
UTRs and Ago-2/miR-335 Complex Restricts Amylin Translation in Insulinoma and Human Pancreatic β-Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
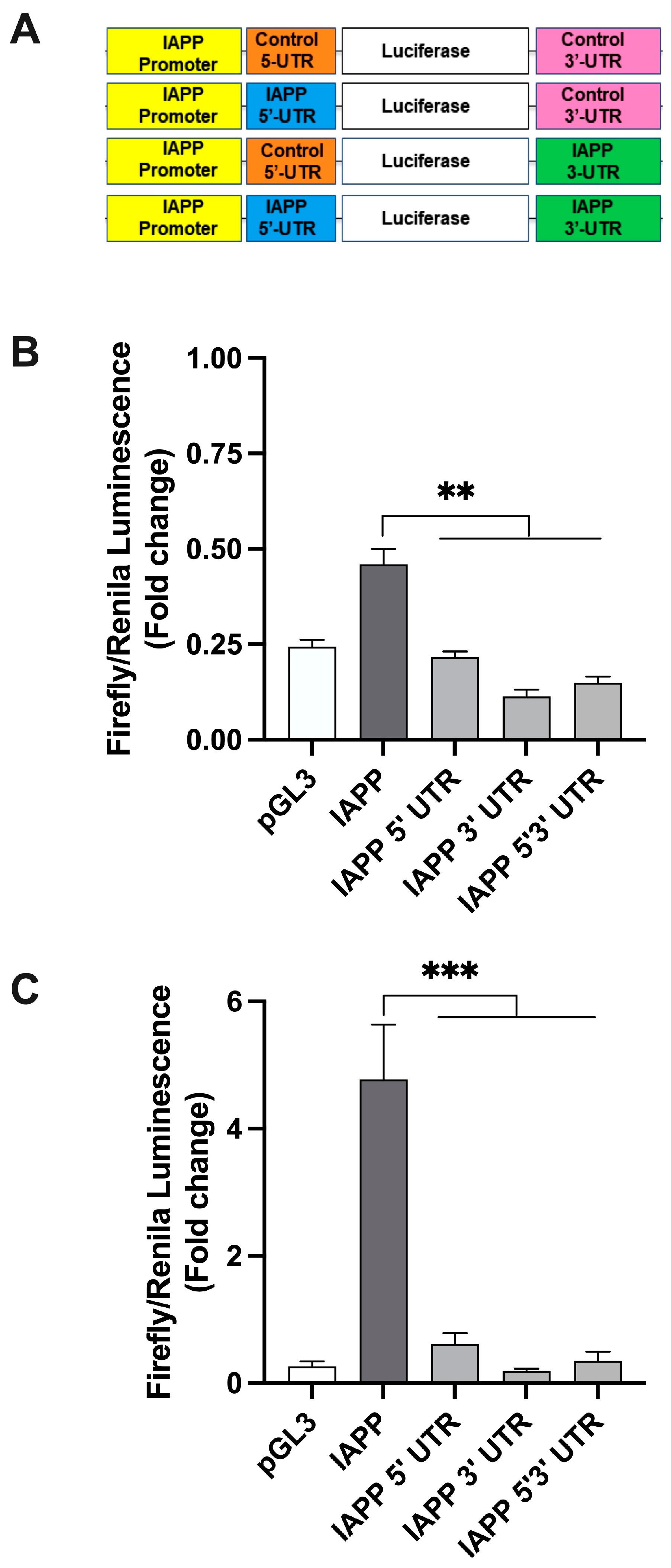
2.1. hIAPP 5′ and 3′ UTR Independently Regulate Protein Translation in Rat Insulinoma Cells
2.2. hIAPP UTRs and SV40 Promoter Do Not Restrict Protein Translation in Insulinoma β-Cells
2.3. 3’UTR of hIAPP Contains Seed Sequences for Several T2DM-Relevant miRNAs
2.4. miR-182, miR-335, and miR-495 Levels Are Downregulated in Stressed Human Islets
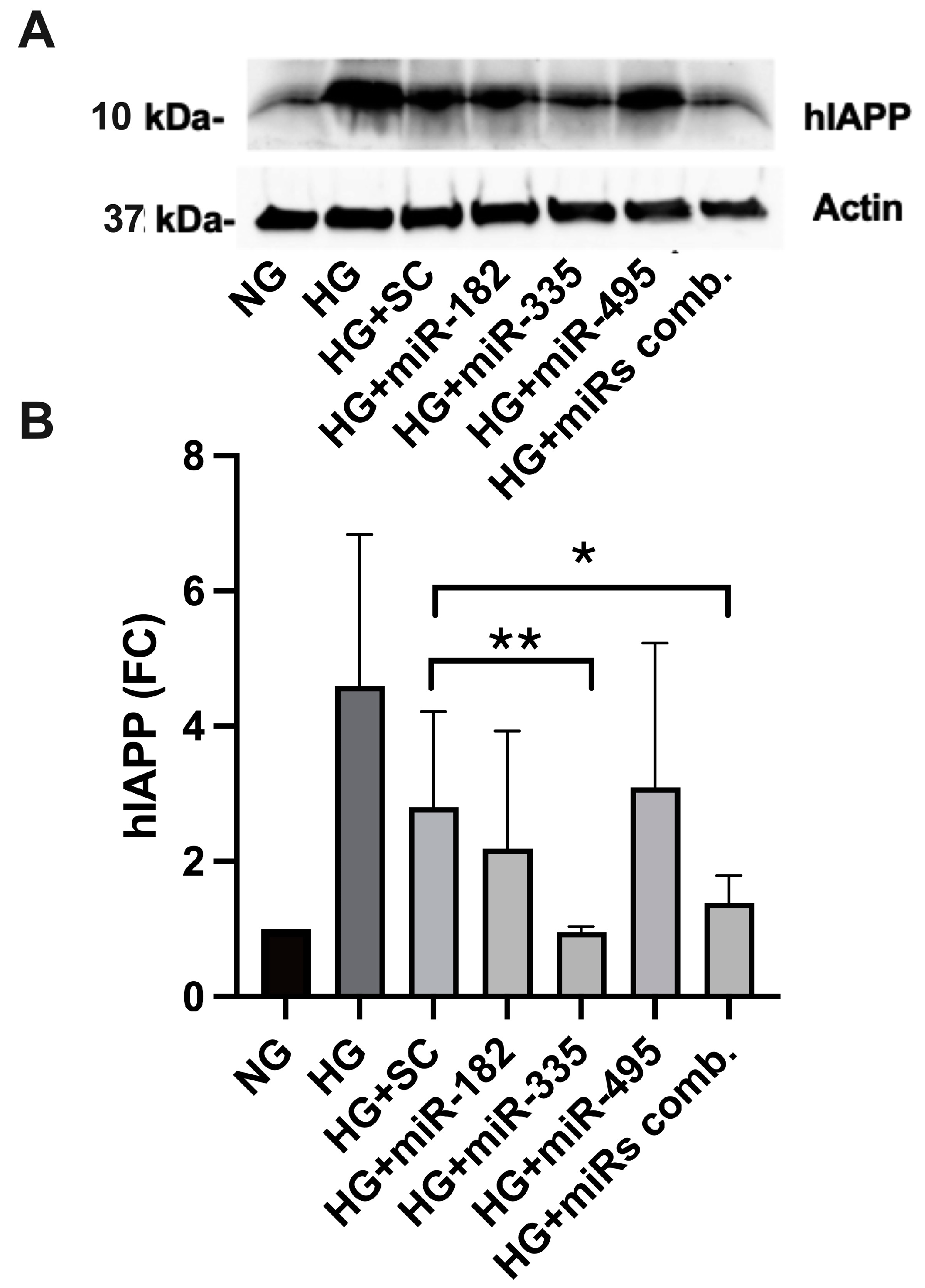
2.5. miR-Mimics Complementary to hIAPP 3′ UTR Seed Sequences Confine hIAPP Protein Expression in HG-Treated Cultured Human Islets
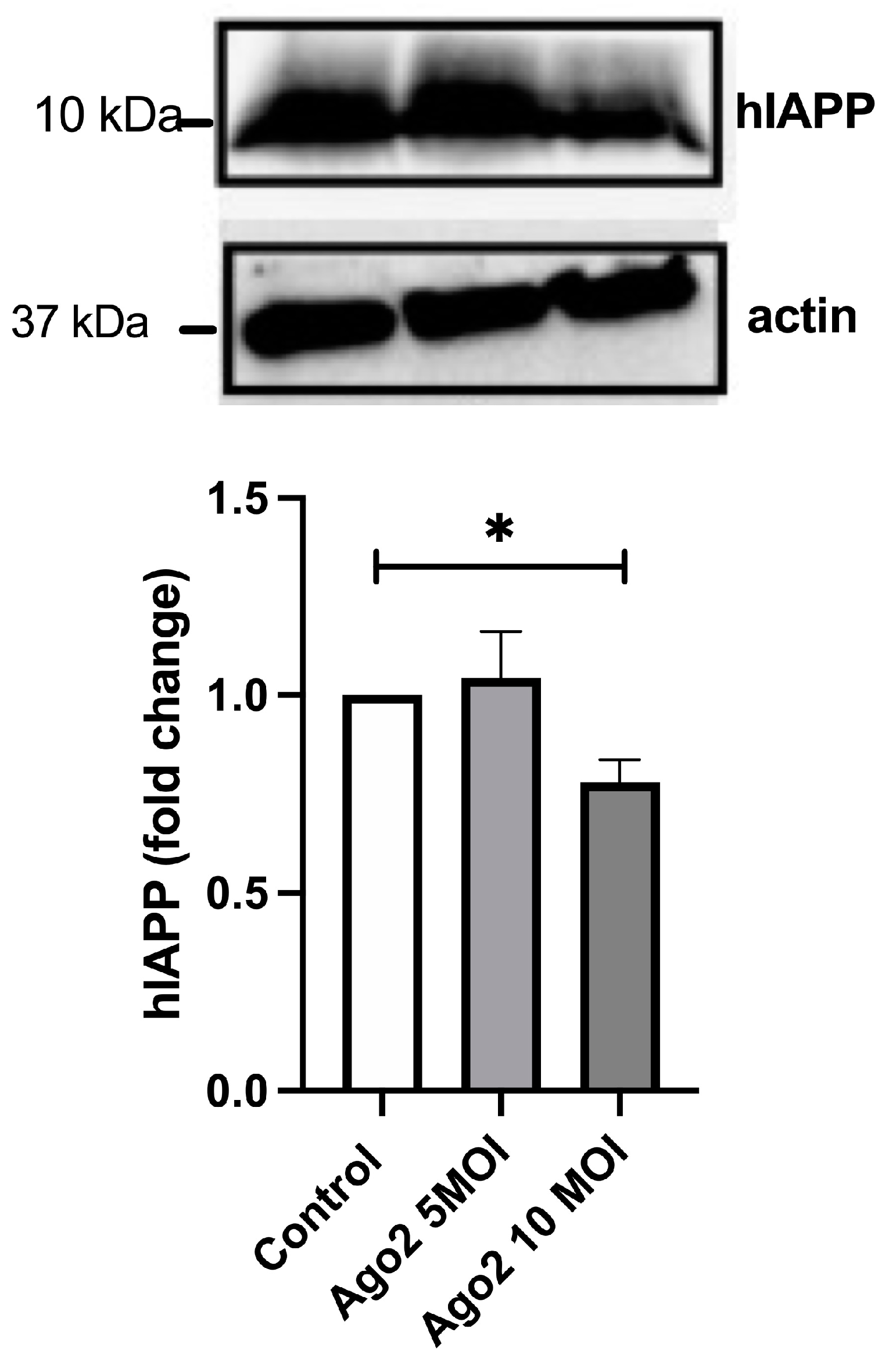
2.6. Catalytic Active Ago-2 Form Limits hIAPP Synthesis in Human and Rat Pancreatic β-Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Transfections
4.3. Translation Activity Assay
4.4. Western Blot Analysis
4.5. Quantitative Real-Time PCR
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Khan, M.A.B.; Hashim, M.J.; King, J.K.; Govender, R.D.; Mustafa, H.; Al Kaabi, J. Epidemiology of Type 2 Diabetes—Global Burden of Disease and Forecasted Trends. J. Epidemiol. Glob. Health 2020, 10, 107–111. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.I.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Primers 2015, 1, 15019. [Google Scholar] [CrossRef]
- Yong, J.; Johnson, J.D.; Arvan, P.; Han, J.; Kaufman, R.J. Therapeutic opportunities for pancreatic beta-cell ER stress in diabetes mellitus. Nat. Rev. Endocrinol. 2021, 17, 455–467. [Google Scholar] [CrossRef] [PubMed]
- Westermark, P.; Andersson, A.; Westermark, G.T. Islet amyloid polypeptide, islet amyloid, and diabetes mellitus. Physiol. Rev. 2011, 91, 795–826. [Google Scholar] [CrossRef]
- Bhowmick, D.C.; Kudaibergenova, Z.; Burnett, L.; Jeremic, A.M. Molecular Mechanisms of Amylin Turnover, Misfolding and Toxicity in the Pancreas. Molecules 2022, 27, 1021. [Google Scholar] [CrossRef]
- Guardado-Mendoza, R.; Davalli, A.M.; Chavez, A.O.; Hubbard, G.B.; Dick, E.J.; Majluf-Cruz, A.; Tene-Perez, C.E.; Goldschmidt, L.; Hart, J.; Perego, C.; et al. Pancreatic islet amyloidosis, beta-cell apoptosis, and alpha-cell proliferation are determinants of islet remodeling in type-2 diabetic baboons. Proc. Natl. Acad. Sci. USA 2009, 106, 13992–13997. [Google Scholar] [CrossRef]
- Akter, R.; Cao, P.; Noor, H.; Ridgway, Z.; Tu, L.H.; Wang, H.; Wong, A.G.; Zhang, X.; Abedini, A.; Schmidt, A.M.; et al. Islet Amyloid Polypeptide: Structure, Function, and Pathophysiology. J. Diabetes Res. 2016, 2016, 2798269. [Google Scholar] [CrossRef] [PubMed]
- Mulder, H.; Ahren, B.; Sundler, F. Islet amyloid polypeptide and insulin gene expression are regulated in parallel by glucose in vivo in rats. Am. J. Physiol. 1996, 271 Pt 1, E1008–E1014. [Google Scholar] [CrossRef] [PubMed]
- Shalev, A.; Pise-Masison, C.A.; Radonovich, M.; Hoffmann, S.C.; Hirshberg, B.; Brady, J.N.; Harlan, D.M. Oligonucleotide microarray analysis of intact human pancreatic islets: Identification of glucose-responsive genes and a highly regulated TGFbeta signaling pathway. Endocrinology 2002, 143, 3695–3698. [Google Scholar] [CrossRef]
- Li, H.; Ha, E.; Donaldson, R.P.; Jeremic, A.M.; Vertes, A. Rapid assessment of human amylin aggregation and its inhibition by copper(II) ions by laser ablation electrospray ionization mass spectrometry with ion mobility separation. Anal. Chem. 2015, 87, 9829–9837. [Google Scholar] [CrossRef]
- Mo, Y.; Lei, J.; Sun, Y.; Zhang, Q.; Wei, G. Conformational Ensemble of hIAPP Dimer: Insight into the Molecular Mechanism by which a Green Tea Extract inhibits hIAPP Aggregation. Sci. Rep. 2016, 6, 33076. [Google Scholar] [CrossRef] [PubMed]
- Rocken, C.; Linke, R.P.; Saeger, W. Immunohistology of islet amyloid polypeptide in diabetes mellitus: Semi-quantitative studies in a post-mortem series. Virchows Arch. A Pathol. Anat. Histopathol. 1992, 421, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Mosselman, S.; Hoppener, J.W.; Lips, C.J.; Jansz, H.S. The complete islet amyloid polypeptide precursor is encoded by two exons. FEBS Lett. 1989, 247, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, W.M.; Campbell, S.C.; Elrick, L.J.; Oates, V.; Bermano, G.; Lindley, K.J.; Aynsley-Green, A.; Dunne, M.J.; James, R.F.; Docherty, K. Glucose regulates islet amyloid polypeptide gene transcription in a PDX1- and calcium-dependent manner. J. Biol. Chem. 2000, 275, 15330–15335. [Google Scholar] [CrossRef]
- Shepherd, L.M.; Campbell, S.C.; Macfarlane, W.M. Transcriptional regulation of the IAPP gene in pancreatic beta-cells. Biochim. Biophys. Acta 2004, 1681, 28–37. [Google Scholar] [CrossRef]
- Watada, H.; Kajimoto, Y.; Kaneto, H.; Matsuoka, T.; Fujitani, Y.; Miyazaki, J.; Yamasaki, Y. Involvement of the homeodomain-containing transcription factor PDX-1 in islet amyloid polypeptide gene transcription. Biochem. Biophys. Res. Commun. 1996, 229, 746–751. [Google Scholar] [CrossRef]
- Le Lay, J.; Matsuoka, T.A.; Henderson, E.; Stein, R. Identification of a novel PDX-1 binding site in the human insulin gene enhancer. J. Biol. Chem. 2004, 279, 22228–22235. [Google Scholar] [CrossRef]
- Gasa, R.; Gomis, R.; Casamitjana, R.; Novials, A. Signals related to glucose metabolism regulate islet amyloid polypeptide (IAPP) gene expression in human pancreatic islets. Regul. Pept. 1997, 68, 99–104. [Google Scholar] [CrossRef]
- Jing, G.; Westwell-Roper, C.; Chen, J.; Xu, G.; Verchere, C.B.; Shalev, A. Thioredoxin-interacting protein promotes islet amyloid polypeptide expression through miR-124a and FoxA2. J. Biol. Chem. 2014, 289, 11807–11815. [Google Scholar] [CrossRef]
- Alarcon, C.; Verchere, C.B.; Rhodes, C.J. Translational control of glucose-induced islet amyloid polypeptide production in pancreatic islets. Endocrinology 2012, 153, 2082–2087. [Google Scholar] [CrossRef]
- Mayya, V.K.; Duchaine, T.F. Ciphers and Executioners: How 3′-Untranslated Regions Determine the Fate of Messenger RNAs. Front. Genet. 2019, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, B.; Seshadri, V.; Fox, P.L. Translational control by the 3′-UTR: The ends specify the means. Trends Biochem. Sci. 2003, 28, 91–98. [Google Scholar] [CrossRef]
- Araujo, P.R.; Yoon, K.; Ko, D.; Smith, A.D.; Qiao, M.; Suresh, U.; Burns, S.C.; Penalva, L.O. Before It Gets Started: Regulating Translation at the 5′ UTR. Comp. Funct. Genom. 2012, 2012, 475731. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee Bhowmick, D.; Burnett, L.; Kudaibergenova, Z.; Jeremic, A.M. FoxA2 and RNA Pol II mediate human islet amyloid polypeptide turnover in ER-stressed pancreatic beta-cells. Biochem. J. 2021, 478, 1261–1282. [Google Scholar] [CrossRef] [PubMed]
- Salunkhe, V.A.; Ofori, J.K.; Gandasi, N.R.; Salo, S.A.; Hansson, S.; Andersson, M.E.; Wendt, A.; Barg, S.; Esguerra, J.L.S.; Eliasson, L. MiR-335 overexpression impairs insulin secretion through defective priming of insulin vesicles. Physiol. Rep. 2017, 5, e13493. [Google Scholar] [CrossRef]
- Tattikota, S.G.; Sury, M.D.; Rathjen, T.; Wessels, H.H.; Pandey, A.K.; You, X.; Becker, C.; Chen, W.; Selbach, M.; Poy, M.N. Argonaute2 regulates the pancreatic beta-cell secretome. Mol. Cell. Proteom. 2013, 12, 1214–1225. [Google Scholar] [CrossRef]
- Matsui, M.; Prakash, T.P.; Corey, D.R. Argonaute 2-dependent Regulation of Gene Expression by Single-stranded miRNA Mimics. Mol. Ther. 2016, 24, 946–955. [Google Scholar] [CrossRef]
- Tattikota, S.G.; Rathjen, T.; McAnulty, S.J.; Wessels, H.H.; Akerman, I.; van de Bunt, M.; Hausser, J.; Esguerra, J.L.; Musahl, A.; Pandey, A.K.; et al. Argonaute2 mediates compensatory expansion of the pancreatic β cell. Cell Metab. 2014, 19, 122–134. [Google Scholar] [CrossRef]
- Kameswaran, V.; Bramswig, N.C.; McKenna, L.B.; Penn, M.; Schug, J.; Hand, N.J.; Chen, Y.; Choi, I.; Vourekas, A.; Won, K.J.; et al. Epigenetic regulation of the DLK1-MEG3 microRNA cluster in human type 2 diabetic islets. Cell Metab. 2014, 19, 135–145. [Google Scholar] [CrossRef]
- Valhmu, W.B.; Palmer, G.D.; Dobson, J.; Fischer, S.G.; Ratcliffe, A. Regulatory activities of the 5′- and 3′-untranslated regions and promoter of the human aggrecan gene. J. Biol. Chem. 1998, 273, 6196–6202. [Google Scholar] [CrossRef]
- Chatterjee Bhowmick, D.; Jeremic, A. Functional proteasome complex is required for turnover of islet amyloid polypeptide in pancreatic beta-cells. J. Biol. Chem. 2018, 293, 14210–14223. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kudaibergenova, Z.; Pany, S.; Placheril, E.; Jeremic, A.M. UTRs and Ago-2/miR-335 Complex Restricts Amylin Translation in Insulinoma and Human Pancreatic β-Cells. Int. J. Mol. Sci. 2024, 25, 9614. https://doi.org/10.3390/ijms25179614
Kudaibergenova Z, Pany S, Placheril E, Jeremic AM. UTRs and Ago-2/miR-335 Complex Restricts Amylin Translation in Insulinoma and Human Pancreatic β-Cells. International Journal of Molecular Sciences. 2024; 25(17):9614. https://doi.org/10.3390/ijms25179614
Chicago/Turabian StyleKudaibergenova, Zhanar, Satyabrata Pany, Elizabeth Placheril, and Aleksandar M. Jeremic. 2024. "UTRs and Ago-2/miR-335 Complex Restricts Amylin Translation in Insulinoma and Human Pancreatic β-Cells" International Journal of Molecular Sciences 25, no. 17: 9614. https://doi.org/10.3390/ijms25179614
APA StyleKudaibergenova, Z., Pany, S., Placheril, E., & Jeremic, A. M. (2024). UTRs and Ago-2/miR-335 Complex Restricts Amylin Translation in Insulinoma and Human Pancreatic β-Cells. International Journal of Molecular Sciences, 25(17), 9614. https://doi.org/10.3390/ijms25179614