T.M.; Koch, U.; Stauber, T. The LRRC8/VRAC anion channel facilitates myogenic differentiation of murine myoblasts by promoting membrane hyperpolarization. J. Biol. Chem. 2019, 294, 14279–14288. [Google Scholar] [CrossRef] [PubMed]
- Molteni, L.; Rizzi, L.; Bresciani, E.; Meanti, R.; Fehrentz, J.-A.; Verdié, P.; Omeljaniuk, R.J.; Biagini, G.; Locatelli, V.; Torsello, A. STIM Proteins and Orai Ca2+ Channels Are Involved in the Intracellular Pathways Activated by TLQP-21 in RAW264.7 Macrophages. Front. Pharmacol. 2018, 9, 1386. [Google Scholar] [CrossRef] [PubMed]
- Demaurex, N.; Nunes, P. The role of STIM and ORAI proteins in phagocytic immune cells. Am. J. Physiol. Physiol. 2016, 310, C496–C508. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, Z.; Ping, J.; Tang, J.; He, B.; Chang, T.; Zhou, Q.; Yuan, S.; Tang, Z.; Li, X.; et al. Cell volume controlled by LRRC8A-formed volume-regulated anion channels fine-tunes T cell activation and function. Nat. Commun. 2023, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Fan, T.; Geng, W.; Chen, Y.H.; Ruan, Q.; Zhang, C. Negative Immune Regulator TIPE2 Promotes M2 Macrophage Differentiation through the Activation of PI3K-AKT Signaling Pathway. PLoS ONE 2017, 12, e0170666. [Google Scholar] [CrossRef]
- Li, S.; Lavagnino, Z.; Lemacon, D.; Kong, L.; Ustione, A.; Ng, X.; Zhang, Y.; Wang, Y.; Zheng, B.; Piwnica-Worms, H.; et al. Ca2+-Stimulated AMPK-Dependent Phosphorylation of Exo1 Protects Stressed Replication Forks from Aberrant Resection. Mol. Cell 2019, 74, 1123–1137.e6. [Google Scholar] [CrossRef]
- Hou, C.-Y.; Ma, C.-Y.; Yuh, C.-H. WNK1 kinase signaling in metastasis and angiogenesis. Cell. Signal. 2022, 96, 110371. [Google Scholar] [CrossRef] [PubMed]
- Friedel, P.; Kahle, K.T.; Zhang, J.; Hertz, N.; Pisella, L.I.; Buhler, E.; Schaller, F.; Duan, J.; Khanna, A.R.; Bishop, P.N.; et al. WNK1-regulated inhibitory phosphorylation of the KCC2 cotransporter maintains the depolarizing action of GABA in immature neurons. Sci. Signal. 2015, 8, ra65. [Google Scholar] [CrossRef]
- Guragain, D.; Gurung, P.; Chang, J.-H.; Katila, N.; Chang, H.W.; Jeong, B.-S.; Choi, D.-Y.; Kim, J.-A. AMPK is essential for IL-10 expression and for maintaining balance between inflammatory and cytoprotective signaling. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2020, 1864, 129631. [Google Scholar] [CrossRef]
- Matzinger, M.; Fischhuber, K.; Pölöske, D.; Mechtler, K.; Heiss, E.H. AMPK leads to phosphorylation of the transcription factor Nrf2, tuning transactivation of selected target genes. Redox Biol. 2020, 29, 101393. [Google Scholar] [CrossRef]
- Glorieux, C.; Enríquez, C.; González, C.; Aguirre-Martínez, G.; Calderon, P.B. The Multifaceted Roles of NRF2 in Cancer: Friend or Foe? Antioxidants 2024, 13, 70. [Google Scholar] [CrossRef]
- Li, C.; Xu, M.M.; Wang, K.; Adler, A.J.; Vella, A.T.; Zhou, B. Macrophage polarization and meta-inflammation. Transl. Res. 2017, 191, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Wu, M.; Zeng, L.; Wang, D. The Beneficial Role of Nrf2 in the Endothelial Dysfunction of Atherosclerosis. Cardiol. Res. Pract. 2022, 2022, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Le, Y.; Ding, J.; Dou, X.; Mao, W.; Zhu, J. CLIC1 Inhibition Protects Against Cellular Senescence and Endothelial Dysfunction Via the Nrf2/HO-1 Pathway. Cell Biochem. Biophys. 2021, 79, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Salao, K.; Jiang, L.; Li, H.; Tsai, V.W.-W.; Husaini, Y.; Curmi, P.M.G.; Brown, L.J.; Brown, D.A.; Breit, S.N. CLIC1 regulates dendritic cell antigen processing and presentation by modulating phagosome acidification and proteolysis. Biol. Open 2016, 5, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Gaffney, S.G.; Cannataro, V.L.; Townsend, J. Transfer RNA methyltransferase gene NSUN2 mRNA expression modifies the effect of T cell activation score on patient survival in head and neck squamous carcinoma. Oral Oncol. 2019, 101, 104554. [Google Scholar] [CrossRef]
- Li, N.; Liu, B.; He, R.; Li, G.; Xiong, R.; Fu, T.; Li, D.; Xu, C.; Wang, B.; Geng, Q. HDAC3 promotes macrophage pyroptosis via regulating histone deacetylation in acute lung injury. iScience 2023, 26, 107158. [Google Scholar] [CrossRef]
- Wang, S.; Wang, J.; Chen, Z.; Luo, J.; Guo, W.; Sun, L.; Lin, L. Targeting M2-like tumor-associated macrophages is a potential therapeutic approach to overcome antitumor drug resistance. npj Precis. Oncol. 2024, 8, 1–19. [Google Scholar] [CrossRef]
- Carpanese, V.; Festa, M.; Prosdocimi, E.; Bachmann, M.; Sadeghi, S.; Bertelli, S.; Stein, F.; Velle, A.; Abdel-Salam, M.A.L.; Romualdi, C.; et al. Interactomic exploration of LRRC8A in volume-regulated anion channels. Cell Death Discov. 2024, 10, 1–28. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsui, M.; Kajikuri, J.; Kito, H.; Elboray, E.E.; Suzuki, T.; Ohya, S. Downregulation of IL-8 and IL-10 by LRRC8A Inhibition through the NOX2–Nrf2–CEBPB Transcriptional Axis in THP-1-Derived M2 Macrophages. Int. J. Mol. Sci. 2024, 25, 9612. https://doi.org/10.3390/ijms25179612
Matsui M, Kajikuri J, Kito H, Elboray EE, Suzuki T, Ohya S. Downregulation of IL-8 and IL-10 by LRRC8A Inhibition through the NOX2–Nrf2–CEBPB Transcriptional Axis in THP-1-Derived M2 Macrophages. International Journal of Molecular Sciences. 2024; 25(17):9612. https://doi.org/10.3390/ijms25179612
Chicago/Turabian StyleMatsui, Miki, Junko Kajikuri, Hiroaki Kito, Elghareeb E. Elboray, Takayoshi Suzuki, and Susumu Ohya. 2024. "Downregulation of IL-8 and IL-10 by LRRC8A Inhibition through the NOX2–Nrf2–CEBPB Transcriptional Axis in THP-1-Derived M2 Macrophages" International Journal of Molecular Sciences 25, no. 17: 9612. https://doi.org/10.3390/ijms25179612