
Insights into the Detoxification of Spruce Monoterpenes by the Eurasian Spruce Bark Beetle
 ,
,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Results
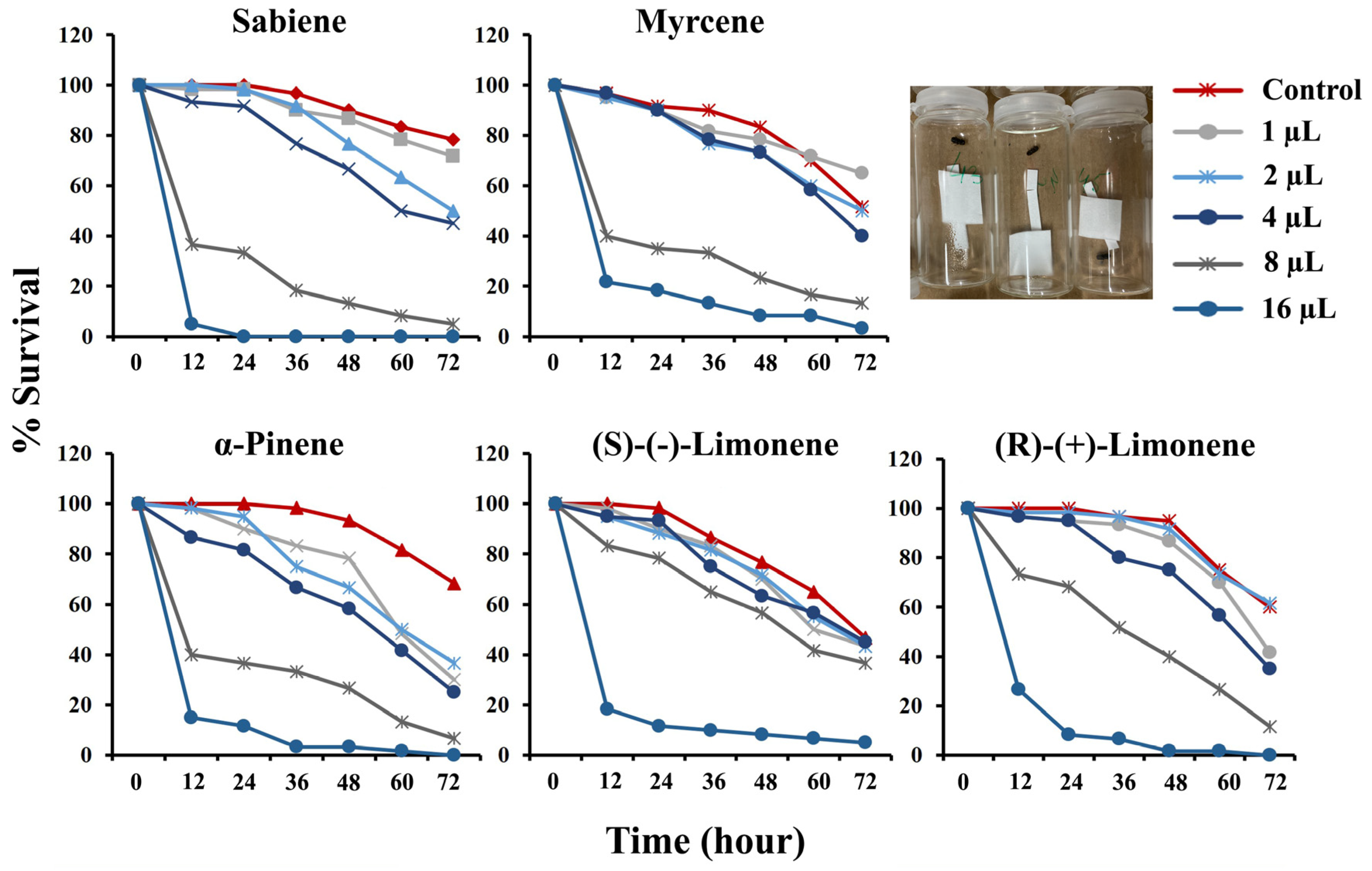
2.1. Toxicity Assay
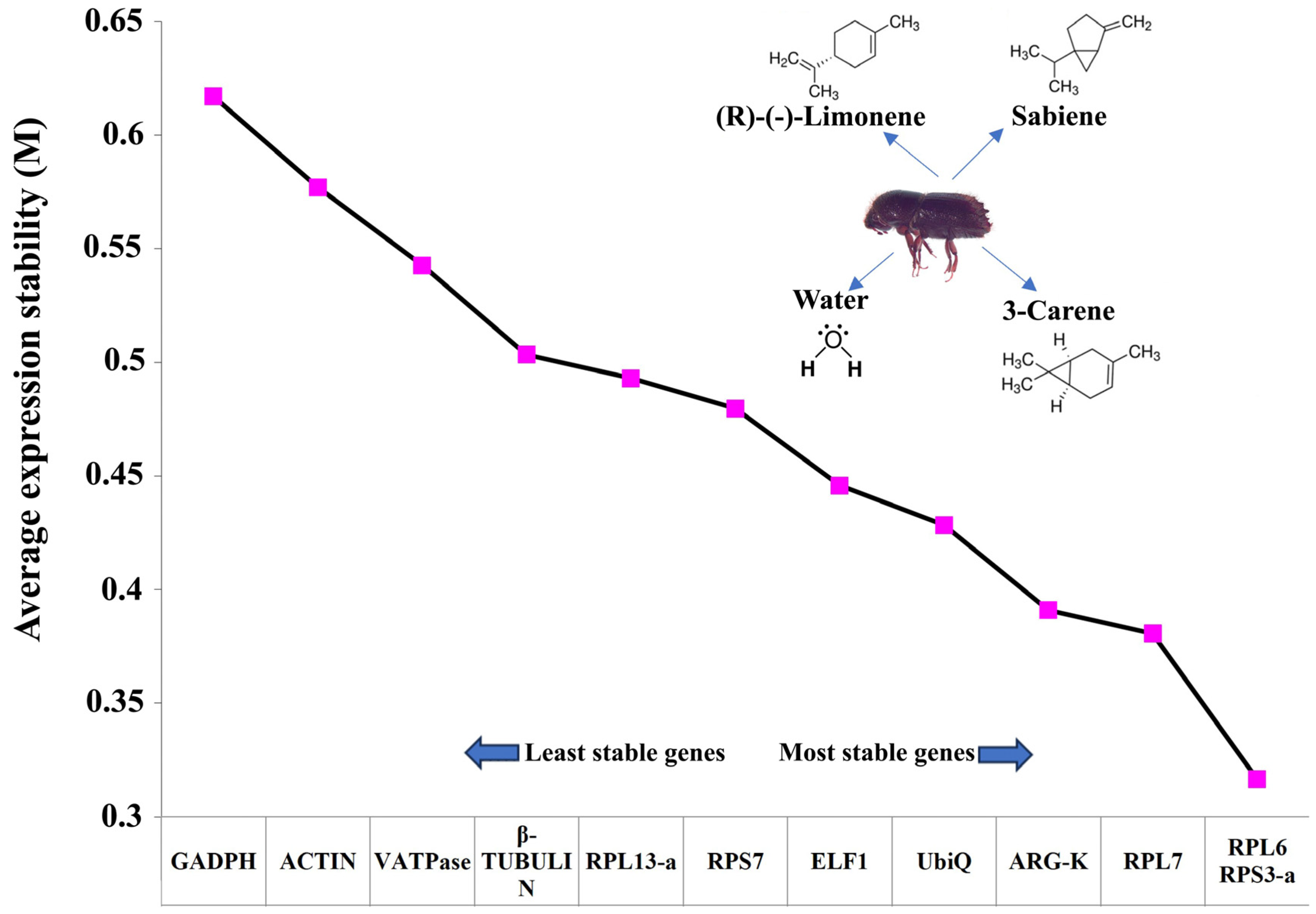
2.2. Reference Gene Selection
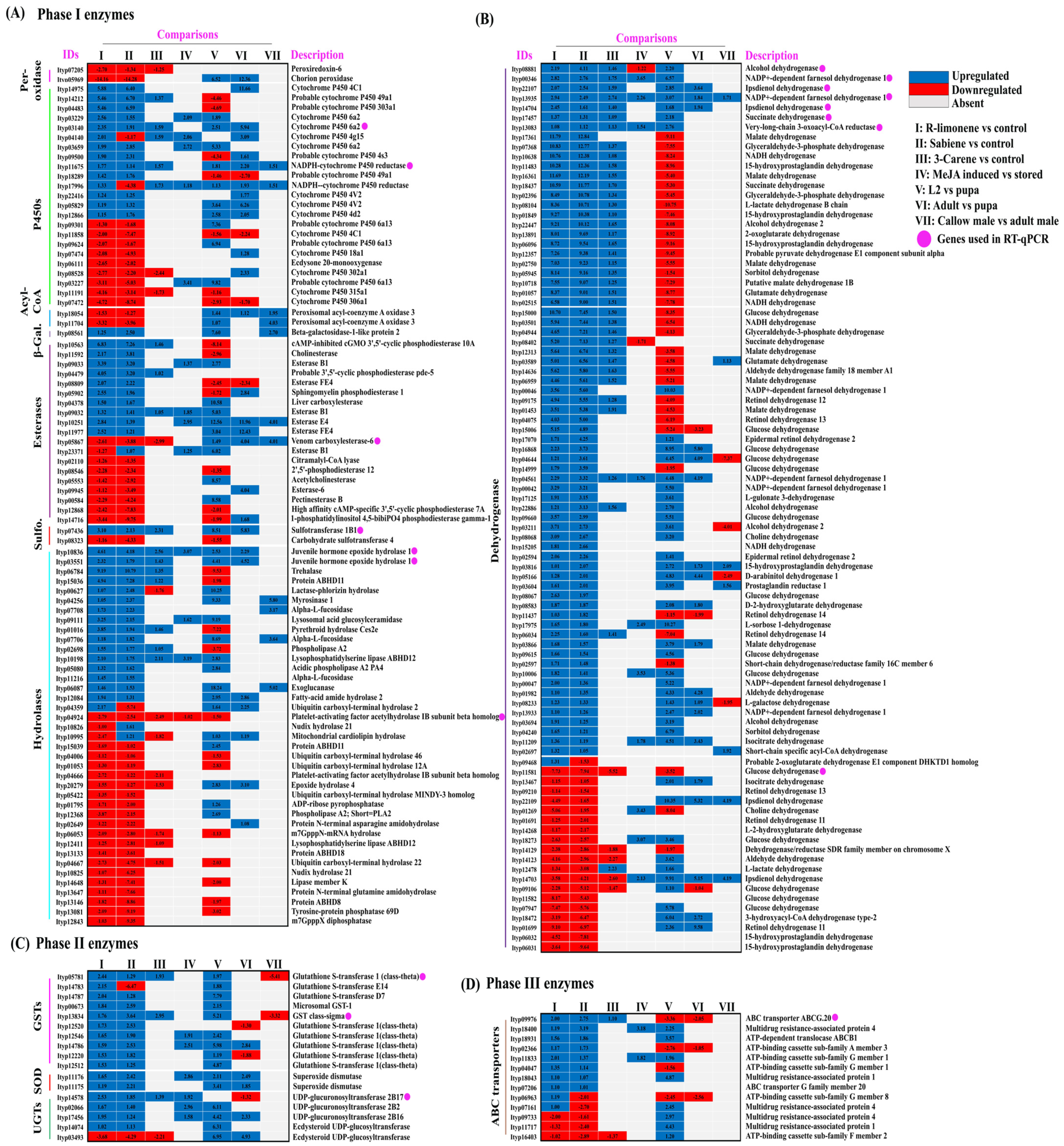
2.3. Differential Gene Expression Analysis
2.3.1. R-Limonene Treatment vs. Control
2.3.2. Sabiene Treatment vs. Control
2.4. RT-qPCR Analysis and Enzymatic Assay Analysis
3. Discussion
3.1. Expression of Phase I Detoxification Enzymes after Monoterpene Exposure
3.2. Expression of Phase II Detoxification Enzymes after Monoterpene Exposure
3.3. Expression of Phase III Detoxification Enzyme after Monoterpene Exposure
4. Materials and Methods
4.1. Insect Collection and Rearing
4.2. Fumigation Bioassay and Toxicity Calculation
4.3. Total RNA Extraction, cDNA Synthesis, and RT-qPCR Analysis
4.4. Reference Gene Selection
4.5. RNAseq Analyses
4.6. Comparison between Multiple In-House Data
4.7. Enzyme Activity Assay
4.8. Statistical Analysis
5. Study Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erbilgin, N.; Zanganeh, L.; Klutsch, J.G.; Chen, S.; Zhao, S.; Ishangulyyeva, G.; Burr, S.J.; Gaylord, M.; Hofstetter, R.; Keefover-Ring, K. Combined drought and bark beetle attacks deplete non-structural carbohydrates and promote death of mature pine trees. Plant Cell Environ. 2021, 44, 3866–3881. [Google Scholar] [CrossRef] [PubMed]
- Jakuš, R.; Edwards-Jonášová, M.; Cudlín, P.; Blaženec, M.; Ježík, M.; Havlíček, F.; Moravec, I. Characteristics of Norway spruce trees (Picea abies) surviving a spruce bark beetle (Ips typographus L.) outbreak. Trees 2011, 25, 965–973. [Google Scholar] [CrossRef]
- Schiebe, C.; Hammerbacher, A.; Birgersson, G.; Witzell, J.; Brodelius, P.E.; Gershenzon, J.; Hansson, B.S.; Krokene, P.; Schlyter, F. Inducibility of chemical defenses in Norway spruce bark is correlated with unsuccessful mass attacks by the spruce bark beetle. Oecologia 2012, 170, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, D.; Zaman, R.; Nakamura, Y.; Zhao, T.; Hartmann, H.; Andersson, M.N.; Hammerbacher, A.; Gershenzon, J. Conifer-killing bark beetles locate fungal symbionts by detecting volatile fungal metabolites of host tree resin monoterpenes. PLoS Biol. 2023, 21, e3001887. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, D.; Gershenzon, J.; Andersson, M.N.; Hammerbacher, A. Volatile organic compounds influence the interaction of the Eurasian spruce bark beetle (Ips typographus) with its fungal symbionts. ISME J. 2019, 13, 1788–1800. [Google Scholar] [CrossRef] [PubMed]
- Hlásny, T.; Krokene, P.; Liebhold, A.; Montagné-Huck, C.; Müller, J.; Qin, H.; Raffa, K.; Schelhaas, M.; Seidl, R.; Svoboda, M. Living with Bark Beetles: Impacts, Outlook and Management Options; European Forest Institute: Joensuu, Finland, 2019. [Google Scholar]
- Lubojacký, J.; Lorenc, F.; Samek, M.; Knížek, M.; Liška, J. Hlavní problémy v ochraně lesa v Česku v roce 2021 a prognóza na rok 2022. In Škodliví Činitelé v Lesích Česka 2021/2022–Škody Zvěří; Sborník Referátů z Celostátního Semináře s Mezinárodní Účastí: Průhonice, Czech Republic, 2022; pp. 17–26. [Google Scholar]
- Singh, V.V.; Naseer, A.; Mogilicherla, K.; Trubin, A.; Zabihi, K.; Roy, A.; Jakuš, R.; Erbilgin, N. Understanding bark beetle outbreaks: Exploring the impact of changing temperature regimes, droughts, forest structure, and prospects for future forest pest management. Rev. Environ. Sci. Bio/Technol. 2024, 23, 257–290. [Google Scholar] [CrossRef]
- Keeling, C.I.; Tittiger, C.; MacLean, M.; Blomquist, G.J. Pheromone production in bark beetles. In Insect Pheromone Biochemistry and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 123–162. [Google Scholar]
- Ramakrishnan, R.; Hradecký, J.; Roy, A.; Kalinová, B.; Mendezes, R.C.; Synek, J.; Bláha, J.; Svatoš, A.; Jirošová, A. Metabolomics and transcriptomics of pheromone biosynthesis in an aggressive forest pest Ips typographus. Insect Biochem. Mol. Biol. 2022, 140, 103680. [Google Scholar] [CrossRef]
- Krokene, P.; Solheim, H. Pathogenicity of four blue-stain fungi associated with aggressive and nonaggressive bark beetles. Phytopathology 1998, 88, 39–44. [Google Scholar] [CrossRef]
- Fäldt, J.; Solheim, H.; Långström, B.; Borg-Karlson, A.-K. Influence of fungal infection and wounding on contents and enantiomeric compositions of monoterpenes in phloem of Pinus sylvestris. J. Chem. Ecol. 2006, 32, 1779–1795. [Google Scholar] [CrossRef]
- Hammerbacher, A.; Schmidt, A.; Wadke, N.; Wright, L.P.; Schneider, B.; Bohlmann, J.; Brand, W.A.; Fenning, T.M.; Gershenzon, J.; Paetz, C. A common fungal associate of the spruce bark beetle metabolizes the stilbene defenses of Norway spruce. Plant Physiol. 2013, 162, 1324–1336. [Google Scholar] [CrossRef]
- Davis, T.S. The ecology of yeasts in the bark beetle holobiont: A century of research revisited. Microb. Ecol. 2015, 69, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Cale, J.A.; Ding, R.; Wang, F.; Rajabzadeh, R.; Erbilgin, N. Ophiostomatoid fungi can emit the bark beetle pheromone verbenone and other semiochemicals in media amended with various pine chemicals and beetle-released compounds. Fungal Ecol. 2019, 39, 285–295. [Google Scholar] [CrossRef]
- Chakraborty, A.; Modlinger, R.; Ashraf, M.Z.; Synek, J.; Schlyter, F.; Roy, A. Core mycobiome and their ecological relevance in the gut of five Ips bark beetles (Coleoptera: Curculionidae: Scolytinae). Front. Microbiol. 2020, 11, 568853. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Ashraf, M.Z.; Modlinger, R.; Synek, J.; Schlyter, F.; Roy, A. Unravelling the gut bacteriome of Ips (Coleoptera: Curculionidae: Scolytinae): Identifying core bacterial assemblage and their ecological relevance. Sci. Rep. 2020, 10, 18572. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Robert, C.A. Sequestration of plant secondary metabolites by insect herbivores: Molecular mechanisms and ecological consequences. Curr. Opin. Insect Sci. 2016, 14, 8–11. [Google Scholar] [CrossRef]
- Heckel, D.G. Insect detoxification and sequestration strategies. Annu. Plant Rev. Insect-Plant Interact. 2014, 47, 77–114. [Google Scholar]
- van Veen, F.F. Plant-modified trophic interactions. Curr. Opin. Insect Sci. 2015, 8, 29–33. [Google Scholar] [CrossRef]
- Ahmad, S. Enzymatic adaptations of herbivorous insects and mites to phytochemicals. J. Chem. Ecol. 1986, 12, 533–560. [Google Scholar] [CrossRef]
- Jin, M.; Liao, C.; Fu, X.; Holdbrook, R.; Wu, K.; Xiao, Y. Adaptive regulation of detoxification enzymes in Helicoverpa armigera to different host plants. Insect Mol. Biol. 2019, 28, 628–636. [Google Scholar] [CrossRef]
- Hilliou, F.; Chertemps, T.; Maïbèche, M.; Le Goff, G. Resistance in the genus Spodoptera: Key insect detoxification genes. Insects 2021, 12, 544. [Google Scholar] [CrossRef]
- Kshatriya, K.; Gershenzon, J. Disarming the defenses: Insect detoxification of plant defense-related specialized metabolites. Curr. Opin. Plant Biol. 2024, 81, 102577. [Google Scholar] [CrossRef] [PubMed]
- Boyland, E.; Chasseaud, L. The role of glutathione and glutathione S-transferases in mercapturic acid biosynthesis. Adv. Enzymol. Relat. Areas Mol. Biol. 1969, 32, 173–219. [Google Scholar] [PubMed]
- Jakoby, W.; Ziegler, D. The enzymes of detoxication. J. Biol. Chem. 1990, 265, 20715–20718. [Google Scholar] [CrossRef] [PubMed]
- Commandeur, J.; Stijntjes, G.J.; Vermeulen, N. Enzymes and transport systems involved in the formation and disposition of glutathione S-conjugates. Role in bioactivation and detoxication mechanisms of xenobiotics. Pharmacol. Rev. 1995, 47, 271–330. [Google Scholar]
- König, J.; Nies, A.T.; Cui, Y.; Leier, I.; Keppler, D. Conjugate export pumps of the multidrug resistance protein (MRP) family: Localization, substrate specificity, and MRP2-mediated drug resistance. Biochim. Biophys. Acta (BBA)-Biomembr. 1999, 1461, 377–394. [Google Scholar] [CrossRef]
- Váradi, A.; Sarkadi, B. Multidrug resistance-associated proteins: Export pumps for conjugates with glutathione, glucuronate or sulfate. Biofactors 2003, 17, 103–114. [Google Scholar]
- Powell, D.; Groβe-Wilde, E.; Krokene, P.; Roy, A.; Chakraborty, A.; Löfstedt, C.; Vogel, H.; Andersson, M.N.; Schlyter, F. A highly-contiguous genome assembly of the Eurasian spruce bark beetle, Ips typographus, provides insight into a major forest pest. Commun. Biol. 2021, 4, 1059. [Google Scholar] [CrossRef]
- Naseer, A.; Mogilicherla, K.; Sellamuthu, G.; Roy, A. Age matters: Life-stage, tissue, and sex-specific gene expression dynamics in Ips typographus (Coleoptera: Curculionidae: Scolytinae). Front. For. Glob. Chang. 2023, 6, 1124754. [Google Scholar] [CrossRef]
- Sellamuthu, G.; Naseer, A.; Hradecký, J.; Chakraborty, A.; Synek, J.; Modlinger, R.; Roy, A. Gene expression plasticity facilitates different host feeding in Ips sexdentatus (Coleoptera: Curculionidae: Scolytinae). Insect Biochem. Mol. Biol. 2024, 165, 104061. [Google Scholar] [CrossRef]
- Dai, L.; Ma, M.; Wang, C.; Shi, Q.; Zhang, R.; Chen, H. Cytochrome P450s from the Chinese white pine beetle, Dendroctonus armandi (Curculionidae: Scolytinae): Expression profiles of different stages and responses to host allelochemicals. Insect Biochem. Mol. Biol. 2015, 65, 35–46. [Google Scholar] [CrossRef]
- López, M.F.; Cano-Ramírez, C.; Cesar-Ayala, A.K.; Ruiz, E.A.; Zúñiga, G. Diversity and expression of P450 genes from Dendroctonus valens LeConte (Curculionidae: Scolytinae) in response to different kairomones. Insect Biochem. Mol. Biol. 2013, 43, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Sarabia, L.E.; López, M.F.; Pineda-Mendoza, R.M.; Obregón-Molina, G.; Gonzalez-Escobedo, R.; Albores-Medina, A.; Zúñiga, G. Time-course of CYP450 Genes expression from Dendroctonus rhizophagus (Curculionidae: Scolytinae) during early hours of drilling bark and settling into the host tree. J. Insect Sci. 2019, 19, 11. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.C.; Keeling, C.I.; Bohlmann, J. Toxicity of pine monoterpenes to mountain pine beetle. Sci. Rep. 2017, 7, 8858. [Google Scholar] [CrossRef] [PubMed]
- Henderson, C.F.; TILTON, E.W. Tests with acaricides against the brown wheat mite. J. Econ. Entomol. 1955, 48, 157–161. [Google Scholar] [CrossRef]
- Sellamuthu, G.; Bílý, J.; Joga, M.R.; Synek, J.; Roy, A. Identifying optimal reference genes for gene expression studies in Eurasian spruce bark beetle, Ips typographus (Coleoptera: Curculionidae: Scolytinae). Sci. Rep. 2022, 12, 4671. [Google Scholar] [CrossRef]
- Hopkins, R.J.; van Dam, N.M.; van Loon, J.J. Role of glucosinolates in insect-plant relationships and multitrophic interactions. Annu. Rev. Entomol. 2009, 54, 57–83. [Google Scholar] [CrossRef]
- Ashraf, M.Z.; Mogilicherla, K.; Sellamuthu, G.; Siino, V.; Levander, F.; Roy, A. Comparative gut proteomics study revealing adaptive physiology of Eurasian spruce bark beetle, Ips typographus (Coleoptera: Scolytinae). Front. Plant Sci. 2023, 14, 1157455. [Google Scholar] [CrossRef]
- Pentzold, S.; Zagrobelny, M.; Rook, F.; Bak, S. How insects overcome two-component plant chemical defence: Plant β-glucosidases as the main target for herbivore adaptation. Biol. Rev. 2014, 89, 531–551. [Google Scholar] [CrossRef]
- War, A.R.; Taggar, G.K.; Hussain, B.; Taggar, M.S.; Nair, R.M.; Sharma, H.C. Plant defence against herbivory and insect adaptations. AoB Plants 2018, 10, ply037. [Google Scholar]
- Mageroy, M.H.; Christiansen, E.; Långström, B.; Borg-Karlson, A.K.; Solheim, H.; Björklund, N.; Zhao, T.; Schmidt, A.; Fossdal, C.G.; Krokene, P. Priming of inducible defenses protects Norway spruce against tree-killing bark beetles. Plant Cell Environ. 2020, 43, 420–430. [Google Scholar] [CrossRef]
- Zhao, T.; Borg-Karlson, A.-K.; Erbilgin, N.; Krokene, P. Host resistance elicited by methyl jasmonate reduces emission of aggregation pheromones by the spruce bark beetle, Ips Typogr. Oecologia 2011, 167, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Krokene, P.; Hu, J.; Christiansen, E.; Björklund, N.; Långström, B.; Solheim, H.; Borg-Karlson, A.-K. Induced terpene accumulation in Norway spruce inhibits bark beetle colonization in a dose-dependent manner. PLoS ONE 2011, 6, e26649. [Google Scholar] [CrossRef] [PubMed]
- Krokene, P. Conifer defense and resistance to bark beetles. In Bark Beetles; Elsevier: Amsterdam, The Netherlands, 2015; pp. 177–207. [Google Scholar]
- Sun, J.; Miao, Z.; Zhang, Z.; Zhang, Z.; Gillette, N.E. Red turpentine beetle, Dendroctonus valens LeConte (Coleoptera: Scolytidae), response to host semiochemicals in China. Environ. Entomol. 2004, 33, 206–212. [Google Scholar] [CrossRef]
- Erbilgin, N.; Mori, S.; Sun, J.; Stein, J.; Owen, D.; Merrill, L.; Bolanos, R.C.; Raffa, K.; Montiel, T.M.; Wood, D. Response to host volatiles by native and introduced populations of Dendroctonus valens (Coleoptera: Curculionidae, Scolytinae) in North America and China. J. Chem. Ecol. 2007, 33, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Erbilgin, N.; Powell, J.S.; Raffa, K.F. Effect of varying monoterpene concentrations on the response of Ips pini (Coleoptera: Scolytidae) to its aggregation pheromone: Implications for pest management and ecology of bark beetles. Agric. For. Entomol. 2003, 5, 269–274. [Google Scholar] [CrossRef]
- Martin, D.; Tholl, D.; Gershenzon, J.; Bohlmann, J.r. Methyl jasmonate induces traumatic resin ducts, terpenoid resin biosynthesis, and terpenoid accumulation in developing xylem of Norway spruce stems. Plant Physiol. 2002, 129, 1003–1018. [Google Scholar] [CrossRef]
- Amezian, D.; Nauen, R.; Le Goff, G. Comparative analysis of the detoxification gene inventory of four major Spodoptera pest species in response to xenobiotics. Insect Biochem. Mol. Biol. 2021, 138, 103646. [Google Scholar] [CrossRef]
- Dai, L.; Gao, H.; Chen, H. Expression levels of detoxification enzyme genes from Dendroctonus armandi (Coleoptera: Curculionidae) fed on a solid diet containing pine phloem and terpenoids. Insects 2021, 12, 926. [Google Scholar] [CrossRef]
- BK, S.K.; Moural, T.; Zhu, F. Functional and structural diversity of insect glutathione S-transferases in xenobiotic adaptation. Int. J. Biol. Sci. 2022, 18, 5713. [Google Scholar]
- Müller, C.; Vogel, H.; Heckel, D.G. Transcriptional responses to short-term and long-term host plant experience and parasite load in an oligophagous beetle. Mol. Ecol. 2017, 26, 6370–6383. [Google Scholar] [CrossRef]
- Noriega, D.D.; Arraes, F.B.; Antonino, J.D.; Macedo, L.L.; Fonseca, F.C.; Togawa, R.C.; Grynberg, P.; Silva, M.C.; Negrisoli Jr, A.S.; Morgante, C.V. Comparative gut transcriptome analysis of Diatraea saccharalis in response to the dietary source. PLoS ONE 2020, 15, e0235575. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Fu, D.; Ning, H.; Tang, M.; Chen, H. Knockdown of CYP6CR2 and CYP6DE5 reduces tolerance to host plant allelochemicals in the Chinese white pine beetle Dendroctonus armandi. Pestic. Biochem. Physiol. 2022, 187, 105180. [Google Scholar] [CrossRef] [PubMed]
- Robert, J.A.; Pitt, C.; Bonnett, T.R.; Yuen, M.M.S.; Keeling, C.I.; Bohlmann, J.; Huber, D.P.W. Disentangling Detoxification: Gene Expression Analysis of Feeding Mountain Pine Beetle Illuminates Molecular-Level Host Chemical Defense Detoxification Mechanisms. PLoS ONE 2013, 8, e77777. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liao, M.; Yang, Q.Q.; Xiao, J.J.; Hu, Z.Y.; Zhou, L.J.; Cao, H.Q. Transcriptome profiling reveals differential gene expression of detoxification enzymes in Sitophilus zeamais responding to terpinen-4-ol fumigation. Pestic. Biochem. Physiol. 2018, 149, 44–53. [Google Scholar] [CrossRef]
- Huang, Y.; Liao, M.; Yang, Q.Q.; Xiao, J.J.; Hu, Z.Y.; Cao, H.Q. iTRAQ-based quantitative proteome revealed metabolic changes of Sitophilus zeamais in response to terpinen-4-ol fumigation. Pest Manag. Sci. 2019, 75, 444–451. [Google Scholar] [CrossRef]
- Kim, J.I.; Kwon, M.; Kim, G.H.; Kim, S.Y.; Lee, S.H. Two mutations in nAChR beta subunit is associated with imidacloprid resistance in the Aphis gossypii. J. Asia-Pac. Entomol. 2015, 18, 291–296. [Google Scholar] [CrossRef]
- Keeling, C.I.; Li, M.; Sandhu, H.K.; Henderson, H.; Saint Yuen, M.M.; Bohlmann, J. Quantitative metabolome, proteome and transcriptome analysis of midgut and fat body tissues in the mountain pine beetle, Dendroctonus ponderosae Hopkins, and insights into pheromone biosynthesis. Insect Biochem. Mol. Biol. 2016, 70, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Sun, Z.; Shi, Q.; Wang, R.; Xu, C.; Wang, H.; Song, Y.; Zeng, R. RNA-Seq analyses of midgut and fat body tissues reveal the molecular mechanism underlying Spodoptera litura resistance to tomatine. Front. Physiol. 2019, 10, 8. [Google Scholar] [CrossRef]
- Shen, X.; Liu, W.; Wan, F.; Lv, Z.; Guo, J. The role of cytochrome P450 4C1 and carbonic anhydrase 3 in response to temperature stress in Bemisia tabaci. Insects 2021, 12, 1071. [Google Scholar] [CrossRef]
- Chung, H.; Sztal, T.; Pasricha, S.; Sridhar, M.; Batterham, P.; Daborn, P.J. Characterization of Drosophila melanogaster cytochrome P450 genes. Proc. Natl. Acad. Sci. USA 2009, 106, 5731–5736. [Google Scholar] [CrossRef]
- Wan, H.; Liu, Y.; Li, M.; Zhu, S.; Li, X.; Pittendrigh, B.R.; Qiu, X. Nrf2/Maf-binding-site-containing functional Cyp6a2 allele is associated with DDT resistance in Drosophila melanogaster. Pest Manag. Sci. 2014, 70, 1048–1058. [Google Scholar] [CrossRef] [PubMed]
- Kang, Z.-W.; Liu, F.-H.; Pang, R.-P.; Tian, H.-G.; Liu, T.-X. Effect of sublethal doses of imidacloprid on the biological performance of aphid endoparasitoid Aphidius gifuensis (Hymenoptera: Aphidiidae) and influence on its related gene expression. Front. Physiol. 2018, 9, 1729. [Google Scholar] [CrossRef]
- Lien, N.T.K.; Ngoc, N.T.H.; Lan, N.N.; Hien, N.T.; Van Tung, N.; Ngan, N.T.T.; Hoang, N.H.; Binh, N.T.H. Transcriptome sequencing and analysis of changes associated with insecticide resistance in the dengue mosquito (Aedes aegypti) in Vietnam. Am. J. Trop. Med. Hyg. 2019, 100, 1240. [Google Scholar] [CrossRef]
- Tsuji, A.; Kotani, E.; Inoue, Y.H. Sesamin Exerts an Antioxidative Effect by Activating the Nrf2 Transcription Factor in the Glial Cells of the Central Nervous System in Drosophila Larvae. Antioxidants 2024, 13, 787. [Google Scholar] [CrossRef] [PubMed]
- Niwa, R.; Matsuda, T.; Yoshiyama, T.; Namiki, T.; Mita, K.; Fujimoto, Y.; Kataoka, H. CYP306A1, a cytochrome P450 enzyme, is essential for ecdysteroid biosynthesis in the prothoracic glands of Bombyx and Drosophila. J. Biol. Chem. 2004, 279, 35942–35949. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-L.; Wang, K.; Zhang, R.; Xu, Q.-Y.; Luo, G.-H.; Fnag, J.-C. Molecular cloning of Cyp302a1 and Cyp315a1 and expression profiles of halloween genes in Chilo suppressalis. J. Nanjing Agric. Univ. 2022, 45, 287–296. [Google Scholar]
- Li, J.; Kim, S.R.; Li, J. Molecular characterization of a novel peroxidase involved in Aedes aegypti chorion protein crosslinking. Insect Biochem. Mol. Biol. 2004, 34, 1195–1203. [Google Scholar] [CrossRef]
- Enayati, A.A.; Ranson, H.; Hemingway, J. Insect glutathione transferases and insecticide resistance. Insect Mol. Biol. 2005, 14, 3–8. [Google Scholar] [CrossRef]
- Song, X.W.; Zhong, Q.S.; Ji, Y.H.; Zhang, Y.M.; Tang, J.; Feng, F.; Bi, J.X.; Xie, J.; Li, B. Characterization of a sigma class GST (GSTS6) required for cellular detoxification and embryogenesis in Tribolium castaneum. Insect Sci. 2022, 29, 215–229. [Google Scholar] [CrossRef]
- Gao, H.; Dai, L.; Fu, D.; Sun, Y.; Chen, H. Isolation, expression profiling, and regulation via host allelochemicals of 16 Glutathione S-Transferases in the Chinese White Pine Beetle, Dendroctonus armandi. Front. Physiol. 2020, 11, 546592. [Google Scholar] [CrossRef]
- Wongtrakul, J.; Pongjaroenkit, S.; Leelapat, P.; Nachaiwieng, W.; Prapanthadara, L.-A.; Ketterman, A.J. Expression and characterization of three new glutathione transferases, an epsilon (AcGSTE2-2), omega (AcGSTO1-1), and theta (AcGSTT1-1) from Anopheles cracens (Diptera: Culicidae), a major Thai malaria vector. J. Med. Entomol. 2014, 47, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-H.; Wang, C.-C.; Hou, Y.-H.; Mao, Y.-C.; Yang, Y.-S. Mechanism of sulfotransferase pharmacogenetics in altered xenobiotic metabolism. Expert Opin. Drug Metab. Toxicol. 2015, 11, 1053–1071. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Hopkins, T. β-glucosylation of plant phenolics by phenol β-glucosyltransferase in larval tissues of the tobacco hornworm, Manduca sexta (L.). Insect Biochem. Mol. Biol. 1993, 23, 581–589. [Google Scholar] [CrossRef]
- Ahn, S.-J.; Vogel, H.; Heckel, D.G. Comparative analysis of the UDP-glycosyltransferase multigene family in insects. Insect Biochem. Mol. Biol. 2012, 42, 133–147. [Google Scholar] [CrossRef]
- Xu, X.; Wang, M.; Wang, Y.; Sima, Y.; Zhang, D.; Li, J.; Yin, W.; Xu, S. Green cocoons in silkworm Bombyx mori resulting from the quercetin 5-O-glucosyltransferase of UGT86, is an evolved response to dietary toxins. Mol. Biol. Rep. 2013, 40, 3631–3639. [Google Scholar] [CrossRef]
- Krempl, C.; Sporer, T.; Reichelt, M.; Ahn, S.-J.; Heidel-Fischer, H.; Vogel, H.; Heckel, D.G.; Joußen, N. Potential detoxification of gossypol by UDP-glycosyltransferases in the two Heliothine moth species Helicoverpa armigera and Heliothis virescens. Insect Biochem. Mol. Biol. 2016, 71, 49–57. [Google Scholar] [CrossRef]
- Roy, A.; Walker, W., III; Vogel, H.; Chattington, S.; Larsson, M.; Anderson, P.; Heckel, D.G.; Schlyter, F. Diet dependent metabolic responses in three generalist insect herbivores Spodoptera spp. Insect Biochem. Mol. Biol. 2016, 71, 91–105. [Google Scholar] [CrossRef]
- Chen, X.; Xia, J.; Shang, Q.; Song, D.; Gao, X. UDP-glucosyltransferases potentially contribute to imidacloprid resistance in Aphis gossypii glover based on transcriptomic and proteomic analyses. Pestic. Biochem. Physiol. 2019, 159, 98–106. [Google Scholar] [CrossRef]
- Pan, Y.; Xu, P.; Zeng, X.; Liu, X.; Shang, Q. Characterization of UDP-glucuronosyltransferases and the potential contribution to nicotine tolerance in Myzus persicae. Int. J. Mol. Sci. 2019, 20, 3637. [Google Scholar] [CrossRef]
- Israni, B.; Wouters, F.C.; Luck, K.; Seibel, E.; Ahn, S.-J.; Paetz, C.; Reinert, M.; Vogel, H.; Erb, M.; Heckel, D.G. The fall armyworm Spodoptera frugiperda utilizes specific UDP-glycosyltransferases to inactivate maize defensive benzoxazinoids. Front. Physiol. 2020, 11, 604754. [Google Scholar] [CrossRef]
- Lu, K.; Li, Y.; Cheng, Y.; Li, W.; Song, Y.; Zeng, R.; Sun, Z. Activation of the NR2E nuclear receptor HR83 leads to metabolic detoxification-mediated chlorpyrifos resistance in Nilaparvata lugens. Pestic. Biochem. Physiol. 2021, 173, 104800. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Wang, C.; Wang, X.; Li, G.; Liu, Z.; Wang, H.; Guo, X.; Xu, B. Molecular Mechanism of the UDP-Glucuronosyltransferase 2B20-like Gene (AccUGT2B20-like) in Pesticide Resistance of Apis cerana cerana. Front. Genet. 2020, 11, 592595. [Google Scholar] [CrossRef]
- Li, X.; Zhu, B.; Gao, X.; Liang, P. Over-expression of UDP–glycosyltransferase gene UGT2B17 is involved in chlorantraniliprole resistance in Plutella xylostella (L.). Pest Manag. Sci. 2017, 73, 1402–1409. [Google Scholar] [CrossRef]
- Hughes, A.L. Origin of Ecdysosteroid UDP-glycosyltransferases of Baculoviruses through horizontal gene transfer from Lepidoptera. Coevolution 2013, 1, 1–7. [Google Scholar] [CrossRef]
- Shen, G.; Wu, J.; Wang, Y.; Liu, H.; Zhang, H.; Ma, S.; Peng, C.; Lin, Y.; Xia, Q. The expression of ecdysteroid UDP-glucosyltransferase enhances cocoon shell ratio by reducing ecdysteroid titre in last-instar larvae of silkworm, Bombyx mori. Sci. Rep. 2018, 8, 17710. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Sun, R.; Shi, W.; Yan, Y.; Li, H.; Guo, X.; Xu, B. Characterization of a mitochondrial manganese superoxide dismutase gene from Apis cerana cerana and its role in oxidative stress. J. Insect Physiol. 2014, 60, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Pu, J.; Chen, F.; Wang, J.; Han, Z. Multiple ATP-binding cassette transporters are involved in insecticide resistance in the small brown planthopper, Laodelphax striatellus. Insect Mol. Biol. 2017, 26, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Gaddelapati, S.C.; Kalsi, M.; Roy, A.; Palli, S.R. Cap’n’collar C regulates genes responsible for imidacloprid resistance in the Colorado potato beetle, Leptinotarsa decemlineata. Insect Biochem. Mol. Biol. 2018, 99, 54–62. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Heu, C.C.; LeRoy, D.M.; DeGain, B.A.; Yelich, A.J.; Unnithan, G.C.; Wu, Y.; Li, X.; Carrière, Y.; Tabashnik, B.E. Knockout of ABC transporter gene ABCA2 confers resistance to Bt toxin Cry2Ab in Helicoverpa zea. Sci. Rep. 2022, 12, 16706. [Google Scholar] [CrossRef]
- Gao, Q.; Lin, Y.; Wang, X.; Jing, D.; Wang, Z.; He, K.; Bai, S.; Zhang, Y.; Zhang, T. Knockout of ABC transporter ABCG4 gene confers resistance to Cry1 proteins in Ostrinia furnacalis. Toxins 2022, 14, 52. [Google Scholar] [CrossRef]
- Xu, J.; Zheng, J.; Zhang, R.; Wang, H.; Du, J.; Li, J.; Zhou, D.; Sun, Y.; Shen, B. Identification and functional analysis of ABC transporter genes related to deltamethrin resistance in Culex pipiens pallens. Pest Manag. Sci. 2023, 79, 3642–3655. [Google Scholar] [CrossRef]
- Tolasz, R.; Míková, T.; Valeriánová, A.; Voženílek, V. Climate Atlas of Czechia; Czech Hydrometeorological Institute: Prague, Czech Republic, 2007; Volume 256. [Google Scholar]
- Singh, V.V.; Zabihi, K.; Trubin, A.; Jakuš, R.; Cudlín, P.; Korolyova, N.; Blaženec, M. Effect of Diurnal Solar Radiation Regime and Tree Density on Sap Flow of Norway Spruce (Picea abies [L.] Karst.) in Fragmented Stands. 2023. Available online: https://www.researchsquare.com/article/rs-3262723/v1 (accessed on 23 August 2024).
- Everaerts, C.; Grégoire, J.-C.; Merlin, J. The toxicity of Norway spruce monoterpenes to two bark beetle species and their associates. In Mechanisms of Woody Plant Defenses against Insects: Search for Pattern; Springer: Berlin/Heidelberg, Germany, 1988; pp. 335–344. [Google Scholar]
- Boone, C.K.; Aukema, B.H.; Bohlmann, J.; Carroll, A.L.; Raffa, K.F. Efficacy of tree defense physiology varies with bark beetle population density: A basis for positive feedback in eruptive species. Can. J. For. Res. 2011, 41, 1174–1188. [Google Scholar] [CrossRef]
- Faccoli, M.; Schlyter, F. Conifer phenolic resistance markers are bark beetle antifeedant semiochemicals. Agric. For. Entomol. 2007, 9, 237–245. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bioinformatics, B.; Valencia, S. OmicsBox-Bioinformatics made easy. March 2019, 3, 2019. [Google Scholar]
- Tarazona, S.; García-Alcalde, F.; Dopazo, J.; Ferrer, A.; Conesa, A. Differential expression in RNA-seq: A matter of depth. Genome Res. 2011, 21, 2213–2223. [Google Scholar] [CrossRef] [PubMed]
- Tarazona, S.; Furió-Tarí, P.; Turrà, D.; Pietro, A.D.; Nueda, M.J.; Ferrer, A.; Conesa, A. Data quality aware analysis of differential expression in RNA-seq with NOISeq R/Bioc package. Nucleic Acids Res. 2015, 43, e140. [Google Scholar] [CrossRef] [PubMed]
- Bosch-Serra, D.; Rodríguez, M.A.; Avilla, J.; Sarasúa, M.J.; Miarnau, X. Esterase, glutathione S-transferase and NADPH-cytochrome P450 reductase activity evaluation in Cacopsylla pyri L. (Hemiptera: Psyllidae) individual adults. Insects 2021, 12, 329. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Joga, M.R.; Mogilicherla, K.; Smagghe, G.; Roy, A. RNA interference-based forest protection products (FPPs) against wood-boring coleopterans: Hope or hype? Front. Plant Sci. 2021, 12, 733608. [Google Scholar] [CrossRef]
- Mogilicherla, K.; Roy, A. RNAi-chitosan biopesticides for managing forest insect pests: An outlook. Front. For. Glob. Chang. 2023, 6, 1219685. [Google Scholar] [CrossRef]
- OmicsBox. Bioinformatics Made Easy; Version 3.2.2; BioBam Bioinformatics: Valencia, Spain, 3 March 2019; Available online: www.biobam.com/omicsbox (accessed on 22 August 2024).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical | 48 H | %Corrected Beetle Mortality for Different Doses * | ||||||
|---|---|---|---|---|---|---|---|---|
| LC50 (µL/20 mL) | LC70 (µL/20 mL) | % Control Mortality | 1 (µL/20 mL) | 2 (µL/20 mL) | 4 (µL/20 mL) | 8 (µL/20 mL) | 16 (µL/20 mL) | |
| Sabiene | 4.4 | 6.0 | 10.0 | −4.0 | 8.0 | 20.0 | 84.0 | 100.0 |
| α-Pinene | 3.6 | 6.6 | 6.7 | 16.1 | 28.6 | 37.5 | 71.4 | 96.4 |
| Myrcene | 5.3 | 8.1 | 16.7 | 6.0 | 12.0 | 12.0 | 72.0 | 90.0 |
| (R)-(+)-Limonene | 6.2 | 8.4 | 5.0 | 8.8 | 3.5 | 21.1 | 57.9 | 98.3 |
| (S)-(−)-Limonene | 7.3 | 10.6 | 23.0 | 8.7 | 6.5 | 17.4 | 26.1 | 89.1 |
| Sr. No. | Genes | ΔCt Method | BestKeeper | RefFinder | NormFinder | Recommended Genes | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Stability | Rank | Stability | Rank | Stability | Rank | Stability | Rank | |||
| 1 | RPS3-a | 0.52 | 1 | 0.31 | 2 | 1.19 | 1 | 0.144 | 1 | RPS3-a and RPL7 |
| 2 | Arg-K | 0.54 | 2 | 0.38 | 6 | 3.25 | 4 | 0.152 | 2 | |
| 3 | RPL7 | 0.55 | 3 | 0.32 | 3 | 3 | 3 | 0.161 | 4 | |
| 4 | RPS7 | 0.56 | 4 | 0.27 | 1 | 2.99 | 2 | 0.223 | 8 | |
| 5 | UbiQ | 0.56 | 5 | 0.33 | 4 | 4.95 | 6 | 0.196 | 6 | |
| 6 | RPL13-a | 0.58 | 6 | 0.37 | 5 | 5.96 | 7 | 0.221 | 7 | |
| 7 | RPL6 | 0.6 | 7 | 0.37 | 5 | 4.14 | 5 | 0.181 | 5 | |
| 8 | V-ATPase | 0.65 | 8 | 0.51 | 8 | 8.49 | 9 | 0.240 | 10 | |
| 9 | ELF1 | 0.69 | 9 | 0.44 | 7 | 8.49 | 8 | 0.160 | 3 | |
| 10 | Actin | 0.74 | 10 | 0.7 | 11 | 10.72 | 10 | 0.269 | 11 | |
| 11 | GADPH | 0.74 | 11 | 0.6 | 10 | 10.74 | 11 | 0.295 | 12 | |
| 12 | β-Tubulin | 0.77 | 12 | 0.56 | 9 | 11.47 | 12 | 0.238 | 9 | |
| Description | R-Limonene vs. Control | Sabiene vs. Control |
|---|---|---|
| Number of total features | 23,937 | 23,937 |
| Number of filtered features | 12,393 | 12,435 |
| Number of features after filtering | 11,544 | 11,502 |
| DE (probability > 0.9) | 5363 | 6815 |
| Upregulated (M ≥ 1, probability > 0.9) | 2069 | 2817 |
| Downregulated (M ≤ −1, probability > 0.9) | 1961 | 3761 |
| Number of DE detoxification genes | 294 | 426 |
| Sr. No. | Name | Purity | Manufacturer | CAS |
|---|---|---|---|---|
| 1 | α-Pinene | 97% | Thermo Scientific Chemicals, Waltham, MA, USA | 80-56-8 |
| 2 | (S)-(−)-Limonene | 97% | Thermo Scientific Chemicals | 5989-54-8 |
| 3 | (R)-(+)-Limonene | ~90% | Sigma-Aldrich, St. Louis, MO, USA | 5989-27-5 |
| 4 | (1S)-(+)-3-Carene | 99% | Sigma-Aldrich | 498-15-7 |
| 5 | Myrcene | ≥90% | Sigma-Aldrich | 123-35-3 |
| 6 | Sabiene | 75% | Sigma-Aldrich | 3387-41-5 |
| Sr. No. | Primer Name | Transcript ID | Transcript Name | Primer Sequence | Amplicon Length | Tm (°C) | |
|---|---|---|---|---|---|---|---|
| 1 | EST6 | Ityp05867 | Venom carboxylesterase-6 | F | CAACCGAAATGGTGAACTG | 119 | 60 |
| R | ATTCTCAACCACCGTAGAC | ||||||
| 2 | HYEP1-a | Ityp10836 | Juvenile hormone epoxide hydrolase 1 | F | CGGCCTGACTAAACACTTT | 110 | 60 |
| R | AGCCAAACCCTTCAGAATAC | ||||||
| 3 | AK1A1 | Ityp08881 | Alcohol dehydrogenase | F | TCCGAACAACTGCAAAGG | 160 | 60 |
| R | TAGCACCAGGACTTCCTAAA | ||||||
| 4 | GST | Ityp13834 | Glutathione S-transferase | F | CTACTCTGGAAGTGGATGG | 138 | 58, 59 |
| R | AATCCGTGACTGTGTCG | ||||||
| 5 | SDR1-a | Ityp00346 | Farnesol dehydrogenase | F | GGCAATAACCACAAGAGAGG | 139 | 60, 61 |
| R | CGCAAATTTACTGGCTGGA | ||||||
| 6 | ABCGK | Ityp09976 | ABC transporter G family member 20 | F | CCACTGACGCTATTAAGCAC | 143 | 61, 60 |
| R | CAGGTACGCCTTGATTTCTC | ||||||
| 7 | IDLDH-a | Ityp22107 | Ipsdienol dehydrogenase | F | GGACAATAATCGGGACGAAG | 131 | 60 |
| R | GGTTGGTCATGGAGATGATG | ||||||
| 8 | SDR1-b | Ityp13935 | Farnesol dehydrogenase | F | GCGTTAACGGAAACTGTTAG | 153 | 59 |
| R | AGGTCTGTCATTTGCTAGAG | ||||||
| 9 | ST1B1 | Ityp07436 | Sulfotransferase 1B1 | F | AACCACGTTCTGCCATTC | 134 | 60 |
| R | GCTTCAGTGAGGTCTTTCTC | ||||||
| 10 | CP6A2 | Ityp03140 | Cytochrome P450 6a2 | F | TTGAGACATCGGCTACCA | 156 | 60 |
| R | GGCGAAGATAGGTCATTTCC | ||||||
| 11 | UDB17 | Ityp14578 | UDP-glucuronosyltransferase 2B17 | F | GATTCCAACGCCGCTAAA | 139 | 60 |
| R | GACGATGCTTCACTTGACTT | ||||||
| 12 | HYEP1-b | Ityp03551 | Juvenile hormone epoxide hydrolase 1 | F | GAGAGATAGTCCGGT | 127 | 60 |
| R | GTCCAGTAATTGGTC | ||||||
| 13 | IDLDH-b | Ityp14704 | Ipsdienol dehydrogenase | F | CAGACAGTTGGGACCTTTAG | 146 | 60 |
| R | CACGTGGTTTGATCATTCTG | ||||||
| 14 | SDHA | Ityp17457 | Succinate dehydrogenase | F | CGTTCTGGATCTGTTGATGG | 145 | 60 |
| R | GGCTGTGCAGGAGAAATATG | ||||||
| 15 | GSTT1 | Ityp05781 | Glutathione S-transferase 1 | F | GTAGATCAGCGCCTCCATTT | 136 | 60 |
| R | GGGACTGATAAGCTTGCACCACT | ||||||
| 16 | NCPR | Ityp11675 | NADPH-cytochrome P450 reductase | F | GCAAACACTGCGAGAAGA | 153 | 60 |
| R | AAACGTGAGTTCTGGGATTC | ||||||
| 17 | DHB12 | Ityp13083 | 17-beta-hydroxysteroid dehydrogenase 12 | F | ATCAACAACGTCGGGATG | 141 | 60 |
| R | CACCATTCCAGGAAGTACAA | ||||||
| 18 | PA1B2 | Ityp04924 | Platelet-activating factor acetylhydrolase IB subunit beta homolog | F | ACTACCTCGAGGACAGAATC | 136 | 60 |
| R | ACCATCAGGTTGGATAAAGC | ||||||
| 19 | DHGL | Ityp11581 | Glucose dehydrogenase | F | GGCTTTCAGAAGTGGAGAAT | 140 | 60 |
| R | GTTCTGGACGGTGGTATTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naseer, A.; Singh, V.V.; Sellamuthu, G.; Synek, J.; Mogilicherla, K.; Kokoska, L.; Roy, A. Insights into the Detoxification of Spruce Monoterpenes by the Eurasian Spruce Bark Beetle. Int. J. Mol. Sci. 2024, 25, 10209. https://doi.org/10.3390/ijms251810209
Naseer A, Singh VV, Sellamuthu G, Synek J, Mogilicherla K, Kokoska L, Roy A. Insights into the Detoxification of Spruce Monoterpenes by the Eurasian Spruce Bark Beetle. International Journal of Molecular Sciences. 2024; 25(18):10209. https://doi.org/10.3390/ijms251810209
Chicago/Turabian StyleNaseer, Aisha, Vivek Vikram Singh, Gothandapani Sellamuthu, Jiří Synek, Kanakachari Mogilicherla, Ladislav Kokoska, and Amit Roy. 2024. "Insights into the Detoxification of Spruce Monoterpenes by the Eurasian Spruce Bark Beetle" International Journal of Molecular Sciences 25, no. 18: 10209. https://doi.org/10.3390/ijms251810209