
Metabolic Engineering of Escherichia coli for Production of a Bioactive Metabolite of Bilirubin

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. Construction of Pathway for BR Biosynthesis in E. coli
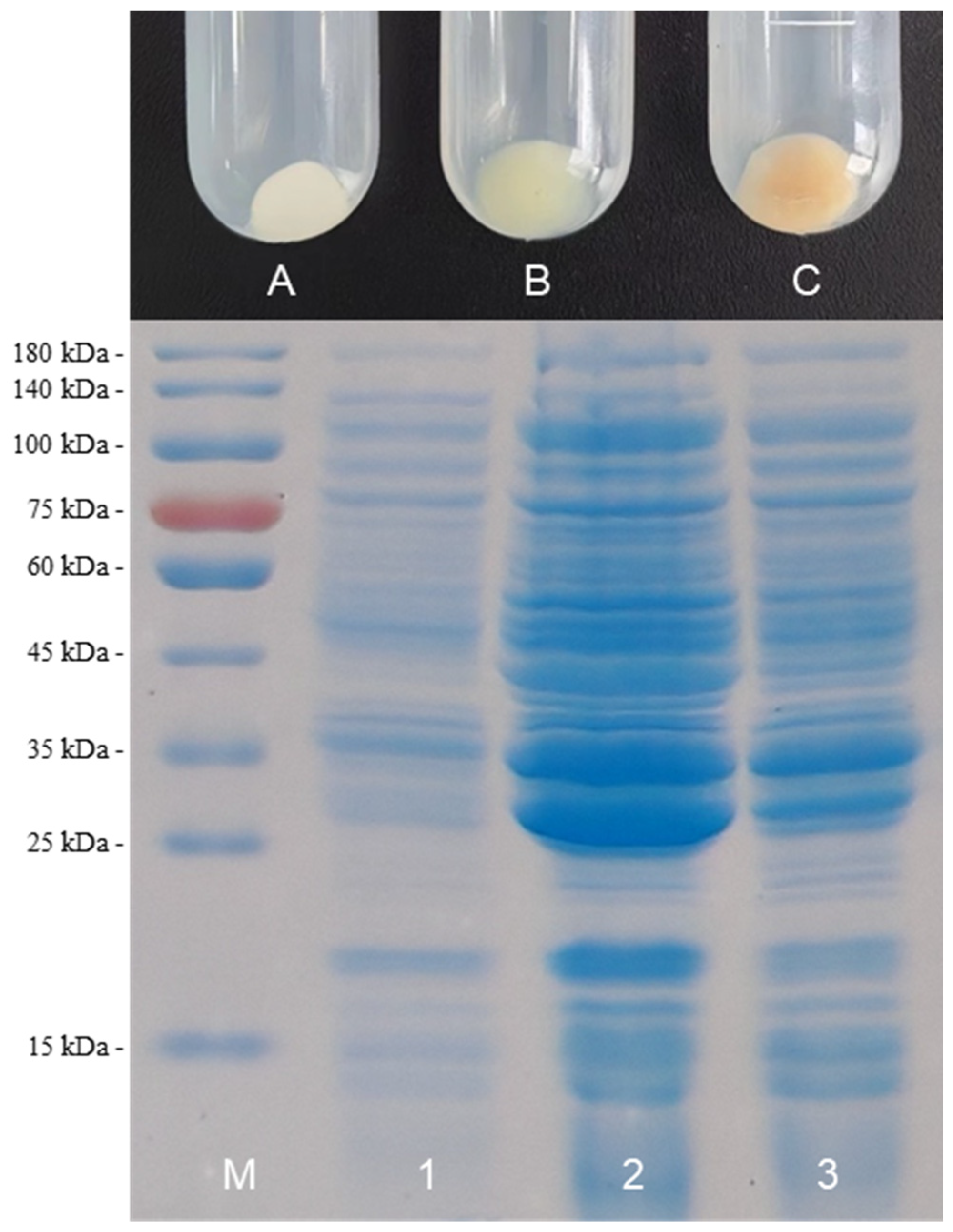
2.2. Screening of HO1 to Improve BR Production
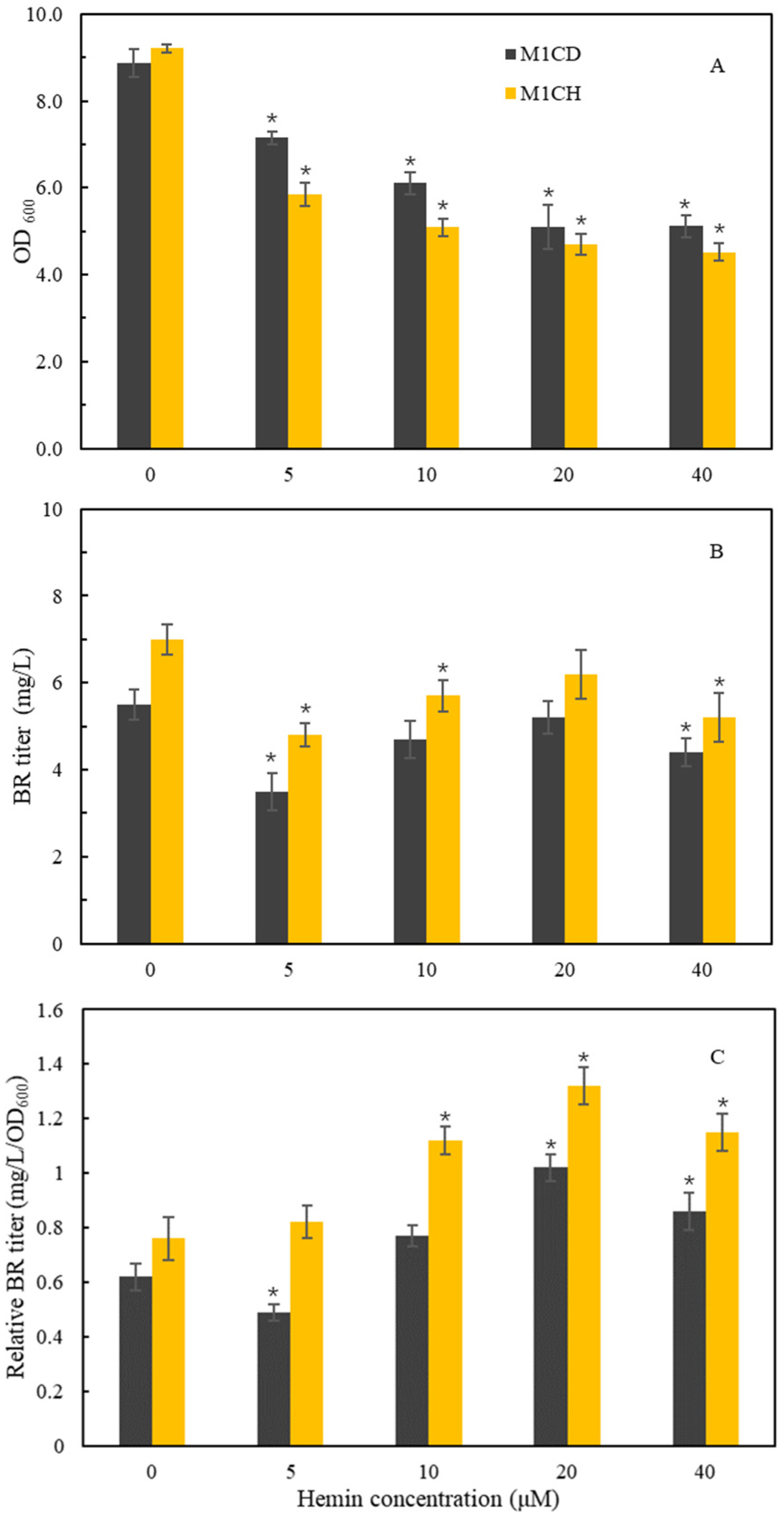
2.3. Effects of Heme Supplementation on BR Accumulation
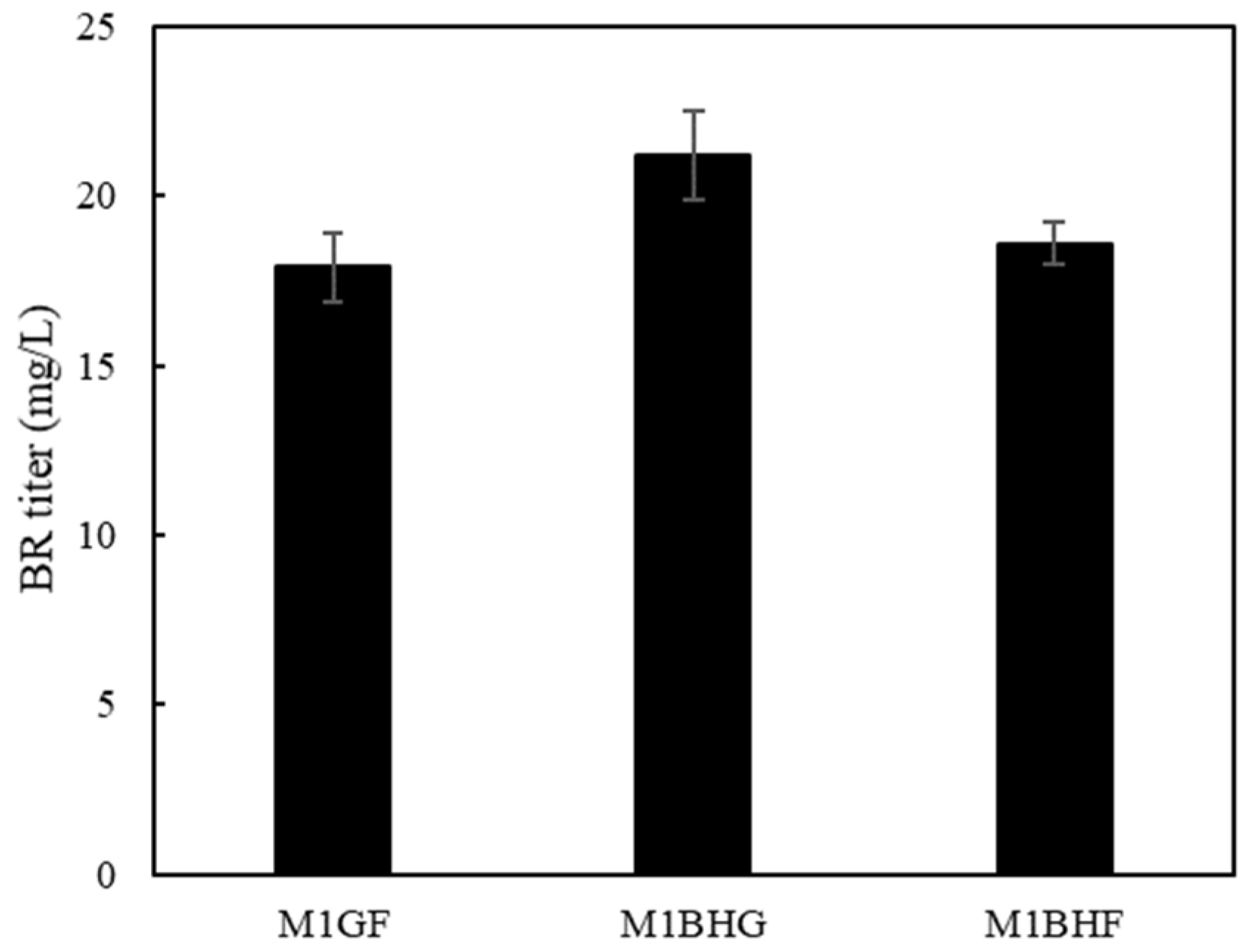
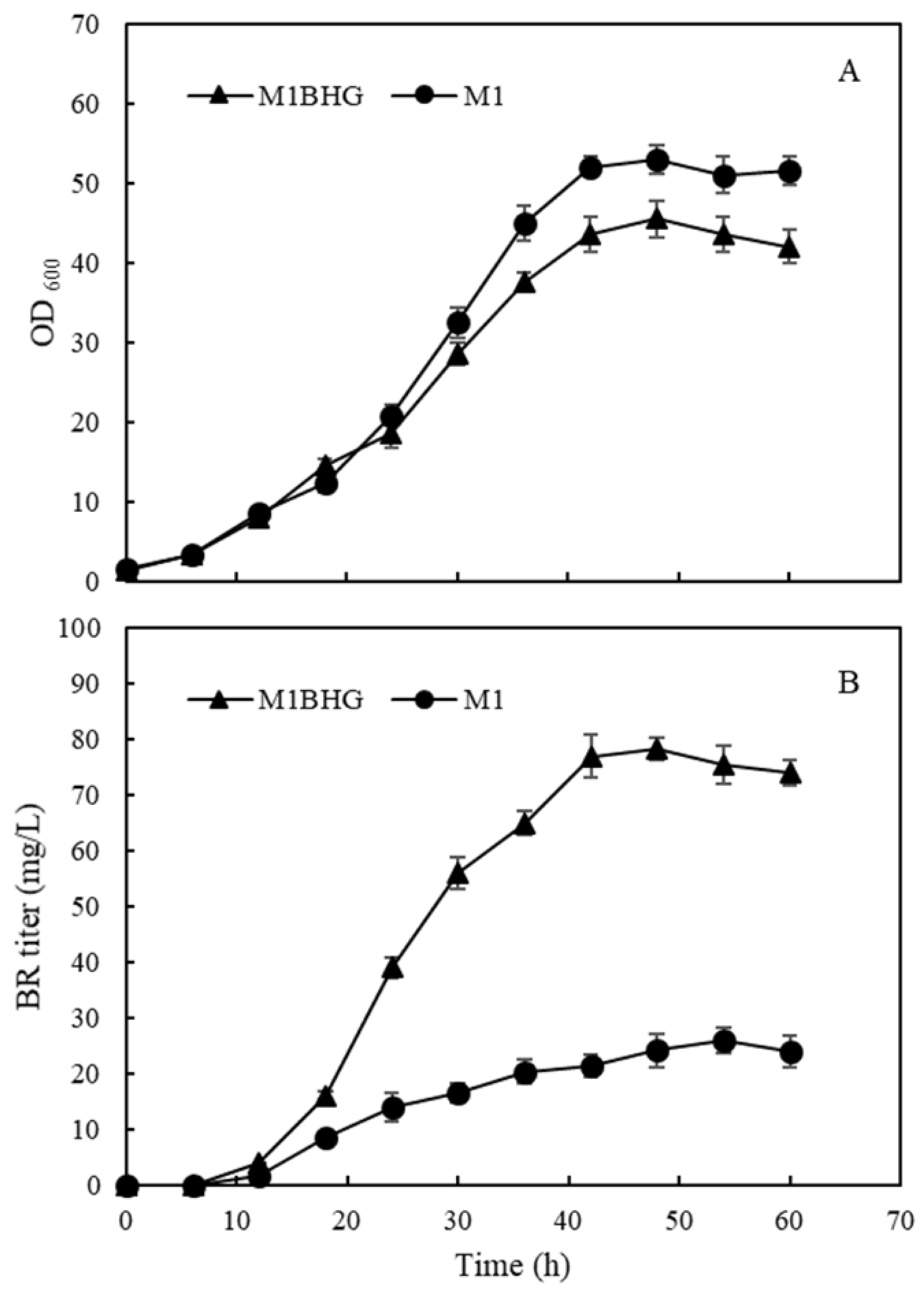
2.4. Regulation of Heme Biosynthesis and Its Effects on BR Biosynthesis
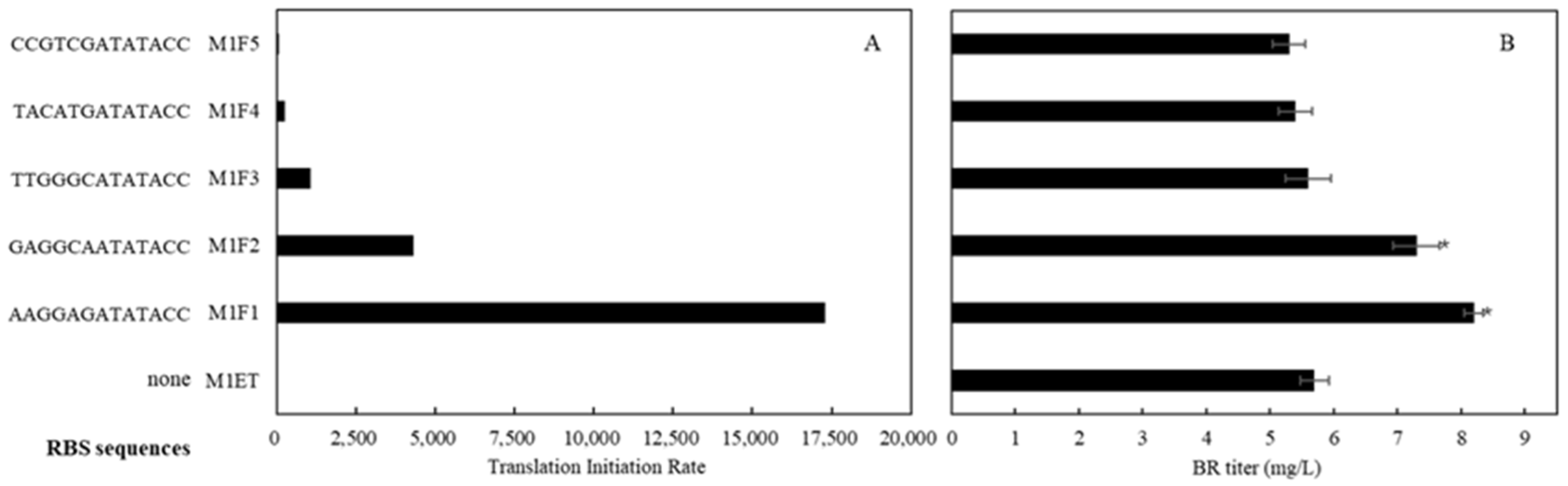
2.5. Regulation of BR Production by Ferredoxin (Fd) and Ferredoxin-NADP+ Reductase (FNR)
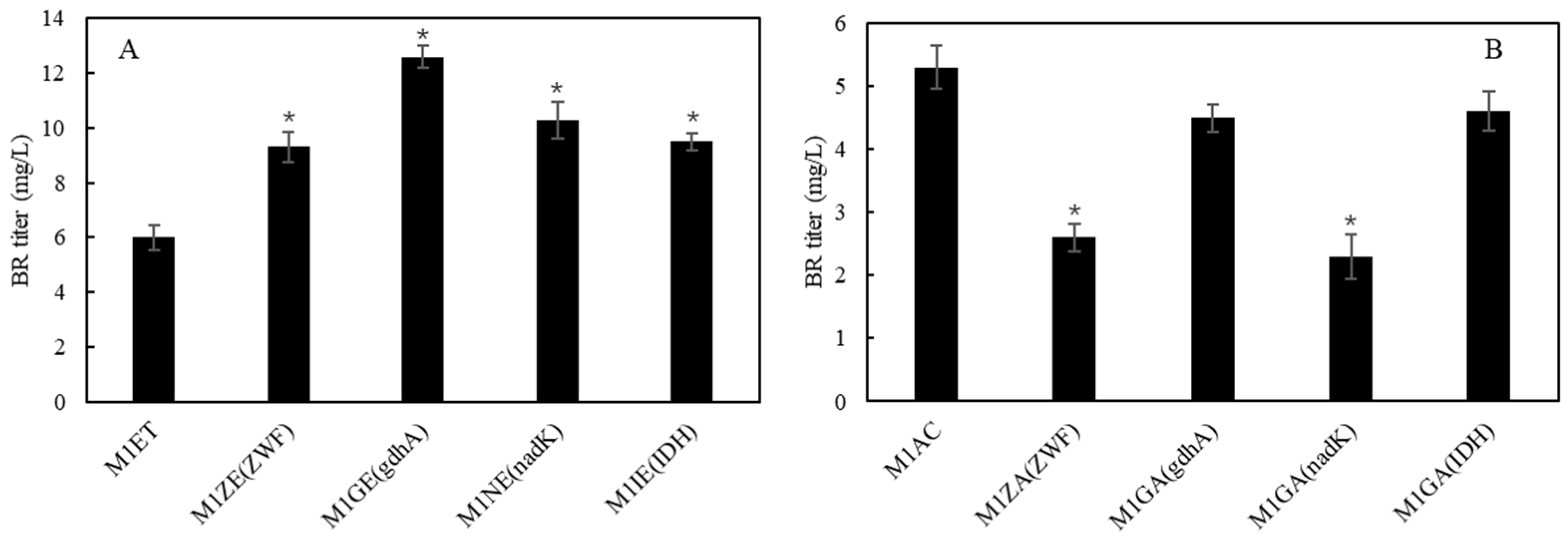
2.6. Overexpression of ZWF, nadK, gdhA and IDH for Improving NADPH Supply on BR Biosynthesis
2.7. Modular Optimization of the Committed Genes for Further Improvement of BR Biosynthesis
2.8. Fed-Batch Fermentation in a 5 L Bioreactor
3. Conclusions
4. Materials and Methods
4.1. Chemicals
4.2. Plasmids and Strains
4.3. Media and Culture Conditions
4.4. SDS-PAGE Analysis
4.5. HPLC and LC–MS Analysis of BR
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adin, C.A. BR as a therapeutic molecule: Challenges and opportunities. Antioxidants 2021, 10, 1536. [Google Scholar] [CrossRef] [PubMed]
- Ollinger, R.; Wang, H.; Yamashita, K.; Wegiel, B.; Thomas, M.; Margreiter, R.; Bach, F.H. Therapeutic applications of BR and biliverdin in transplantation. Antioxid. Redox. Signal. 2007, 9, 2175–2185. [Google Scholar] [CrossRef] [PubMed]
- Vítek, L.; Tiribelli, C. Bilirubin: The yellow hormone? J. Hepatol. 2021, 75, 1485–1490. [Google Scholar] [CrossRef]
- Vítek, L. The role of bilirubin in diabetes, metabolic syndrome, and cardiovascular diseases. Front. Pharmacol. 2012, 3, 55. [Google Scholar] [CrossRef]
- Vítek, L. Bilirubin and atherosclerotic diseases. Physiol. Res. 2017, 66, S11–S20. [Google Scholar] [CrossRef]
- Gazzin, S.; Vítek, L.; Watchko, J.; Shapiro, S.M.; Tiribelli, C. A novel perspective on the biology of bilirubin in health and disease. Trends. Mol. Med. 2016, 22, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.J.; Xu, Y.; Peng, W.; Liu, Y.J.; Zhang, J.M.; Li, J.S.; Sun, T.; Wang, P. Calculus bovis: A review of the traditional usages, origin, chemistry, pharmacological activities and toxicology. J. Ethnopharmacol. 2020, 254, 112649. [Google Scholar] [CrossRef] [PubMed]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of People’s Republic of China; China Medical Science Press: Beijing, China, 2020. [Google Scholar]
- Chen, F.; Chen, Y.; Luo, Z.; Pan, F.; Zhao, X.; Zhou, J.; Chen, R.; Eisen, M.S. Efficient Total Synthesis of Bilirubin IXα from Butyrolactone. Org. Process Res. Dev. 2024, 28, 2777–2788. [Google Scholar] [CrossRef]
- Huang, S.Y.; Hsu, R.P. Separation and purification of bilirubin from hog bile. In Biochemical Engineering for 2001; Furusaki, S., Endo, I., Matsuno, R., Eds.; Springer: Tokyo, Japan, 1992; pp. 551–553. [Google Scholar]
- Cao, H. Production Method for Extracting Bilirubin by Promoting Degradation by Animal Blood In Vitro Enzyme. China Invention Patent CN101487032A, 21 September 2011. [Google Scholar]
- Mei, J.; Wu, X.; Zheng, S.; Yi, Y.; Wang, X.; Ying, G. Production of BR by biotransformation of biliverdin using recombinant Escherichia coli cells. Bioprocess Biosyst Eng. 2022, 45, 563–571. [Google Scholar] [CrossRef]
- Wang, Y.; Li, N.; Shan, X.; Zhao, X.; Sun, Y.; Zhou, J. Enhancement of phycocyanobilin biosynthesis in Escherichia coli by strengthening the supply of precursor and artificially self-assembly complex. Synth. Syst. Biotechnol. 2023, 8, 227–234. [Google Scholar] [CrossRef]
- Yu, F.; Wang, Z.; Zhang, Z.; Zhou, J.; Li, J.; Chen, J.; Du, G.; Zhao, X. Biosynthesis, acquisition, regulation, and upcycling of heme: Recent advances. Crit. Rev. Biotechnol. 2024, 1–17. [Google Scholar] [CrossRef]
- Fiege, K.; Querebillo, C.J.; Hildebrandt, P.; Frankenberg-Dinkel, N. Improved Method for the Incorporation of Heme Cofactors into Recombinant Proteins Using Escherichia coli Nissle 1917. Biochemistry 2018, 57, 2747–2755. [Google Scholar] [CrossRef] [PubMed]
- Varnado, C.L.; Goodwin, D.C. System for the expression of recombinant hemoproteins in Escherichia coli. Protein. Expr. Purif. 2004, 35, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Ge, B.; Chen, Y.; Yu, Q.; Lin, X.; Li, J.; Qin, S. Regulation of the heme biosynthetic pathway for combinational biosynthesis of phycocyanobilin in Escherichia coli. Process Biochem. 2018, 71, 23–30. [Google Scholar] [CrossRef]
- Schobert, M.; Jahn, D. Regulation of heme biosynthesis in non-phototrophic bacteria. J. Mol. Microbiol. Biotechnol. 2002, 4, 287–294. [Google Scholar] [PubMed]
- Kery, V.; Elleder, D.; Kraus, J.P. Delta-aminolevulinate increases heme saturation and yield of human cystathionine beta-synthase expressed in Escherichia coli. Arch. Biochem. Biophys. 1995, 316, 24–29. [Google Scholar] [CrossRef]
- Wang, L.; Wilson, S.; Elliott, T.A. Mutant HemA protein with positive charge close to the N terminus is stabilized against heme-regulated proteolysis in Salmonella typhimurium. J. Bacteriol. 1999, 181, 6033–6041. [Google Scholar] [CrossRef]
- Zhang, J.; Kang, Z.; Chen, J.; Du, G. Optimization of the heme biosynthesis pathway for the production of 5-aminolevulinic acid in Escherichia coli. Sci. Rep. 2015, 26, 8584. [Google Scholar] [CrossRef]
- Zhao, X.R.; Choi, K.R.; Lee, S.Y. Metabolic engineering of Escherichia coli for secretory production of free haem. Nat. Catal. 2018, 1, 720–728. [Google Scholar] [CrossRef]
- Uda, Y.; Goto, Y.; Oda, S.; Kohchi, T.; Matsuda, M.; Aoki, K. Efficient synthesis of phycocyanobilin in mammalian cells for optogenetic control of cell signaling. Proc. Natl. Acad. Sci. USA 2017, 114, 11962–11967. [Google Scholar] [CrossRef]
- Kyriakakis, P.; Catanho, M.; Hoffner, N.; Thavarajah, W.; Hu, V.J.; Chao, S.S.; Hsu, A.; Pham, V.; Naghavian, L.; Dozier, L.E.; et al. Biosynthesis of Orthogonal Molecules Using Ferredoxin and Ferredoxin-NADP+ Reductase Systems Enables Genetically Encoded PhyB Optogenetics. ACS. Synth. Biol. 2018, 7, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Salis, H.M.; Mirsky, E.A.; Voigt, C.A. Automated design of synthetic ribosome binding sites to control protein expression. Nat. Biotechnol. 2009, 27, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Wang, S.; Bilal, M.; Ge, X.; Zhang, C.; Fickers, P.; Cheng, H. Identification, characterization of two NADPH-dependent erythrose reductases in the yeast Yarrowia lipolytica and improvement of erythritol productivity using metabolic engineering. Microb. Cell. Fact 2018, 17, 133. [Google Scholar] [CrossRef]
- Li, Z.; Ding, D.; Wang, H.; Liu, L.; Fang, H.; Chen, T.; Zhang, D. Engineering Escherichia coli to improve tryptophan production via genetic manipulation of precursor and cofactor pathways. Synth. Syst. Biotechnol. 2020, 5, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.; Farasat, I.; Maranas, C.D.; Salis, H.M. Rational design of a synthetic Entner–Doudoroff pathway for improved and controllable NADPH regeneration. Metab. Eng. 2015, 29, 86–96. [Google Scholar] [CrossRef]
- Mathur, A.; Chand, S. Model-based evaluation of plasmid segregational instability in repeated batch culture with recombinant Escherichia coli. Chem. Eng. J. 2009, 153, 227–230. [Google Scholar] [CrossRef]
- Popov, M.; Petrov, S.; Nacheva, G.; Ivanov, I.; Reichl, U. Effects of a recombinant gene expression on ColE1-like plasmid segregation in Escherichia coli. BMC Biotechnol. 2011, 11, 18. [Google Scholar] [CrossRef]
- Krysenko, S. Impact of nitrogen-containing compounds on secondary metabolism in Streptomyces spp.—A source of metabolic engineering strategies. SynBio 2023, 1, 204–225. [Google Scholar] [CrossRef]
- Chen, H.; Jiang, P. Metabolic engineering of Escherichia coli for efficient biosynthesis of fluorescent phycobiliprotein. Microb. Cell. Fact 2019, 18, 58. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Xiong, P.; Guo, N.; Liu, Z. Metabolic Engineering of Escherichia coli for Production of a Bioactive Metabolite of Bilirubin. Int. J. Mol. Sci. 2024, 25, 9741. https://doi.org/10.3390/ijms25179741
Chen H, Xiong P, Guo N, Liu Z. Metabolic Engineering of Escherichia coli for Production of a Bioactive Metabolite of Bilirubin. International Journal of Molecular Sciences. 2024; 25(17):9741. https://doi.org/10.3390/ijms25179741
Chicago/Turabian StyleChen, Huaxin, Peng Xiong, Ning Guo, and Zhe Liu. 2024. "Metabolic Engineering of Escherichia coli for Production of a Bioactive Metabolite of Bilirubin" International Journal of Molecular Sciences 25, no. 17: 9741. https://doi.org/10.3390/ijms25179741
APA StyleChen, H., Xiong, P., Guo, N., & Liu, Z. (2024). Metabolic Engineering of Escherichia coli for Production of a Bioactive Metabolite of Bilirubin. International Journal of Molecular Sciences, 25(17), 9741. https://doi.org/10.3390/ijms25179741