
MicroRNA-27a-5p Downregulates Expression of Proinflammatory Cytokines in Lipopolysaccharide-Stimulated Human Dental Pulp Cells via the NF-κB Signaling Pathway

 , , , ,
, , , ,
Abstract
1. Introduction
2. Results
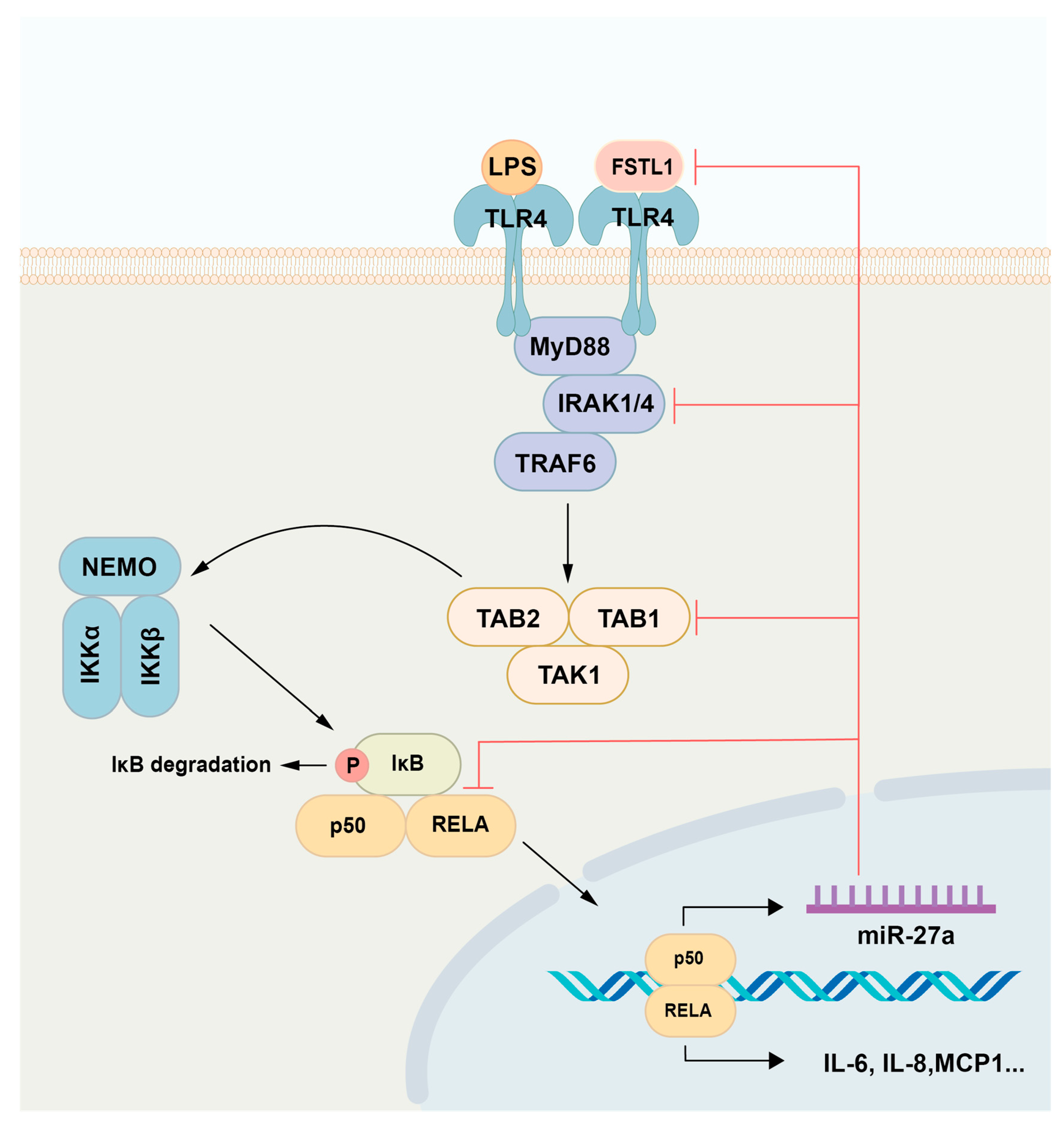
2.1. LPS/NF-κB Signaling Promotes miR-27a-5p Expression and Proinflammatory Mediator Levels in LPS-Stimulated hDPCs
2.2. MiR-27a-5p Downregulates Synthesis of Proinflammatory Mediators via the NF-κB Pathway in LPS-Stimulated hDPCs
2.3. MiR-27a-5p Targets TAB1, IRAK4, and RELA in NF-κB Signaling
2.4. MiR-27a-5p Exerts a Downregulatory Effect on NF-κB Activity and Proinflammatory Cytokine Expression in LPS-Stimulated Rat Pulp Tissue Ex Vivo
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatments
4.2. Establishment of the Pulpitis Model in Rats
4.3. Ex Vivo miR-27a Mimic Transfection
4.4. Reverse Transcription-Quantitative Polymerase Chain Reaction
4.5. Western Blotting
4.6. Cytometric Bead Array
4.7. Luciferase Assays Using NF-κB and TAB1 3′-UTR Reporter Vectors
4.8. Immunofluorescence
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hahn, C.-L.; Liewehr, F.R. Relationships between Caries Bacteria, Host Responses, and Clinical Signs and Symptoms of Pulpitis. J. Endod. 2007, 33, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Jontell, M.; Okiji, T.; Dahlgren, U.; Bergenholtz, G. Immune Defense Mechanisms of the Dental Pulp. Crit. Rev. Oral Biol. Med. Off. Publ. Am. Assoc. Oral Biol. 1998, 9, 179–200. [Google Scholar] [CrossRef]
- Jiang, H.-W.; Zhang, W.; Ren, B.-P.; Zeng, J.-F.; Ling, J.-Q. Expression of Toll like Receptor 4 in Normal Human Odontoblasts and Dental Pulp Tissue. J. Endod. 2006, 32, 747–751. [Google Scholar] [CrossRef]
- Mutoh, N.; Tani-Ishii, N.; Tsukinoki, K.; Chieda, K.; Watanabe, K. Expression of Toll-like Receptor 2 and 4 in Dental Pulp. J. Endod. 2007, 33, 1183–1186. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef]
- Kokkas, A.B.; Goulas, A.; Varsamidis, K.; Mirtsou, V.; Tziafas, D. Irreversible but Not Reversible Pulpitis Is Associated with Up-Regulation of Tumour Necrosis Factor-Alpha Gene Expression in Human Pulp. Int. Endod. J. 2007, 40, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.R.; Takahashi, Y.; Graham, L.W.; Simon, S.; Imazato, S.; Smith, A.J. Inflammation–Regeneration Interplay in the Dentine–Pulp Complex. J. Dent. 2010, 38, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Hahn, C.-L.; Liewehr, F.R. Innate Immune Responses of the Dental Pulp to Caries. J. Endod. 2007, 33, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Giraud, T.; Jeanneau, C.; Rombouts, C.; Bakhtiar, H.; Laurent, P.; About, I. Pulp Capping Materials Modulate the Balance between Inflammation and Regeneration. Dent. Mater. 2019, 35, 24–35. [Google Scholar] [CrossRef]
- Lara, V.S.; Figueiredo, F.; da Silva, T.A.; Cunha, F.Q. Dentin-Induced in Vivo Inflammatory Response and in Vitro Activation of Murine Macrophages. J. Dent. Res. 2003, 82, 460–465. [Google Scholar] [CrossRef]
- Yamada, M.; Fujino, N.; Ichinose, M. Inflammatory Responses in the Initiation of Lung Repair and Regeneration: Their Role in Stimulating Lung Resident Stem Cells. Inflamm. Regen. 2016, 36, 15. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.R.; Holder, M.J.; Smith, A.J. Inflammation and Regeneration in the Dentin-Pulp Complex: A Double-Edged Sword. J. Endod. 2014, 40 (Suppl. S4), S46–S51. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.; Njeh, A.; Uzunoglu, E. Is Pulp Inflammation a Prerequisite for Pulp Healing and Regeneration? Mediat. Inflamm. 2015, 2015, e347649. [Google Scholar] [CrossRef]
- Lai, E.C. Micro RNAs Are Complementary to 3′ UTR Sequence Motifs That Mediate Negative Post-Transcriptional Regulation. Nat. Genet. 2002, 30, 363–364. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Mens, M.M.J.; Ghanbari, M. Cell Cycle Regulation of Stem Cells by MicroRNAs. Stem Cell Rev. 2018, 14, 309–322. [Google Scholar] [CrossRef]
- Hui, T.; Wang, C.; Chen, D.; Zheng, L.; Huang, D.; Ye, L. Epigenetic Regulation in Dental Pulp Inflammation. Oral Dis. 2017, 23, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Carrillo, J.L.; Vázquez-Alcaraz, S.J.; Vargas-Barbosa, J.M.; Ramos-Gracia, L.G.; Alvarez-Barreto, I.; Medina-Quiroz, A.; Díaz-Huerta, K.K. 2. The Role of MicroRNAs in Pulp Inflammation. Cells 2021, 10, 2142. [Google Scholar] [CrossRef]
- Taganov, K.D.; Boldin, M.P.; Chang, K.-J.; Baltimore, D. NF-ΚB-Dependent Induction of MicroRNA MiR-146, an Inhibitor Targeted to Signaling Proteins of Innate Immune Responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef]
- Wang, D.; Sun, S.; Xue, Y.; Qiu, J.; Ye, T.; Zhang, R.; Song, B.; He, W.; Zhang, Y.; Jiang, W. MicroRNA-223 Negatively Regulates LPS-Induced Inflammatory Responses by Targeting NLRP3 in Human Dental Pulp Fibroblasts. Int. Endod. J. 2021, 54, 241–254. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Tian, J.; Liu, G.; Li, X.; Shen, D. Sevoflurane Alleviates LPS-induced Acute Lung Injury via the MicroRNA-27a-3p/TLR4/MyD88/NF-κB Signaling Pathway. Int. J. Mol. Med. 2019, 44, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Q.; Lv, H.-W.; Wang, Z.-L.; Tan, W.-F.; Fang, B.; Ma, H. MiR-27a Ameliorates Inflammatory Damage to the Blood-Spinal Cord Barrier after Spinal Cord Ischemia: Reperfusion Injury in Rats by Downregulating TICAM-2 of the TLR4 Signaling Pathway. J. Neuroinflamm. 2015, 12, 25. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.-J.; Xu, M.-Z.; Zhu, X.-D.; Ji, Y.-H. MicroRNA-27a Alleviates IL-1β-Induced Inflammatory Response and Articular Cartilage Degradation via TLR4/NF-ΚB Signaling Pathway in Articular Chondrocytes. Int. Immunopharmacol. 2019, 76, 105839. [Google Scholar] [CrossRef]
- Romay, M.C.; Che, N.; Becker, S.N.; Pouldar, D.; Hagopian, R.; Xiao, X.; Lusis, A.J.; Berliner, J.A.; Civelek, M. Regulation of NF-ΚB Signaling by Oxidized Glycerophospholipid and IL-1β Induced MiRs-21-3p and -27a-5p in Human Aortic Endothelial Cells [S]. J. Lipid Res. 2015, 56, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Lau, M.C.-C.; Ng, K.Y.; Wong, T.L.; Tong, M.; Lee, T.K.; Ming, X.-Y.; Law, S.; Lee, N.P.; Cheung, A.L.; Qin, Y.-R.; et al. FSTL1 Promotes Metastasis and Chemoresistance in Esophageal Squamous Cell Carcinoma through NFκB–BMP Signaling Cross-Talk. Cancer Res. 2017, 77, 5886–5899. [Google Scholar] [CrossRef]
- Lv, Y.-N.; Ou-Yang, A.-J.; Fu, L.-S. MicroRNA-27a Negatively Modulates the Inflammatory Response in Lipopolysaccharide-Stimulated Microglia by Targeting TLR4 and IRAK4. Cell. Mol. Neurobiol. 2017, 37, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Shi, G.; Xie, J.; Du, X.; Yang, H. MicroRNA-27a Inhibits Cell Migration and Invasion of Fibroblast-Like Synoviocytes by Targeting Follistatin-Like Protein 1 in Rheumatoid Arthritis. Moleucles Cells 2016, 39, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gao, T.; Wang, Y. Geniposide Alleviates Lipopolysaccharide (LPS)-Induced Inflammation by Downregulation of MiR-27a in Rat Pancreatic Acinar Cell AR42J. Biol. Chem. 2019, 400, 1059–1068. [Google Scholar] [CrossRef]
- Wang, M.-Q.; Zhou, C.-H.; Cong, S.; Han, D.-X.; Wang, C.-J.; Tian, Y.; Zhang, J.-B.; Jiang, H.; Yuan, B. Lipopolysaccharide Inhibits Triglyceride Synthesis in Dairy Cow Mammary Epithelial Cells by Upregulating MiR-27a-3p, Which Targets the PPARG Gene. J. Dairy Sci. 2021, 104, 989–1001. [Google Scholar] [CrossRef]
- Iannotta, M.; Belardo, C.; Trotta, M.C.; Iannotti, F.A.; Vitale, R.M.; Maisto, R.; Boccella, S.; Infantino, R.; Ricciardi, F.; Mirto, B.F.; et al. N-Palmitoyl-D-Glucosamine, A Natural Monosaccharide-Based Glycolipid, Inhibits TLR4 and Prevents LPS-Induced Inflammation and Neuropathic Pain in Mice. Int. J. Mol. Sci. 2021, 22, 1491. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Signaling to NF-ΚB by Toll-like Receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, F.P.; de Faria Amormino, S.A.; Dutra, W.O.; Ribeiro Sobrinho, A.P.; Moreira, P.R. Methylation Pattern of the CD14 and TLR2 Genes in Human Dental Pulp. J. Endod. 2014, 40, 384–386. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.-M.; Song, Z.; Qin, W.; Li, J.; Li, W.-J.; Zhu, H.-Y.; Zhang, L. Expression of Nucleotide-Binding Oligomerization Domain 2 in Normal Human Dental Pulp Cells and Dental Pulp Tissues. J. Endod. 2009, 35, 838–842. [Google Scholar] [CrossRef]
- He, W.; Qu, T.; Yu, Q.; Wang, Z.; Lv, H.; Zhang, J.; Zhao, X.; Wang, P. LPS Induces IL-8 Expression through TLR4, MyD88, NF-KappaB and MAPK Pathways in Human Dental Pulp Stem Cells. Int. Endod. J. 2013, 46, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Feng, G.; Xing, J.; Shen, B.; Tan, W.; Huang, D.; Lu, X.; Tao, T.; Zhang, J.; Li, L.; et al. Repeated Lipopolysaccharide Stimulation Promotes Cellular Senescence in Human Dental Pulp Stem Cells (DPSCs). Cell Tissue Res. 2014, 356, 369–380. [Google Scholar] [CrossRef]
- Hirao, K.; Yumoto, H.; Takahashi, K.; Mukai, K.; Nakanishi, T.; Matsuo, T. Roles of TLR2, TLR4, NOD2, and NOD1 in Pulp Fibroblasts. J. Dent. Res. 2009, 88, 762–767. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, X.; Ding, X.; Wang, H.; Chen, X.; Zhao, H.; Jia, Y.; Liu, S.; Liu, Y. MiR-27 Inhibits Adipocyte Differentiation via Suppressing CREB Expression. Acta Biochim. Biophys. Sin. 2014, 46, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Li, S.; Zhu, Y.; Liu, L.; Pan, Y.; Wang, J.; Shi, W.; Song, Y.; Yang, L.; Gao, L.; et al. MicroRNA-27a/b Mediates Endothelin-1-Induced PPARγ Reduction and Proliferation of Pulmonary Artery Smooth Muscle Cells. Cell Tissue Res. 2017, 369, 527–539. [Google Scholar] [CrossRef]
- Nara, K.; Kawashima, N.; Noda, S.; Fujii, M.; Hashimoto, K.; Tazawa, K.; Okiji, T. Anti-inflammatory Roles of MicroRNA 21 in Lipopolysaccharide-stimulated Human Dental Pulp Cells. J. Cell Physiol. 2019, 234, 21331–21341. [Google Scholar] [CrossRef]
- Han, P.; Sunada-Nara, K.; Kawashima, N.; Fujii, M.; Wang, S.; Kieu, T.Q.; Yu, Z.; Okiji, T. MicroRNA-146b-5p Suppresses Pro-Inflammatory Mediator Synthesis via Targeting TRAF6, IRAK1, and RELA in Lipopolysaccharide-Stimulated Human Dental Pulp Cells. Int. J. Mol. Sci. 2023, 24, 7433. [Google Scholar] [CrossRef]
- Nishimoto, N. Interleukin-6 as a Therapeutic Target in Candidate Inflammatory Diseases. Clin. Pharmacol. Ther. 2010, 87, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Remick, D.G. Interleukin-8. Crit. Care Med. 2005, 33, S466. [Google Scholar] [CrossRef]
- Silva, T.A.; Garlet, G.P.; Fukada, S.Y.; Silva, J.S.; Cunha, F.Q. Chemokines in Oral Inflammatory Diseases: Apical Periodontitis and Periodontal Disease. J. Dent. Res. 2007, 86, 306–319. [Google Scholar] [CrossRef]
- Kilikevicius, A.; Meister, G.; Corey, D.R. Reexamining Assumptions about MiRNA-Guided Gene Silencing. Nucleic Acids Res. 2022, 50, 617–634. [Google Scholar] [CrossRef] [PubMed]
- Roh, Y.S.; Song, J.; Seki, E. TAK1 Regulates Hepatic Cell Survival and Carcinogenesis. J. Gastroenterol. 2014, 49, 185–194. [Google Scholar] [CrossRef]
- Mendoza, H.; Campbell, D.G.; Burness, K.; Hastie, J.; Ronkina, N.; Shim, J.-H.; Arthur, J.S.C.; Davis, R.J.; Gaestel, M.; Johnson, G.L.; et al. Roles for TAB1 in Regulating the IL-1-Dependent Phosphorylation of the TAB3 Regulatory Subunit and Activity of the TAK1 Complex. Biochem. J. 2008, 409, 711–722. [Google Scholar] [CrossRef]
- Yuan, L.; Bu, S.; Du, M.; Wang, Y.; Ju, C.; Huang, D.; Xu, W.; Tan, X.; Liang, M.; Deng, S.; et al. RNF207 Exacerbates Pathological Cardiac Hypertrophy via Post-Translational Modification of TAB1. Cardiovasc. Res. 2022, cvac039. [Google Scholar] [CrossRef] [PubMed]
- Pathak, S.; Borodkin, V.S.; Albarbarawi, O.; Campbell, D.G.; Ibrahim, A.; van Aalten, D.M. O-GlcNAcylation of TAB1 Modulates TAK1-Mediated Cytokine Release. EMBO J. 2012, 31, 1394–1404. [Google Scholar] [CrossRef]
- Zhao, J.; Cai, B.; Shao, Z.; Zhang, L.; Zheng, Y.; Ma, C.; Yi, F.; Liu, B.; Gao, C. TRIM26 Positively Regulates the Inflammatory Immune Response through K11-Linked Ubiquitination of TAB1. Cell Death Differ. 2021, 28, 3077–3091. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.-S. Therapeutic Advances of MiRNAs: A Preclinical and Clinical Update. J. Adv. Res. 2021, 28, 127–138. [Google Scholar] [CrossRef]
- Kawashima, N.; Okiji, T. Characteristics of Inflammatory Mediators in Dental Pulp Inflammation and the Potential for Their Control. Front. Dent. Med. 2024, 5, 1426887. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse | Accession No. | Size, bp |
|---|---|---|---|---|
| <human> | ||||
| ACTB | 5′-GTAGCACAGCTTCTCCTTAATGTCA-3′ | 5′-CTGACTGACTACCTCATGAAGATCC-3′ | NM_001101.3 | 102 |
| IL6 | 5′-TATACCTCAAACTCCAAAAGACCAG-3′ | 5′-ACAAGAGTAACATGTGTGAAAGCAG-3′ | NM_000600.4 | 157 |
| IL8 | 5′-TCAGTGCATAAAGACATACTCCAAA-3′ | 5′-TCTTCCATCAGAAAGCTTTACAATAA-3′ | NM_000584.4 | 121 |
| MCP1 | 5′-CACCTGCTGTTATAACTTCACCAAT-3′ | 5′-GTTGAAAGATGATAAGCCCACTCTA-3′ | NM_002982.4 | 130 |
| TAB1 | 5′-ATCCCTCAGTGCCAACTAAACC-3′ | 5′-GAAGATCCCAGTGCACAAGTCA-3′ | NM_153497.3 | 137 |
| IRAK4 | 5′-CGGAAATCTCTTTATCATCCGTGAG-3′ | 5′-GCACATATGTTGATGGTGTTATGGG-3′ | NM_001351341.2 | 126 |
| RELA | 5′-TTCCAAGTTCCTATAGAAGAGCAGC-3′ | 5′-TCAAAGATGGGATGAGAAAGGACAG-3′ | NM_021975.4 | 134 |
| FSTL1 | 5′-CCATGACCTGTGACGGAAAGAAT-3′ | 5′-TTAGATCTCTTTGGTGCTCACTCT-3′ | NM_007085.5 | 137 |
| <rat> | ||||
| Actb | 5′-GTAAAGACCTCTATGCCAACACAGT-3′ | 5′-GGAGCAATGATCTTGATCTTCATGG -3′ | NM_031144.3 | 127 |
| Il6 | 5′-TAAGGACCAAGACCATCCAACTCAT-3′ | 5′-AGTGAGGAATGTCCACAAACTGATA-3′ | NM_012589.2 | 125 |
| Mcp1 | 5′-CTAAGGACTTCAGCACCTTTGAATG-3′ | 5′-GTTCTCTGTCATACTGGTCACTTCT-3′ | NM_031530.1 | 120 |
| Tab1 | 5′-TAGTGTCTGCTTCTGTTAGATCCTG-3′ | 5′-AATCAGCTTCCTCATCAGAGTGAAA-3′ | NM_001109976.2 | 134 |
| Irak4 | 5′-CTGAACGTGCTTTGTCTTTAACAAG-3′ | 5′-GTTGAAATGAGCTCCGTACTAAGTG-3′ | NM_001106791.2 | 121 |
| Rela | 5′-CTTTCTCAAGTGCCTTAATAGCAGG-3′ | 5′-TTCAGAGCTAGAAAGAGCAAGAGTC-3′ | NM_199267.2 | 121 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Kawashima, N.; Han, P.; Sunada-Nara, K.; Yu, Z.; Tazawa, K.; Fujii, M.; Kieu, T.Q.; Okiji, T. MicroRNA-27a-5p Downregulates Expression of Proinflammatory Cytokines in Lipopolysaccharide-Stimulated Human Dental Pulp Cells via the NF-κB Signaling Pathway. Int. J. Mol. Sci. 2024, 25, 9694. https://doi.org/10.3390/ijms25179694
Wang S, Kawashima N, Han P, Sunada-Nara K, Yu Z, Tazawa K, Fujii M, Kieu TQ, Okiji T. MicroRNA-27a-5p Downregulates Expression of Proinflammatory Cytokines in Lipopolysaccharide-Stimulated Human Dental Pulp Cells via the NF-κB Signaling Pathway. International Journal of Molecular Sciences. 2024; 25(17):9694. https://doi.org/10.3390/ijms25179694
Chicago/Turabian StyleWang, Shihan, Nobuyuki Kawashima, Peifeng Han, Keisuke Sunada-Nara, Ziniu Yu, Kento Tazawa, Mayuko Fujii, Thoai Quoc Kieu, and Takashi Okiji. 2024. "MicroRNA-27a-5p Downregulates Expression of Proinflammatory Cytokines in Lipopolysaccharide-Stimulated Human Dental Pulp Cells via the NF-κB Signaling Pathway" International Journal of Molecular Sciences 25, no. 17: 9694. https://doi.org/10.3390/ijms25179694
APA StyleWang, S., Kawashima, N., Han, P., Sunada-Nara, K., Yu, Z., Tazawa, K., Fujii, M., Kieu, T. Q., & Okiji, T. (2024). MicroRNA-27a-5p Downregulates Expression of Proinflammatory Cytokines in Lipopolysaccharide-Stimulated Human Dental Pulp Cells via the NF-κB Signaling Pathway. International Journal of Molecular Sciences, 25(17), 9694. https://doi.org/10.3390/ijms25179694