
Cardiotoxicity of Iron and Zinc and Their Association with the Mitochondrial Unfolded Protein Response in Humans
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Concentrations of Essential Trace Elements in Healthy and Heart-Failure Myocardium
2.2. Cumulative Concentration Index of Iron and Zinc and the Patient’s Age of Receiving Heart Transplantation or LVAD
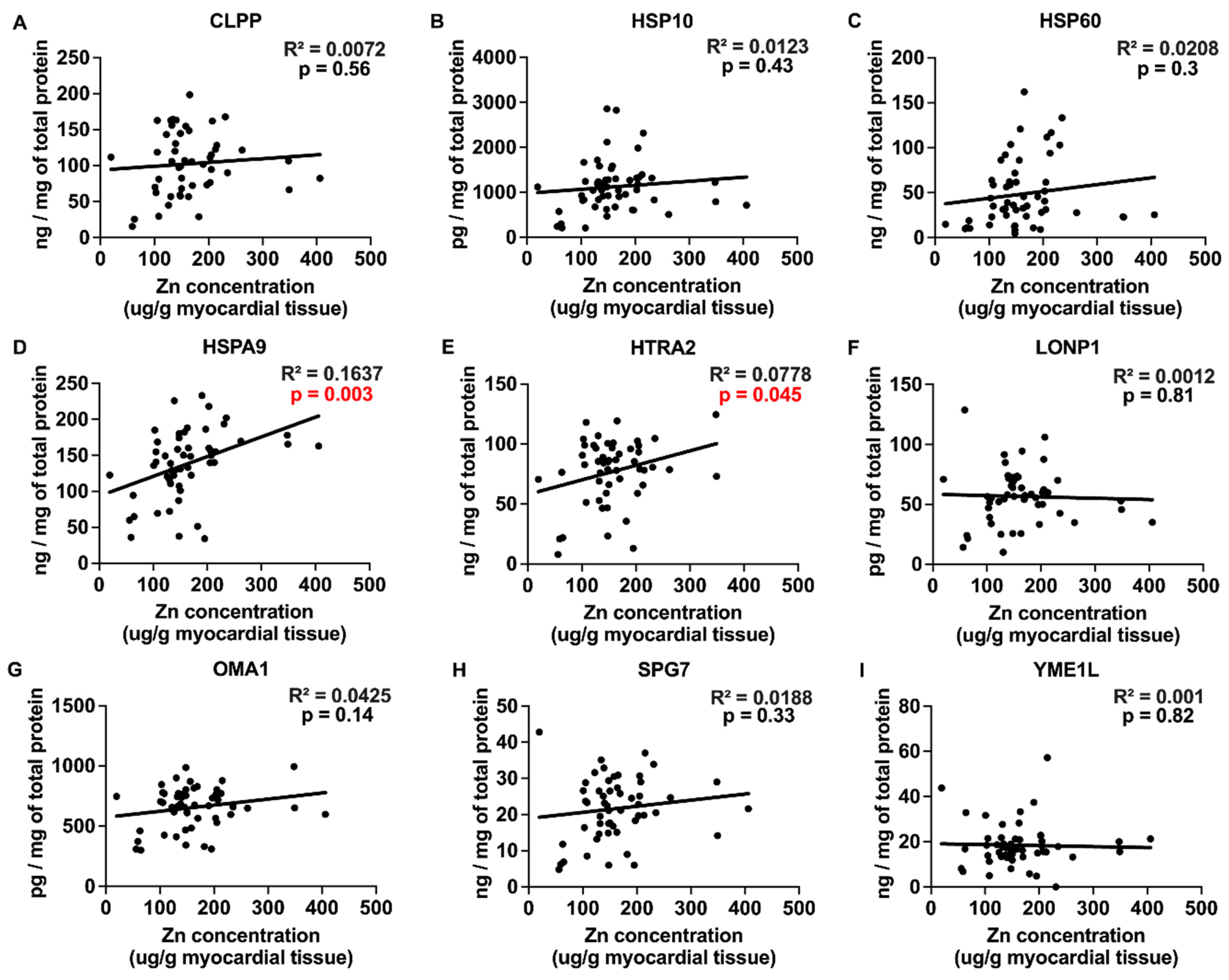
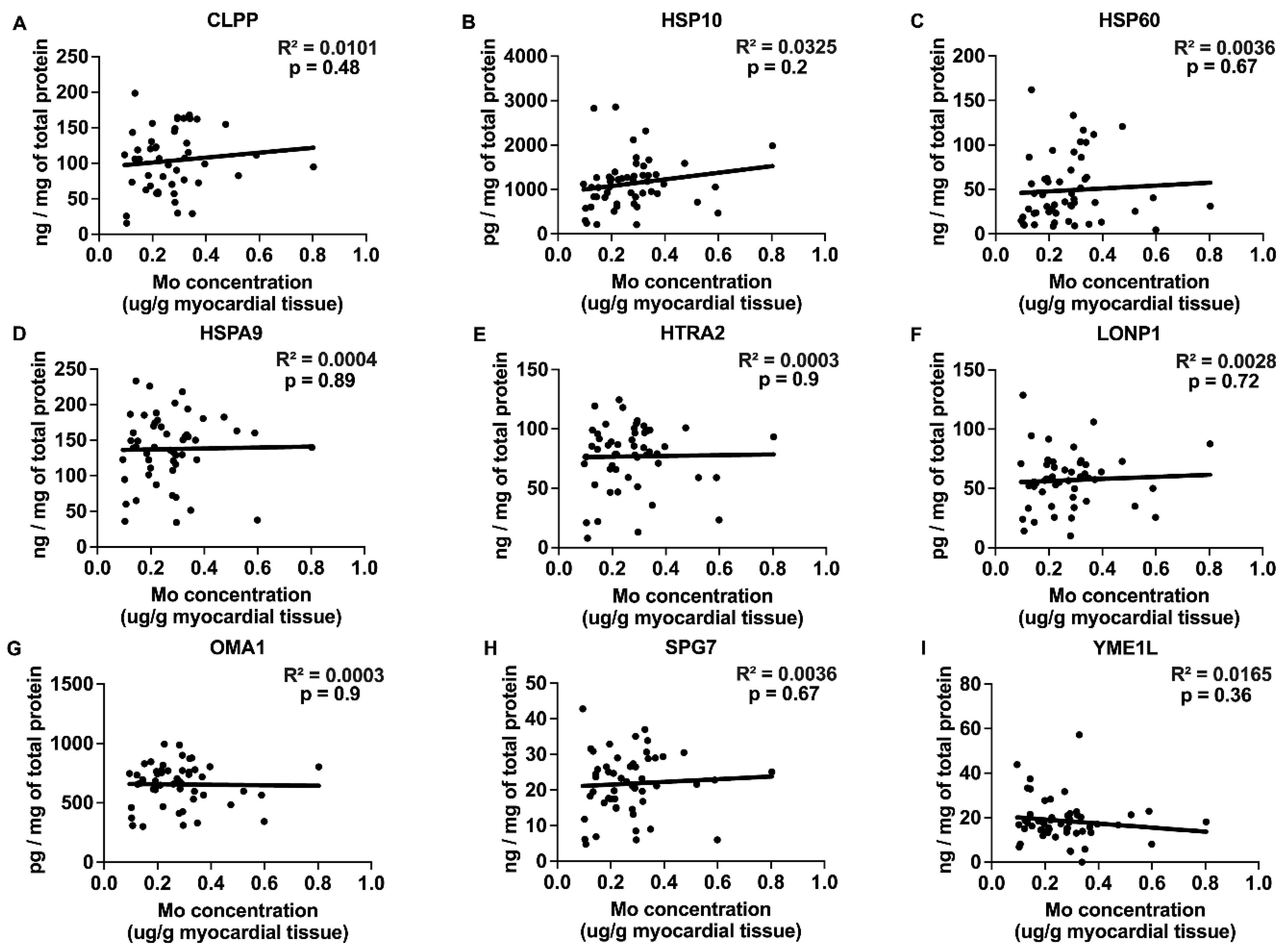
2.3. Correlations between Myocardial Trace Elements and the Expression of UPRmt Proteins
2.4. Trace Elements and Oxidative Stress by 4-HNE Detection in the Myocardium of Failing Hearts
2.5. Trace Element Concentrations in ICM and DCM
3. Discussion
4. Materials and Methods
4.1. Human Heart Samples
4.2. Analysis of Trace Element Concentrations in Human Myocardium
4.3. Calculation of the Cumulative Concentration Index of Trace Elements
4.4. Quantification of Myocardial UPRmt Protein Expression and 4-HNE by ELISA
4.5. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pu, W.; Chu, X.; Xu, S.; Dai, X.; Xiao, L.; Cui, T.; Huang, B.; Hu, G.; Zhang, C. Molybdenum Exposure Induces Inflammatory Response via the Regulatory Effects of lncRNA-00072124/miR-308/OSMR Crosstalk on JAK/STAT Axis in Duck Kidneys. Sci. Total Environ. 2024, 912, 169374. [Google Scholar] [CrossRef] [PubMed]
- Joshi, P.K.; Patel, S.C.; Shreya, D.; Zamora, D.I.; Patel, G.S.; Grossmann, I.; Rodriguez, K.; Soni, M.; Sange, I. Hereditary Hemochromatosis: A Cardiac Perspective. Cureus 2021, 13, e20009. [Google Scholar] [CrossRef] [PubMed]
- Ćirović, A.; Buha Đorđević, A.; Ćirović, A.; Jevtić, J.; Tasić, D.; Janković, S.; Antonijević, B.; Petrović, Z.; Orisakwe, O.E.; Tasić, N. Trace Element Concentrations in Autopsied Heart Tissues from Patients with Secondary Cardiomyopathy. Biol. Trace Elem. Res. 2024, 202, 2442–2449. [Google Scholar] [CrossRef]
- Pan, M.; Cheng, Z.-W.; Huang, C.-G.; Ye, Z.-Q.; Sun, L.-J.; Chen, H.; Fu, B.-B.; Zhou, K.; Fang, Z.-R.; Wang, Z.-J.; et al. Long-Term Exposure to Copper Induces Mitochondria-Mediated Apoptosis in Mouse Hearts. Ecotoxicol. Environ. Saf. 2022, 234, 113329. [Google Scholar] [CrossRef]
- Lipiński, P.; Starzyński, R.R.; Styś, A.; Straciło, M. Iron homeostasis, a defense mechanism in oxidative stress. Postepy Biochem. 2010, 56, 305–316. [Google Scholar] [PubMed]
- Touati, D. Iron and Oxidative Stress in Bacteria. Arch. Biochem. Biophys. 2000, 373, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kerins, M.J.; Ooi, A. The Roles of NRF2 in Modulating Cellular Iron Homeostasis. Antioxid. Redox Signal. 2018, 29, 1756–1773. [Google Scholar] [CrossRef]
- Chen, P.; Bornhorst, J.; Aschner, M. Manganese Metabolism in Humans. Front. Biosci. Landmark Ed. 2018, 23, 1655–1679. [Google Scholar] [CrossRef]
- Franklin, R.; Hare, M.; Beckman, J.S. Determining Copper and Zinc Content in Superoxide Dismutase Using Electron Capture Dissociation under Native Spray Conditions. Methods Mol. Biol. 2022, 2500, 201–210. [Google Scholar] [CrossRef]
- Horn, D.; Barrientos, A. Mitochondrial Copper Metabolism and Delivery to Cytochrome c Oxidase. IUBMB Life 2008, 60, 421–429. [Google Scholar] [CrossRef]
- Müller, M.; Donhauser, E.; Maske, T.; Bischof, C.; Dumitrescu, D.; Rudolph, V.; Klinke, A. Mitochondrial Integrity Is Critical in Right Heart Failure Development. Int. J. Mol. Sci. 2023, 24, 11108. [Google Scholar] [CrossRef]
- Bisaccia, G.; Ricci, F.; Gallina, S.; Di Baldassarre, A.; Ghinassi, B. Mitochondrial Dysfunction and Heart Disease: Critical Appraisal of an Overlooked Association. Int. J. Mol. Sci. 2021, 22, 614. [Google Scholar] [CrossRef] [PubMed]
- Pravdic, D.; Sedlic, F.; Mio, Y.; Vladic, N.; Bienengraeber, M.; Bosnjak, Z.J. Anesthetic-Induced Preconditioning Delays Opening of Mitochondrial Permeability Transition Pore via Protein Kinase C-Epsilon-Mediated Pathway. Anesthesiology 2009, 111, 267–274. [Google Scholar] [CrossRef]
- Muravyeva, M.; Baotic, I.; Bienengraeber, M.; Lazar, J.; Bosnjak, Z.J.; Sedlic, F.; Warltier, D.C.; Kersten, J.R. Cardioprotection during Diabetes: The Role of Mitochondrial DNA. Anesthesiology 2014, 120, 870–879. [Google Scholar] [CrossRef]
- Sepac, A.; Sedlic, F.; Si-Tayeb, K.; Lough, J.; Duncan, S.A.; Bienengraeber, M.; Park, F.; Kim, J.; Bosnjak, Z.J. Isoflurane Preconditioning Elicits Competent Endogenous Mechanisms of Protection from Oxidative Stress in Cardiomyocytes Derived from Human Embryonic Stem Cells. Anesthesiology 2010, 113, 906–916. [Google Scholar] [CrossRef] [PubMed]
- Svaguša, T.; Martinić, M.; Martinić, M.; Kovačević, L.; Šepac, A.; Miličić, D.; Bulum, J.; Starčević, B.; Sirotković-Skerlev, M.; Seiwerth, F.; et al. Mitochondrial Unfolded Protein Response, Mitophagy and Other Mitochondrial Quality Control Mechanisms in Heart Disease and Aged Heart. Croat. Med. J. 2020, 61, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Wei, Y.; Song, Q.; Du, B.; Wang, H.; Chu, Y.; Hu, Y. The Role of Myocardial Mitochondrial Quality Control in Heart Failure. Front. Pharmacol. 2019, 10, 1404. [Google Scholar] [CrossRef]
- Svagusa, T.; Sikiric, S.; Milavic, M.; Sepac, A.; Seiwerth, S.; Milicic, D.; Gasparovic, H.; Biocina, B.; Rudez, I.; Sutlic, Z.; et al. Heart Failure in Patients Is Associated with Downregulation of Mitochondrial Quality Control Genes. Eur. J. Clin. Investig. 2023, 53, e14054. [Google Scholar] [CrossRef]
- Rizzo, C.; Carbonara, R.; Ruggieri, R.; Passantino, A.; Scrutinio, D. Iron Deficiency: A New Target for Patients with Heart Failure. Front. Cardiovasc. Med. 2021, 8, 709872. [Google Scholar] [CrossRef]
- Sica, D.A. Pharmacotherapy in Congestive Heart Failure: Drug Absorption in the Management of Congestive Heart Failure: Loop Diuretics. Congest. Heart Fail. 2003, 9, 287–292. [Google Scholar] [CrossRef]
- Zischka, H.; Borchard, S. Chapter 8—Mitochondrial Copper Toxicity with a Focus on Wilson Disease. In Clinical and Translational Perspectives on WILSON DISEASE; Kerkar, N., Roberts, E.A., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 65–75. ISBN 978-0-12-810532-0. [Google Scholar]
- Sumneang, N.; Siri-Angkul, N.; Kumfu, S.; Chattipakorn, S.C.; Chattipakorn, N. The Effects of Iron Overload on Mitochondrial Function, Mitochondrial Dynamics, and Ferroptosis in Cardiomyocytes. Arch. Biochem. Biophys. 2020, 680, 108241. [Google Scholar] [CrossRef] [PubMed]
- Schoofs, H.; Schmit, J.; Rink, L. Zinc Toxicity: Understanding the Limits. Molecular 2024, 29, 3130. [Google Scholar] [CrossRef] [PubMed]
- Tarnacka, B.; Jopowicz, A.; Maślińska, M. Copper, Iron, and Manganese Toxicity in Neuropsychiatric Conditions. Int. J. Mol. Sci. 2021, 22, 7820. [Google Scholar] [CrossRef] [PubMed]
- Wai, T.; García-Prieto, J.; Baker, M.J.; Merkwirth, C.; Benit, P.; Rustin, P.; Rupérez, F.J.; Barbas, C.; Ibañez, B.; Langer, T. Imbalanced OPA1 Processing and Mitochondrial Fragmentation Cause Heart Failure in Mice. Science 2015, 350, aad0116. [Google Scholar] [CrossRef]
- Venkatesh, S.; Li, M.; Saito, T.; Tong, M.; Rashed, E.; Mareedu, S.; Zhai, P.; Bárcena, C.; López-Otín, C.; Yehia, G.; et al. Mitochondrial LonP1 Protects Cardiomyocytes from Ischemia/Reperfusion Injury In Vivo. J. Mol. Cell. Cardiol. 2019, 128, 38–50. [Google Scholar] [CrossRef]
- Seiferling, D.; Szczepanowska, K.; Becker, C.; Senft, K.; Hermans, S.; Maiti, P.; König, T.; Kukat, A.; Trifunovic, A. Loss of CLPP Alleviates Mitochondrial Cardiomyopathy without Affecting the Mammalian UPRmt. EMBO Rep. 2016, 17, 953–964. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, L.; Tso, A.; Wang, S.; Fang, X.; Ouyang, K.; Han, Z. Mitochondrial Chaperones and Proteases in Cardiomyocytes and Heart Failure. Front. Mol. Biosci. 2021, 8, 630332. [Google Scholar] [CrossRef]
- Smyrnias, I.; Gray, S.P.; Okonko, D.O.; Sawyer, G.; Zoccarato, A.; Catibog, N.; López, B.; González, A.; Ravassa, S.; Díez, J.; et al. Cardioprotective Effect of the Mitochondrial Unfolded Protein Response During Chronic Pressure Overload. J. Am. Coll. Cardiol. 2019, 73, 1795–1806. [Google Scholar] [CrossRef]
- Hu, X.; Van Marion, D.M.S.; Wiersma, M.; Zhang, D.; Brundel, B.J.J.M. The Protective Role of Small Heat Shock Proteins in Cardiac Diseases: Key Role in Atrial Fibrillation. Cell Stress Chaperones 2017, 22, 665–674. [Google Scholar] [CrossRef]
- Endale, H.T.; Tesfaye, W.; Mengstie, T.A. ROS Induced Lipid Peroxidation and Their Role in Ferroptosis. Front. Cell Dev. Biol. 2023, 11, 1226044. [Google Scholar] [CrossRef]
- Li, L.; Yang, X. The Essential Element Manganese, Oxidative Stress, and Metabolic Diseases: Links and Interactions. Oxid. Med. Cell. Longev. 2018, 2018, 7580707. [Google Scholar] [CrossRef]
- Choi, S.; Liu, X.; Pan, Z. Zinc Deficiency and Cellular Oxidative Stress: Prognostic Implications in Cardiovascular Diseases. Acta Pharmacol. Sin. 2018, 39, 1120–1132. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Li, K.; Xing, W.; Dong, M.; Yi, M.; Zhang, H. Role of Iron-Related Oxidative Stress and Mitochondrial Dysfunction in Cardiovascular Diseases. Oxid. Med. Cell. Longev. 2022, 2022, 5124553. [Google Scholar] [CrossRef]
- Marreiro, D.d.N.; Cruz, K.J.C.; Morais, J.B.S.; Beserra, J.B.; Severo, J.S.; de Oliveira, A.R.S. Zinc and Oxidative Stress: Current Mechanisms. Antioxidants 2017, 6, 24. [Google Scholar] [CrossRef]
- Rosenblum, H.; Wessler, J.D.; Gupta, A.; Maurer, M.S.; Bikdeli, B. Zinc Deficiency and Heart Failure: A Systematic Review of the Current Literature. J. Card. Fail. 2020, 26, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Suliburska, J.; Skrypnik, K.; Szulińska, M.; Kupsz, J.; Markuszewski, L.; Bogdański, P. Diuretics, Ca-Antagonists, and Angiotensin-Converting Enzyme Inhibitors Affect Zinc Status in Hypertensive Patients on Monotherapy: A Randomized Trial. Nutrients 2018, 10, 1284. [Google Scholar] [CrossRef] [PubMed]
- Kamalov, G.; Bhattacharya, S.K.; Ahokas, R.A.; Sun, Y.; Weber, K.T.; Gerling, I.C.; Holewinski, J.P. Nutrient Dyshomeostasis in Congestive Heart Failure. Am. J. Med. Sci. 2009, 338, 28–33. [Google Scholar] [CrossRef]
- Ripa, S.; Ripa, R. Zinc and arterial pressure. Minerva Med. 1994, 85, 455–459. [Google Scholar]
- Ahmed, S.; Peterson, S.J.; Parikh, M.A.; Frishman, W.H. Cardiovascular Manifestations of Hemochromatosis: A Review of Pathophysiology, Mechanisms, and Treatment Options. Cardiol. Rev. 2023. [Google Scholar] [CrossRef]
- Liu, Z.-Y.; Liu, Z.-Y.; Lin, L.-C.; Song, K.; Tu, B.; Zhang, Y.; Yang, J.-J.; Zhao, J.-Y.; Tao, H. Redox Homeostasis in Cardiac Fibrosis: Focus on Metal Ion Metabolism. Redox Biol. 2024, 71, 103109. [Google Scholar] [CrossRef]
- Harding, D.; Chong, M.H.A.; Lahoti, N.; Bigogno, C.M.; Prema, R.; Mohiddin, S.A.; Marelli-Berg, F. Dilated Cardiomyopathy and Chronic Cardiac Inflammation: Pathogenesis, Diagnosis and Therapy. J. Intern. Med. 2023, 293, 23–47. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Xu, Z.; Chen, L.; Ye, D.; Yu, Y.; Zhang, Y.; Cao, Y.; Djibril, B.; Guo, X.; Gao, X.; et al. Iron Overload Inhibits Self-Renewal of Human Pluripotent Stem Cells via DNA Damage and Generation of Reactive Oxygen Species. FEBS Open Bio 2020, 10, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Chen, Y.; Luz, A.; Hu, G.; Tokar, E.J. Cardiac Development in the Presence of Cadmium: An In Vitro Study Using Human Embryonic Stem Cells and Cardiac Organoids. Environ. Health Perspect. 2022, 130, 117002. [Google Scholar] [CrossRef] [PubMed]
- Sepac, A.; Si-Tayeb, K.; Sedlic, F.; Barrett, S.; Canfield, S.; Duncan, S.A.; Bosnjak, Z.J.; Lough, J.W. Comparison of Cardiomyogenic Potential among Human ESC and iPSC Lines. Cell Transplant. 2012, 21, 2523–2530. [Google Scholar] [CrossRef]
- Sepac, A.; Bosnjak, J.Z.; Seiwerth, S.; Sikiric, S.; Regovic Dzombeta, T.; Kulic, A.; Karsaj, J.M.; Sedlic, F. Human C2a and C6a iPSC Lines and H9 ESC Line Have Less Efficient Cardiomyogenesis than H1 ESC Line: Beating Enhances Cardiac Differentiation. Int. J. Dev. Biol. 2021, 65, 523–536. [Google Scholar] [CrossRef]
- Wang, M.; Tian, Y.; Yu, P.; Li, N.; Deng, Y.; Li, L.; Kang, H.; Chen, D.; Wang, H.; Liu, Z.; et al. Association between Congenital Heart Defects and Maternal Manganese and Iron Concentrations: A Case-Control Study in China. Environ. Sci. Pollut. Res. Int. 2022, 29, 26950–26959. [Google Scholar] [CrossRef]
- Piavchenko, G.; Alekseev, A.; Stelmashchuk, O.; Seryogina, E.; Zherebtsov, E.; Kuznetsova, E.; Dunaev, A.; Volkov, Y.; Kuznetsov, S. A Complex Morphofunctional Approach for Zinc Toxicity Evaluation in Rats. Heliyon 2020, 6, e03768. [Google Scholar] [CrossRef]
- Svaguša, T.; Sedlić, F.; Županić, S.; Manola, Š.; Bakoš, M.; Mirošević, V.; Livun, A. A Rare Form of LIM Domain-Binding Protein 3 (LDB3) Mutation Causes Hypertrophic Cardiomyopathy and Myofibrillar Myopathy Type 4. Clin. Genet. 2024. [Google Scholar] [CrossRef]
- Quiroz, C.J.C.; Choque, G.J.M.; Mamani, M.C.; Quispe, G.D.L.F. Evaluation of the Content of Metals and Contamination Indices Generated by Environmental Liabilities, in Tacna, Peru. Contamination 2022. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Control Group | Heart Failure Group | p |
|---|---|---|---|
| CLPP | 174.5 ± 24.7 ng/mg | 106.4 ± 6.3 ng/mg | 0.0015 |
| HSPA9 | 163.3 ± 17.8 ng/mg | 135.4 ± 6.3 ng/mg | 0.1786 |
| SPG7 | 35.2 ± 7.9 ng/mg | 27.6 ± 1.2 ng/mg | 0.0028 |
| OMA1 | 947.2 ± 61.5 pg/mg | 665.7 ± 24.2 pg/mg | 0.0004 |
| LONP1 | 109.5 ± 21.6 pg/mg | 58.0 ± 3.3 pg/mg | <0.0001 |
| HSP10 | 2590 ± 312 pg/mg | 1075 ± 66 pg/mg | <0.0001 |
| HSP60 | 139 ± 55 ng/mg | 46 ± 4 ng/mg | <0.0001 |
| HTRA2 | 77.5 ± 8.7 ng/mg | 74.5 ± 3.6 ng/mg | 0.0891 |
| YME1L | 20.9 ± 4.3 ng/mg | 17.7 ± 1.1 ng/mg | 0.4171 |
| Dependent Variables | ||||||||||
| Independent Variable | CLPP | HSP10 | HSP60 | HSPA9 | ||||||
| t | p | t | p | t | p | t | p | |||
| Cu | 1.25 | 0.22 | −0.08 | 0.93 | 0.74 | 0.47 | 0.59 | 0.56 | ||
| Fe | 0.07 | 0.94 | 0.777 | 0.44 | 0.18 | 0.86 | −1.86 | 0.07 | ||
| Mn | 0.53 | 0.6 | 2.24 | 0.03 | 1.738 | 0.09 | 1.893 | 0.06 | ||
| Mo | −0.44 | 0.65 | −0.19 | 0.85 | −1.12 | 0.25 | −0.84 | 0.41 | ||
| Zn | −0.11 | 0.91 | 0.62 | 0.54 | 0.88 | 0.38 | 3.11 | 0.003 | ||
| Age of Htx/LVAD | 2.81 | 0.008 | 0.92 | 0.36 | 0.4 | 0.69 | 2.27 | 0.03 | ||
| Sex of patients | 2.40 | 0.02 | 1.47 | 0.15 | 3.50 | 0.001 | 0.68 | 0.50 | ||
| Dependent Variables | ||||||||||
| Independent Variable | HTRA2 | LONP1 | OMA1 | SPG7 | YME1L | |||||
| t | p | t | p | t | p | t | p | t | p | |
| Cu | 0.22 | 0.83 | 0.22 | 0.83 | −0.69 | 0.50 | 0.24 | 0.81 | 0.40 | 0.69 |
| Fe | −0.36 | 0.72 | 0.14 | 0.89 | −0.39 | 0.70 | −1.31 | 0.20 | −2.60 | 0.01 |
| Mn | 2.16 | 0.04 | 0.51 | 0.61 | 0.778 | 0.44 | 0.89 | 0.38 | 0.54 | 0.60 |
| Mo | −0.87 | 0.39 | 0.27 | 0.79 | −0.49 | 0.63 | −0.35 | 0.73 | −0.12 | 0.91 |
| Zn | 1.44 | 0.16 | 0.23 | 0.82 | 1.978 | 0.049 | 1.20 | 0.24 | 0.10 | 0.92 |
| Age of Htx/LVAD | 1.23 | 0.22 | 1.43 | 0.16 | 1.642 | 0.11 | 2.31 | 0.03 | 0.59 | 0.56 |
| Gender | 1.56 | 0.13 | 2.08 | 0.04 | 0.04 | 0.96 | 1.43 | 0.16 | −1.23 | 0.23 |
| Number of patients | 128 |
| Male sex, n (%) | 104 (81%) |
| Age (y), means ± SEM | 57 ± 1 |
| BMI (kg/m2), means ± SEM | 27.0 ± 0.3 |
| Smoking, n (%) | 59 (46) |
| AH, n (%) | 81 (63) |
| DM, n (%) | 44 (34) |
| Dyslipidemia, n (%) | 76 (59) |
| AF, n (%) | 47 (36) |
| IHD, n (%) | 63 (49) |
| CKD, n (%) | 39 (30) |
| TGD, n (%) | 20 (15) |
| EF (%) means ± SEM | 25 ± 1 |
| LVIDd (cm), means ± SEM | 7.1 ± 0.1 |
| LVIDs (cm), means ± SEM | 6.3 ± 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirosevic, V.; Svagusa, T.; Matic, N.; Maldini, K.; Siljeg, M.; Milicic, D.; Gasparovic, H.; Rudez, I.; Sepac, A.; Gojmerac, L.; et al. Cardiotoxicity of Iron and Zinc and Their Association with the Mitochondrial Unfolded Protein Response in Humans. Int. J. Mol. Sci. 2024, 25, 9648. https://doi.org/10.3390/ijms25179648
Mirosevic V, Svagusa T, Matic N, Maldini K, Siljeg M, Milicic D, Gasparovic H, Rudez I, Sepac A, Gojmerac L, et al. Cardiotoxicity of Iron and Zinc and Their Association with the Mitochondrial Unfolded Protein Response in Humans. International Journal of Molecular Sciences. 2024; 25(17):9648. https://doi.org/10.3390/ijms25179648
Chicago/Turabian StyleMirosevic, Vid, Tomo Svagusa, Natalija Matic, Kresimir Maldini, Mario Siljeg, Davor Milicic, Hrvoje Gasparovic, Igor Rudez, Ana Sepac, Lucija Gojmerac, and et al. 2024. "Cardiotoxicity of Iron and Zinc and Their Association with the Mitochondrial Unfolded Protein Response in Humans" International Journal of Molecular Sciences 25, no. 17: 9648. https://doi.org/10.3390/ijms25179648