
The Serine Acetyltransferase (SAT) Gene Family in Tea Plant (Camellia sinensis): Identification, Classification and Expression Analysis under Salt Stress


Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification of SAT Genes in Tea Plant
2.2. Analysis of Multiple Sequence Alignment and Phylogenetic Relationships
2.3. Analysis of the Conserved Structure and Genetic Architecture of CssSATs
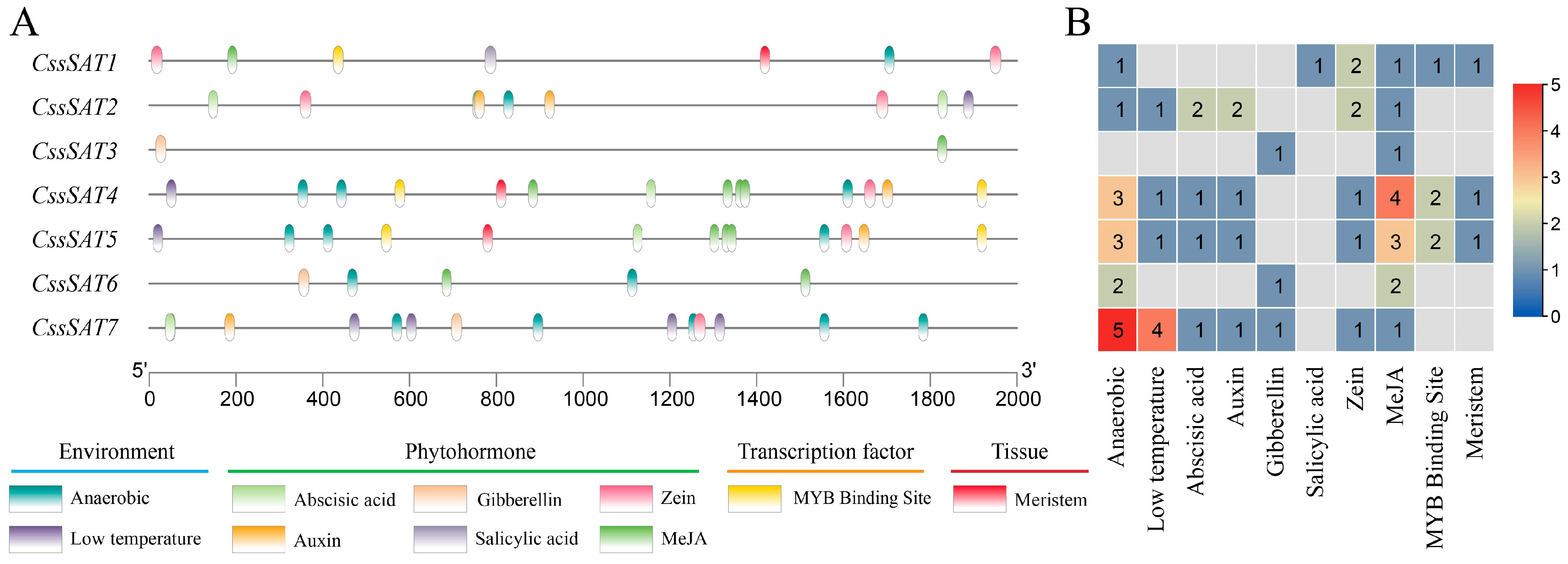
2.4. Analysis of Cis-Acting Elements in the Promoter Region of the CssSATs Genes
2.5. Analysis of Expression Profiles of the CssSATs Genes under Different Stress Conditions
2.6. Analysis of the Expression Profile of CssSATs Genes in Diverse Tissues of Tea Plant
2.7. Analysis of qRT-PCR Results for the CssSATs Genes
2.8. Analysis of Prokaryotic Expression of the CssSATs Genes
3. Discussion
4. Materials and Methods
4.1. Experimental Materials
4.2. Screening and Sequence Analysis of CssSATs Genes
4.3. Alignment and Phylogenetic Analysis of CssSATs Sequences
4.4. Analysis of the Structure and Conserved Motifs of CssSATs Genes
4.5. Analysis of Cis-Acting Elements within the Promoter Region of CssSATs Genes
4.6. Analysis of the Transcriptional Expression of CssSATs Genes
4.7. Analysis of the Prokaryotic Expression of CssSATs Genes and Salt Tolerance of the Recombinant Bacterial Strain
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, C.S.; Lambert, J.D.; Sang, S. Antioxidative and anti-carcinogenic activities of tea polyphenols. Arch. Toxicol. 2009, 83, 11–21. [Google Scholar] [CrossRef]
- Waltner-Law, M.E.; Wang, X.L.; Law, B.K.; Hall, R.K.; Nawano, M.; Granner, D.K. Epigallocatechin gallate, a constituent of green tea, represses hepatic glucose production. J. Biol. Chem. 2002, 277, 34933–34940. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Gwee, X.; Kua, E.H.; Ng, T.P. Cognitive function and tea consumption in community dwelling older Chinese in Singapore. J. Nutr. Health Aging 2010, 14, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.-H.; Li, N.; Zhu, H.-T.; Wang, D.; Yang, C.-R.; Zhang, Y.-J. Plant resources, chemical constituents, and bioactivities of tea plants from the genus Camellia section Thea. J. Agric. Food Chem. 2019, 67, 5318–5349. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.M.; Sousa, C. A review on the biological activity of Camellia species. Molecules 2021, 26, 2178. [Google Scholar] [CrossRef]
- Sun, Y.; Zhou, J.; Guo, J. Advances in the knowledge of adaptive mechanisms mediating abiotic stress responses in Camellia sinensis. Front. Biosci.-Landmark 2021, 26, 1714–1722. [Google Scholar] [CrossRef]
- Zhu, J.-K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef]
- Romero, L.C.; Aroca, M.Á.; Laureano, A.M.; Moreno, I.; García, I.; Gotor, C. Cysteine and cysteine-related signaling pathways in Arabidopsis thaliana. Mol. Plant 2014, 7, 264–276. [Google Scholar] [CrossRef]
- Noji, M.; Inoue, K.; Kimura, N.; Gouda, A.; Saito, K. Isoform-dependent differences in feedback regulation and subcellular localization of serine acetyltransferase involved in cysteine biosynthesis from Arabidopsis thaliana. J. Biol. Chem. 1998, 273, 32739–32745. [Google Scholar] [CrossRef]
- Chen, Y.; Luo, C.; Liu, Y.; Xiao, Z.; Lu, Y.; Hou, Z.; Xiao, Q.; Zhou, Y.; Tang, Q. Gene identification, expression analysis, and molecular docking of SAT and OASTL in the metabolic pathway of selenium in Cardamine hupingshanensis. Plant Cell Rep. 2024, 43, 148. [Google Scholar] [CrossRef]
- Yeon, J.Y.; Yoo, S.J.; Takagi, H.; Kang, H.A. A novel mitochondrial serine O-acetyltransferase, OpSAT1, plays a critical role in sulfur metabolism in the thermotolerant methylotrophic yeast Ogataea parapolymorpha. Sci. Rep. 2018, 8, 2377. [Google Scholar] [CrossRef] [PubMed]
- Tavares, S.; Wirtz, M.; Beier, M.P.; Bogs, J.; Hell, R.; Amâncio, S. Characterization of the serine acetyltransferase gene family of Vitis vinifera uncovers differences in regulation of OAS synthesis in woody plants. Front. Plant Sci. 2015, 6, 74. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Hubberten, H.M.; Saito, K.; Hoefgen, R. General regulatory patterns of plant mineral nutrient depletion as revealed by serat quadruple mutants disturbed in cysteine synthesis. Mol. Plant 2010, 3, 438–466. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Tohge, T.; Fernie, A.R.; Hoefgen, R. The effect of single and multiple SERAT mutants on serine and sulfur metabolism. Front. Plant Sci. 2018, 9, 702. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Mochida, K.; Kato, T.; Tabata, S.; Yoshimoto, N.; Noji, M.; Saito, K. Comparative genomics and reverse genetics analysis reveal indispensable functions of the serine acetyltransferase gene family in Arabidopsis. Plant Cell 2008, 20, 2484–2496. [Google Scholar] [CrossRef] [PubMed]
- Na, G.; Salt, D.E. Differential regulation of serine acetyltransferase is involved in nickel hyperaccumulation in Thlaspi goesingense. J. Biol. Chem. 2011, 286, 40423–40432. [Google Scholar] [CrossRef]
- Kawashima, C.G.; Berkowitz, O.; Hell, R.; Noji, M.; Saito, K. Characterization and expression analysis of a serine acetyltransferase gene family involved in a key step of the sulfur assimilation pathway in Arabidopsis. Plant Physiol. 2005, 137, 220–230. [Google Scholar] [CrossRef]
- Freeman, J.L.; Salt, D.E. The metal tolerance profile of Thlaspi goesingense is mimicked in Arabidopsis thaliana heterologously expressing serine acetyl-transferase. BMC Plant Biol. 2007, 7, 63. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Dominguez, J.R.; He, Z.; Lima, A.; Ting, J.; Buchanan, B.B.; Luan, S. A cyclophilin links redox and light signals to cysteine biosynthesis and stress responses in chloroplasts. Proc. Natl. Acad. Sci. USA 2008, 105, 16386–16391. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef]
- Wirtz, M.; Hell, R. Dominant-negative modification reveals the regulatory function of the multimeric cysteine synthase protein complex in transgenic tobacco. Plant Cell 2007, 19, 625–639. [Google Scholar] [CrossRef]
- Xiang, X.; Wu, Y.; Planta, J.; Messing, J.; Leustek, T. Overexpression of serine acetyltransferase in maize leaves increases seed-specific methionine-rich zeins. Plant Biotechnol. J. 2018, 16, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.-H.; Tong, W.; Wu, Q.; Wei, S.; Zhao, J.; Zhang, Z.-Z.; Wei, C.-L.; Wan, X.-C. Tea plant genomics: Achievements, challenges and perspectives. Hortic. Res. 2020, 7, 7. [Google Scholar] [CrossRef]
- Wirtz, M.; Droux, M. Synthesis of the sulfur amino acids: Cysteine and methionine. Photosynth. Res. 2005, 86, 345–362. [Google Scholar] [CrossRef]
- Howarth, J.R.; Domínguez, J.R.; Gutiérrez, G.; Wray, J.L.; Romero, L.C.; Gotor, C. The serine acetyltransferase gene family in Arabidopsis thaliana and the regulation of its expression by cadmium. Plant Mol. Biol. 2003, 51, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Li, M.; Guo, T.; Lu, J.; Xie, Y.; Hao, Y.; Wang, L.; Zhao, D.; Zhang, L.; Liu, Z.; et al. Functional characterization of the serine acetyltransferase family genes uncovers the diversification and conservation of cysteine biosynthesis in tomato. Front. Plant Sci. 2022, 13, 913856. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, C.; Yin, W.; Wei, W.; Wang, Y.; Sa, W.; Liang, J. Single-molecule real-time sequencing of the full-length transcriptome of purple garlic (Allium sativum L. cv. Leduzipi) and identification of serine O-acetyltransferase family proteins involved in cysteine biosynthesis. J. Sci. Food Agric. 2022, 102, 2864–2873. [Google Scholar] [CrossRef]
- Chen, J.D.; Zheng, C.; Ma, J.Q.; Jiang, C.K.; Ercisli, S.; Yao, M.Z.; Chen, L. The chromosome-scale genome reveals the evolution and diversification after the recent tetraploidization event in tea plant. Hortic. Res. 2020, 7, 63. [Google Scholar] [CrossRef]
- Ma, H.; Song, Y.; Zhang, Y.; Guo, H.; Lv, G.; Chen, H.; Liu, J.; Liu, X.; An, Z.; Wang, L.; et al. Critical sites of serine acetyltransferase in Lathyrus sativus L. affecting its enzymatic activities. J. Agric. Food Chem. 2023, 71, 7858–7865. [Google Scholar] [CrossRef] [PubMed]
- Dharavath, S.; Kumari, K.; Kumar, S.; Gourinath, S. Structural and functional studies of serine acetyltransferase isoform from Entamoeba histolytica reveals novel role of the c-terminal tail in loss of regulation from feedback inhibition. Int. J. Biol. Macromol. 2022, 217, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Francois, J.A.; Kumaran, S.; Jez, J.M. Structural basis for interaction of O-acetylserine sulfhydrylase and serine acetyltransferase in the Arabidopsis cysteine synthase complex. Plant Cell 2006, 18, 3647–3655. [Google Scholar] [CrossRef]
- Campanini, B.; Speroni, F.; Salsi, E.; Cook, P.F.; Roderick, S.L.; Huang, B.; Bettati, S.; Mozzarelli, A. Interaction of serine acetyltransferase with O-acetylserine sulfhydrylase active site: Evidence from fluorescence spectroscopy. Protein Sci. 2005, 14, 2115–2124. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, M.; Hell, R. Production of cysteine for bacterial and plant biotechnology: Application of cysteine feedback-insensitive isoforms of serine acetyltransferase. Amino Acids 2003, 24, 195–203. [Google Scholar] [CrossRef]
- Genisel, M.; Erdal, S.; Kizilkaya, M. The mitigating effect of cysteine on growth inhibition in salt-stressed barley seeds is related to its own reducing capacity rather than its effects on antioxidant system. Plant Growth Regul. 2015, 75, 187–197. [Google Scholar] [CrossRef]
- Kurt, F.; Filiz, E.; Aydın, A. Genome-wide identification of serine acetyltransferase (SAT) gene family in rice (Oryza sativa) and their expressions under salt stress. Mol. Biol. Rep. 2021, 48, 6277–6290. [Google Scholar] [CrossRef]
- Wang, J.; Song, L.; Gong, X.; Xu, J.; Li, M. Functions of jasmonic acid in plant regulation and response to abiotic stress. Int. J. Mol. Sci. 2020, 21, 1446. [Google Scholar] [CrossRef]
- Jing, Y.; Gui, D.; Chun, L.; Lin, L.; Guang, H.; Ya, Z.; Chang, W. The crosstalks between jasmonic acid and other plant hormone signaling highlight the involvement of jasmonic acid as a core component in plant response to biotic and abiotic stresses. Front. Plant Sci. 2019, 10, 1349. [Google Scholar] [CrossRef]
- Shahzad, A.N.; Pitann, B.; Ali, H.; Qayyum, M.F.; Fatima, A.; Bakhat, H.F. Maize genotypes differing in salt resistance vary in jasmonic acid accumulation during the first phase of salt stress. J. Agron. Crop Sci. 2015, 201, 443–451. [Google Scholar] [CrossRef]
- Zong, Q.; Jun, G.; Ai, Z.; Liang, Z.; Man, Z. Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress. Ecotoxicol. Environ. Saf. 2014, 104, 202–208. [Google Scholar] [CrossRef]
- Lv, Z.; Zhang, C.; Shao, C.; Liu, B.; Liu, E.; Yuan, D.; Zhou, Y.; Shen, C. Research progress on the response of tea catechins to drought stress. J. Sci. Food Agric. 2021, 101, 5305–5313. [Google Scholar] [CrossRef]
- Zuo, H.; Chen, J.; Lv, Z.; Shao, C.; Chen, Z.; Zhou, Y.; Shen, C. Tea-derived polyphenols enhance drought resistance of tea plants (Camellia sinensis) by alleviating jasmonate–isoleucine pathway and flavonoid metabolism flow. Int. J. Mol. Sci. 2024, 25, 3817. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Malagoli, M.; Wirtz, M.; Hell, R. Drought stress in maize causes differential acclimation responses of glutathione and sulfur metabolism in leaves and roots. BMC Plant Biol. 2016, 16, 247. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.J.; Wang, Z.; Zhao, Q.; Mao, J.L.; Speiser, A.; Wirtz, M.; Hell, R.; Zhu, J.K.; Xiang, C.B. Sulfate availability affects ABA levels and germination response to ABA and salt stress in Arabidopsis thaliana. Plant J. 2014, 77, 604–615. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, Y.; Zhai, F.; Zhang, J.; Zhang, F.; Yuan, X.; Xie, Y. Hydrogen sulfide promotes rice drought tolerance via reestablishing redox homeostasis and activation of ABA biosynthesis and signaling. Plant Physiol. Biochem. 2020, 155, 213–220. [Google Scholar] [CrossRef]
- Velayutham, M.; Ojha, B.; Issac, P.K.; Lite, C.; Guru, A.; Pasupuleti, M.; Arasu, M.V.; Al-Dhabi, N.A.; Arockiaraj, J. NV14 from serine O-acetyltransferase of cyanobacteria influences the antioxidant enzymes in vitro cells, gene expression against H2O2 and other responses in vivo zebrafish larval model. Cell Biol. Int. 2021, 45, 2331–2346. [Google Scholar] [CrossRef]
- Rajab, H.; Khan, M.S.; Wirtz, M.; Malagoli, M.; Qahar, F.; Hell, R. Sulfur metabolic engineering enhances cadmium stress tolerance and root to shoot iron translocation in Brassica napus L. Plant Physiol. Biochem. 2020, 152, 32–43. [Google Scholar] [CrossRef]
- Qahar, F.; Khan, M.S. Evaluating the performance of genetically engineered serine acetyltransferase 4 (NtSAT4) overexpression Brassica napus L. lines under xenobiotics exposure. Sarhad J. Agric. 2023, 39, 765–772. [Google Scholar] [CrossRef]
- Sadak, M.S.; Abd El-Hameid, A.R.; Zaki, F.S.A.; Dawood, M.G.; El-Awadi, M.E. Physiological and biochemical responses of soybean (Glycine max L.) to cysteine application under sea salt stress. Bull. Natl. Res. Cent. 2019, 44, 5692–5699. [Google Scholar] [CrossRef]
- Gao, Q.; Tong, W.; Li, F.; Wang, Y.; Wu, Q.; Wan, X.; Xia, E. TPIA2: An updated tea plant information archive for Camellia genomics. Nucleic Acids Res. 2024, 52, D1661–D1667. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; et al. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Yang, H.; Wang, S.; Zhao, J.; Liu, C.; Gao, L.; Xia, E.; Lu, Y.; Tai, Y.; She, G.; et al. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA 2018, 115, E4151–E4158. [Google Scholar] [CrossRef]
- Zhang, Q.; Cai, M.; Yu, X.; Wang, L.; Guo, C.; Ming, R.; Zhang, J. Transcriptome dynamics of Camellia sinensis in response to continuous salinity and drought stress. Tree Genet. Genomes 2017, 13, 78. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Ban, Q.; Zhu, X.; Jiang, C.; Wei, C.; Bennetzen, J.L. Comparative transcriptomic analysis reveals gene expression associated with cold adaptation in the tea plant Camellia sinensis. BMC Genom. 2019, 20, 624. [Google Scholar] [CrossRef]
- Shi, J.; Ma, C.; Qi, D.; Lv, H.; Yang, T.; Peng, Q.; Chen, Z.; Lin, Z. Transcriptional responses and flavor volatiles biosynthesis in methyl jasmonate-treated tea leaves. BMC Plant Biol. 2015, 15, 233. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chromosomal Localization | AA 1 | MW 2 (kD) | pI 3 | Subcellular Localization |
|---|---|---|---|---|---|---|
| CssSAT1 | CSS0013877.1 | Chr2: 48272763-48290939 | 354 | 38.42 | 5.23 | Cytoplasm |
| CssSAT2 | CSS0023998.1 | Chr6: 45069503-45071954 | 270 | 28.69 | 6.25 | Cytoplasm |
| CssSAT3 | CSS0003072.1 | Chr7: 11430067-11431056 | 300 | 32.90 | 6.85 | Chloroplast |
| CssSAT4 | CSS0044287.1 | Chr8: 2394885-2396035 | 336 | 36.39 | 8.07 | Cytoplasm |
| CssSAT5 | CSS0029099.1 | Chr8: 2527843-2528854 | 322 | 34.90 | 8.55 | Cytoplasm |
| CssSAT6 | CSS0026852.1 | Chr14: 118914206-118915594 | 294 | 31.28 | 6.70 | Cytoplasm |
| CssSAT7 | CSS0009950.1 | Chr15: 15007141-15008025 | 283 | 29.98 | 6.29 | Cytoplasm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Liu, D.; Jiao, X.; Wu, Q.; Wang, W. The Serine Acetyltransferase (SAT) Gene Family in Tea Plant (Camellia sinensis): Identification, Classification and Expression Analysis under Salt Stress. Int. J. Mol. Sci. 2024, 25, 9794. https://doi.org/10.3390/ijms25189794
Wang L, Liu D, Jiao X, Wu Q, Wang W. The Serine Acetyltransferase (SAT) Gene Family in Tea Plant (Camellia sinensis): Identification, Classification and Expression Analysis under Salt Stress. International Journal of Molecular Sciences. 2024; 25(18):9794. https://doi.org/10.3390/ijms25189794
Chicago/Turabian StyleWang, Leigang, Dandan Liu, Xiaoyu Jiao, Qiong Wu, and Wenjie Wang. 2024. "The Serine Acetyltransferase (SAT) Gene Family in Tea Plant (Camellia sinensis): Identification, Classification and Expression Analysis under Salt Stress" International Journal of Molecular Sciences 25, no. 18: 9794. https://doi.org/10.3390/ijms25189794