Abstract
Pancreatic cancer presents formidable challenges due to rapid progression and resistance to conventional treatments. Oncolytic viruses (OVs) selectively infect cancer cells and cause cancer cells to lyse, releasing molecules that can be identified by the host’s immune system. Moreover, OV can carry immune-stimulatory payloads such as interleukin-12, which when delivered locally can enhance immune system-mediated tumor killing. OVs are very well tolerated by cancer patients due to their ability to selectively target tumors without affecting surrounding normal tissues. OVs have recently been combined with other therapies, including chemotherapy and immunotherapy, to improve clinical outcomes. Several OVs including adenovirus, herpes simplex viruses (HSVs), vaccinia virus, parvovirus, reovirus, and measles virus have been evaluated in preclinical and clinical settings for the treatment of pancreatic cancer. We evaluated the safety and tolerability of a replication-competent oncolytic adenoviral vector carrying two suicide genes (thymidine kinase, TK; and cytosine deaminase, CD) and human interleukin-12 (hIL12) in metastatic pancreatic cancer patients in a phase 1 trial. This vector was found to be safe and well-tolerated at the highest doses tested without causing any significant adverse events (SAEs). Moreover, long-term follow-up studies indicated an increase in the overall survival (OS) in subjects receiving the highest dose of the OV. Our encouraging long-term survival data provide hope for patients with advanced pancreatic cancer, a disease that has not seen a meaningful increase in OS in the last five decades. In this review article, we highlight several preclinical and clinical studies and discuss future directions for optimizing OV therapy in pancreatic cancer. We envision OV-based gene therapy to be a game changer in the near future with the advent of newer generation OVs that have higher specificity and selectivity combined with personalized treatment plans developed under AI guidance.
1. Introduction
Pancreatic cancer remains one of the most lethal malignancies, with a five-year survival rate of 13% [1]. The primary reasons for this dismal prognosis include late diagnosis, aggressive tumor biology, and limited treatment options [2]. Traditional therapies such as surgery, chemotherapy, and radiation often fall short due to the tumor’s resistance mechanisms and the complex tumor microenvironment (TME) [3]. While complete surgical resection can significantly extend patient survival and is considered the only curative approach, only about 20% of patients are eligible for surgery, mainly due to diagnosis at an advanced stage [4,5,6]. The standard-of-care for metastatic pancreatic adenocarcinoma is continuous chemotherapy with FOLFIRINOX or gemcitabine and nanoparticle albumin-bound paclitaxel based on the PRODIGE-4/ACCORD-11 [7] and MPACT trials [8]. While initial stages of chemotherapy often lead to patient improvement, most eventually develop drug resistance, which severely impacts their prognosis [9,10,11,12]. Consequently, there is an urgent need for innovative therapeutic strategies that can overcome these barriers and improve patient outcomes.
The potential of viruses as anticancer agents was first recognized in the early 20th century when physicians observed tumor regression in some cancer patients following viral infections. These initial findings sparked the idea of using viruses to fight cancer. By the 1960s, research into oncolytic viruses gained momentum, with scientists investigating naturally occurring viruses for their cancer-killing properties. However, early studies faced challenges such as uncontrolled infections and immune responses that limited viral efficacy. The field was revolutionized in the late 20th century with advances in genetic engineering, which led to the development of viruses that selectively target cancer cells while minimizing their pathogenicity [13,14]. In recent years, immunotherapy has revolutionized cancer treatment, offering new hope where conventional therapies have failed [15]. Among emerging modalities, OVs have garnered significant attention for their dual ability to selectively infect and lyse cancer cells while simultaneously stimulating robust antitumor immune responses to destroy neighboring cancer cells [16]. OVs are either naturally occurring oncotropic viruses or genetically engineered to selectively infect and destroy cancer cells while sparing normal tissues [17]. OVs exploit the unique characteristics of cancer cells, such as abnormal responses to stress, impaired antiviral responses, and specific surface receptors, to selectively infect them. For example, reovirus targets cells with Ras mutations, while vesicular stomatitis virus (VSV) and Newcastle disease virus preferentially infect cells with interferon response-related mutations [16,17]. Certain viruses utilize specific cell surface receptors; poliovirus targets CD155, while measles virus targets CD46 or CD150 [18]. Other viruses like herpes simplex virus type 1 (HSV-1) take advantage of malignancy-driven cellular changes, including alterations in the extracellular matrix for infecting cancer cells [19].
1.1. Oncolytic Viruses and Mechanism of Action
OVs employ a multifaceted mechanism of action that includes both direct tumor cell destruction (oncolysis) and the stimulation of the immune system. Upon infection, the virus replicates within the cancer cell and hijacks the cell’s protein production machinery leading to cell lysis and the release of new viral particles that can infect adjacent cancer cells, further propagating the oncolytic effect [20]. The lysis of tumor cells releases tumor antigens, damage-associated molecular patterns (DAMPs), and pathogen-associated molecular patterns (PAMPs). These components trigger host immune responses and activate antitumor immunity, which further activates T cells and other immune responses against tumors [21,22]. The presence of viral particles and tumor antigens thus stimulate both the innate and adaptive immune responses and transform the immunosuppressive TME to a more immunologically responsive state [23,24]. The modulation of the TME by OVs is further discussed in detail later in this review.
Additionally, OVs with natural infection abilities can be genetically modified to improve their immunogenicity and effectiveness against cancer cells [18]. This enhancement involves knocking out certain genes to reduce infection of normal cells, inserting new genes to boost oncolytic activity, and transferring foreign genes to improve immune responses [25]. Tumor-specific promoters ensure the virus replicates selectively in cancer cells. Engineered OVs can express cytokines such as TNF, GM-CSF, IL-7, IL-12, and IFN-β, which enhance cell-lysing capabilities and stimulate antitumor immunity [26,27,28,29]. These modifications improve safety, tumor specificity, and potency while reducing pathogenicity. Modified OVs evaluated in clinical trials include adenoviruses, vesicular stomatitis, vaccinia, measles, and herpes simplex viruses [17].
Furthermore, OVs can be combined with other cancer therapies such as chemotherapy, radiation therapy, or immunotherapy to enhance their effectiveness [30]. For example, combining OVs with agents that target the dense stromal environment of tumors like pancreatic tumors can improve viral delivery and penetration into the tumor mass [31]. Overall, OVs may offer several advantages over conventional immunotherapies, including accurate targeting, high efficacy in killing cancer cells, and minimal side effects [32,33,34]. This makes OVs particularly suited for overcoming significant challenges posed by pancreatic cancer.
1.2. Types of Oncolytic Viruses
OVs are classified based on their genetic material into DNA and RNA viruses. OVs with DNA as genetic material include adenoviruses, herpes simplex virus (HSV), parvoviruses, and poxviruses (e.g., vaccinia virus, myxoma virus). OVs based on RNA viruses include coxsackievirus, maraba virus, measles virus (MV), Newcastle disease virus (NDV), poliovirus, reovirus, retroviruses, Seneca Valley virus (SVV), Semliki Forest virus (SFV), vesicular stomatitis virus (VSV), and sindbis virus (SBV). Each virus type has unique characteristics, advantages, and limitations, making them suitable for different cancers and treatment strategies [34].
1.3. Oncolytic Viruses and the Tumor Microenvironment
Pancreatic cancer is characterized by a challenging tumor microenvironment and dense, fibrotic tissue known as desmoplasia [35]. This desmoplastic stroma, composed of both cellular and noncellular components, forms a thick extracellular matrix (ECM) barrier around the tumor [35] that hinders the effectiveness of treatments and promotes tumor growth and invasion [36]. Moreover, pancreatic cancer is thought to be an “immune cold” tumor, with an immunosuppressive TME [37] that helps the tumor evade the body’s natural defenses and supports cancer progression [38].
OVs can alter the TME to boost immune activation and destroy cancer cells in several ways [39]. First, they exploit cancer cells’ dysfunctional interferon (IFN) pathway for easy infection [40]. OVs induce immunogenic cell death (ICD), releasing molecules like calreticulin, ATP, and HMGB1 [41], which activate dendritic cells (DCs) and T lymphocytes, overcoming the tumor’s immune evasion [21]. This local immune activation often leads to a long-lasting anticancer response, even in advanced stages [42].
Additionally, OVs influence tumor vasculature, tumor-associated fibroblasts, ECM, and various other components [43]. Oncolytic viruses (OVs) can target both nascent and established blood vessels in tumors without harming normal vasculature. They can directly infect and lyse tumor endothelial cells, induce immune responses that reduce tumor perfusion, and express viral proteins with antiangiogenic properties [44]. Genetically engineered OVs can additionally deliver antivascular agents to further enhance these effects [45].
Cancer-associated fibroblasts (CAFs) and the ECM are crucial components of the TME that support tumor growth and metastasis [38]. OVs can disrupt the interaction between cancer cells and CAFs leading to a less supportive environment for tumor growth. Ilkow et al. demonstrated that vesicular stomatitis virus (VSV)-based therapeutics are enhanced through interactions between CAFs and cancer cells [46]. Transforming growth factor-beta 1 (TGF-β1) secreted by tumor cells promotes OV infection in CAFs, while high levels of fibroblast growth factor 2 (FGF2) make tumor cells more susceptible to viral infection [46]. Additionally, oncolytic adenoviruses (OAds) have been shown to target both glioblastoma cells and glioblastoma-associated stromal FAP+ cells, effectively disrupting tumor and stromal cell interactions [45].
To enhance their spread within tumors, OVs use strategies like expressing ECM-degrading proteins such as relaxin and decorin [47,48,49]. Hyaluronidase-armed adenoviruses such as VCN-01 demonstrated promise in overcoming ECM barriers in clinical trials, including in pancreatic cancer (NCT02045589, NCT02045602) [50,51]. Additionally, OVs can convert unresponsive “cold” tumors into more responsive “hot” tumors, thus enhancing the effectiveness of immunotherapy [23,24,52]. Thus, OVs not only directly kill cancer cells but also reshape the TME to enhance immune-mediated destruction of tumors. By inducing inflammation, normalizing vasculature, disrupting stromal components, targeting the ECM and the crosstalk between the TME components, OVs create an environment hostile to cancer cells and improve clinical outcome. The mechanisms of OV activity and tumor microenvironment remodeling in pancreatic cancer are shown in Figure 1.
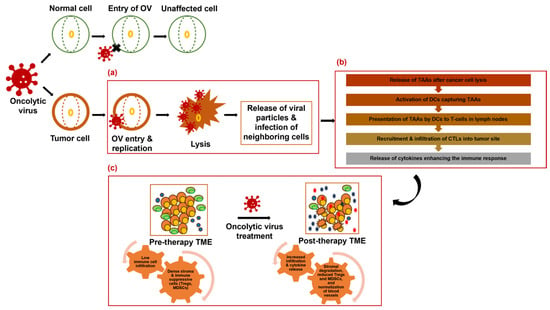
Figure 1.
Mechanism of oncolytic virus activity and tumor microenvironment remodeling in pancreatic cancer. (a) OVs specifically infect and lyse cancer cells, releasing viral particles and infecting neighboring cells. (b) Release of tumor-associated antigens (TAAs) after cancer cell lysis leads to an immune response characterized by dendritic cell activation, T-cell priming, and recruitment of cytotoxic T lymphocytes (CTLs) to the tumor site. (c) OVs also remodel the tumor microenvironment (TME) by degrading stromal components, reducing immunosuppressive cells, and promoting immune cell infiltration. These combined effects result in tumor shrinkage and potentially synergize with other therapeutic modalities.
1.4. Development of OVs
OVs as a cancer treatment strategy have evolved significantly over the years. The concept of using viruses for cancer therapy dates to the late 19th century [53], but initial trials with wild-type RNA viruses and adenoviruses in the mid-20th century had limited success [54]. Interest in OVs as anticancer agents increased in the 1990s with the demonstration of genetically engineered herpes simplex virus (HSV) [55]. Since then, a diverse range of DNA and RNA viruses tailored for tumor specificity and safety have entered clinical trials [54]. Key breakthroughs include FDA and EMA approval of Talimogene Laherparepvec (T-Vec; Herpes simplex virus) in 2015 for advanced melanoma in the USA and Europe [56]. Other country-specific approved OVs include Rigvir (picornavirus, treatment of melanoma, Latvia), H101 (adenovirus, treatment of head and neck cancer, China), and DELYTACT (Herpes simplex virus, treatment of brain cancers such as glioblastoma, Japan) [57].
Today, OVs represent a promising frontier in cancer therapy, with ongoing research exploring their efficacy across various cancer types and refining treatment approaches. Numerous preclinical and clinical studies have demonstrated the safety and efficacy of OVs in various cancer models, including glioblastoma, breast, prostate, and pancreatic cancers [17,58,59,60].
2. Oncolytic Viruses for Pancreatic Cancer
2.1. Preclinical Studies of Oncolytic Viruses in Pancreatic Cancer
Precise tumor targeting is crucial for the effective systemic delivery of OVs. Several different viruses including adenovirus, HSV, vaccinia virus, parvovirus, reovirus, and measles virus have been tested and validated in preclinical pancreatic cancer models. Some key studies are discussed below.
Delta-24-RGD (DNX-2401) is an OV designed to selectively replicate in tumor cells with p16/RB/E2F pathway abnormalities. It is an OAd with a 24-base pair deletion in the E1A region and an RGD-4C modification in the virus fiber which enhances its ability to infect cancer cells independently of coxsackievirus and adenovirus receptor expression. DNX-2401 significantly reduced tumor growth and enhanced anticancer activity in pancreatic cancer models, particularly when combined with the phosphatidylserine-targeting antibody 1N11, suggesting synergistic anticancer immune responses [61]. Similarly, a neurotensin peptide-conjugated polyethylene glycol (PEG-NT)-coated OAd was engineered to express both decorin (DCN) and a soluble Wnt decoy peptide sLRP6E1E2 (oAd/DCN/LRP-PEG-NT) specifically targeting pancreatic cancer [62]. This dual-function approach aims to degrade the extracellular matrix (ECM) and disrupt Wnt signaling, thus enhancing the therapeutic efficacy in pancreatic tumors. OAd/DCN/LRP-PEG-NT effectively targeted neurotensin receptor 1 (NTR)-overexpressing pancreatic cancer cells. It increased cancer cell killing, improved transduction efficiency, reduced immune responses, prolonged blood retention, and significantly suppressed tumor growth in vivo [62]. Na et al. explored a novel approach to improve systemic delivery of OAds using human bone marrow-derived mesenchymal stromal cells (hMSCs) as carriers. hMSCs were chosen due to their natural ability to migrate to tumors and their low immunogenicity, which protects the virus from the immune system. OAd was complexed with relaxin to help break down the dense ECM, allowing better penetration of the virus, and biodegradable polymer (PCDP) to boost the internalization of the virus into hMSCs [63]. The oAd/RLX-PCDP complex demonstrated enhanced internalization, superior viral replication and release within tumors, and relaxin expression, resulting in a stronger antitumor effect compared to naked oAd/RLX or oAd/RLX-treated hMSCs. The study suggests that hMSCs, along with relaxin-expressing oAd, could effectively overcome the barriers of the tumor microenvironment, offering a promising strategy for improving pancreatic cancer treatment [63]. Another study targeting the desmoplastic TME of pancreatic cancer investigated the effect of ECM-degrading relaxin expressing OAd (YDC002) combined with gemcitabine in chemo-resistant pancreatic cancer. YDC002 degraded ECM, overcame ECM-induced chemoresistance, and enhanced gemcitabine-mediated cytotoxicity and oncolytic effects [47]. Brugada-Vilà et al. aimed to enhance the systemic delivery and efficacy of oncolytic adenoviruses by coating them with PEGylated oligopeptide-modified poly(β-amino ester)s (OM-pBAEs). This formulation improved transduction, evaded neutralizing antibodies, reduced liver sequestration, and enhanced therapeutic potential in PDAC mouse models compared to noncoated viruses [64].
Similarly, IFN-α expressed from oncolytic adenoviruses synergistically increased the effectiveness of radiation and chemotherapy (5-FU, gemcitabine, and cisplatin) for in vitro and in vivo pancreatic cancer models. This study highlighted that combining IFN-expressing oncolytic adenoviruses with chemoradiation offered a promising innovative approach for pancreatic cancer patients, particularly those unable to tolerate standard chemotherapy [65]. Watanabe et al. demonstrated that combining mesothelin-redirected chimeric antigen receptor T cells (meso-CAR T cells) with an oncolytic adenovirus expressing TNF-α and IL-2 (OAd-TNFa-IL2) significantly enhanced antitumor efficacy in pancreatic ductal adenocarcinoma (PDAC) models. This approach increased tumor-infiltrating lymphocytes (TILs), enhanced T-cell function, prevented tumor metastasis, and induced tumor regression [66]. OAds are designed to target cancer cells, but genetic variability in tumors, like abnormal miRNA expression in pancreatic cancer, can hinder their replication. Raimondi et al. found that inhibiting the overexpressed miR-222 in cancer cells sensitized them to viral oncolysis. A novel OAd engineered to reduce miR-222 (AdNuPARmE1A-miR222-S) enhanced viral replication and cytotoxicity and effectively controlled tumor growth in vivo. The improved antitumor effects resulted from miR-222 inhibition and restoration of target [67].
To address the challenges faced by traditional Ad5-based OAds in entry into cancer cells due to low levels of coxsackievirus and adenovirus receptor (CAR), Doerner et al. developed novel (OAds) derived from different serotypes [68]. These new OAds include a tumor-selective mutation to allow the viruses to selectively replicate in tumor cells and express the RNAi inhibitor P19 to enhance viral replication and oncolytic activity. OAds based on Ad1, Ad2, and Ad6 were more effective at lysing cancer cells than the traditional Ad5-based OAd [68]. Hashimoto et al. developed a telomerase-specific OAd armed with the p53 gene (OBP-702) and investigated its potential to promote long-term antitumor immunity. OBP-702 increased effector memory precursor cells and led to tissue-resident and effector memory T cells in murine pancreatic tumors. OBP-702 in combination with gemcitabine and nab-paclitaxel (GN) maintained memory T-cell activation and showed significant antitumor effects. In a neoadjuvant model, GN with OBP-702 provided long-term antitumor effects post-tumor resection, highlighting OBP-702’s potential as a long-term immunostimulant in pancreatic cancer [69]. Ge et al. [70] developed an OAd that expressed tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) and second mitochondria-derived activator of caspase (Smac) (ZD55-TRAIL-IETD-Smac) and evaluated its activity in pancreatic cancer models with a CDK inhibitor SNS-032. SNS-032 improved virus-induced apoptosis and cell death through modulating antiapoptotic signaling pathways and significantly reduced tumor growth in vivo [70].
The propensity of reoviruses to replicate and induce cell death specifically in cells with activated Ras led to the development of Reolysin (Pelareorep). Reolysin, a reovirus serotype-3-Dearing strain targets tumors with Ras pathway mutations which are present in up to 70% of pancreatic cancers. Carew et al. found that Reolysin boosts selective reovirus replication and reduces cell viability in KRas-transformed human pancreatic cells and pancreatic cancer cell lines [71]. Additionally, combining Reolysin with ER stress inducers tunicamycin and bortezomib intensified its anticancer effects in both in vitro and in vivo models [71]. Similarly, Baertsch et al. integrated synthetic microRNA target sites (miRTS) into the genome of oncolytic measles viruses (MVs) virus to enhance selectivity and safety [72]. They developed MV-EGFPmtd with miRTS for miR-122, miR-7, and miR-148a, which are abundant in vital organs. V-EGFPmtd’s replication was inhibited in cell lines and primary hepatocytes expressing these microRNAs but maintained oncolytic potency in pancreatic cancer models. This observation demonstrates the feasibility of targeting multiple microRNAs to modify oncolytic vector tropism without compromising efficacy [72]. Likewise, tumor-targeted oncolytic vaccinia virus (VV) with deleted thymidine kinase armed with murine IL-10 (VVLΔTK-IL-10) was developed. VVLΔTK-IL-10 increased survival in immunocompetent mice [73]. Wang et al. developed an oncolytic herpes simplex virus-1 (oHSV) armed with CD40 ligand (oHSV-CD40L) to potentially enhance the antitumoral immune response in PDAC. CD40L activates immune cells, particularly dendritic cells and T cells, by binding to the CD40 receptor. Intratumoral administration of oHSV-CD40L effectively slowed tumor growth, prolonged survival, increased mature dendritic cells, activated cytotoxic T cells, and reduced regulatory T cells [74]. In an immunocompetent syngeneic PDAC model, oHSV treatment reduced tumor burden and improved survival. Single-cell RNA sequencing and FACS analysis revealed that oHSV decreased tumor-associated macrophages and boosted tumor-infiltrating lymphocytes, including activated CD8+ T and Th1 cells, leading to enhanced PDAC responsiveness to immunotherapy [24].
Vienne et al. explored the oncolytic potential of the fibrotropic minute virus of mice prototype (MVMp) in PDAC [75]. MVMp selectively infected, replicated in, and killed PDAC cells while sparing epithelial-type cells. In immune-competent mouse models, MVMp not only inhibited tumor growth and extended survival but also enhanced immune cell infiltration into tumors, particularly myeloid cells and cytotoxic T cells, with a less exhausted phenotype. The study identified a five-protein classifier (FAK, N-cadherin, E-cadherin, β-catenin, Snail) that predicted a response to MVMp which could guide patient selection for virotherapy [75]. Schäfer et al. evaluated the effectiveness of OVs against PDAC by testing them on fourteen patient-derived PDAC cultures, reflecting the disease’s heterogeneity. Twelve of the fourteen cultures responded to at least one OV, but no single virus was universally superior. Sensitivity to OVs varied by PDAC subtype, with the quasi-mesenchymal/basal-like subtype being more responsive to certain OVs like H-1PV, jin-3, and T-VEC. Key findings included Galectin-1 as a potential biomarker for H-1PV effectiveness and high interferon-stimulated gene (ISG) expression as a marker of resistance to the oncolytic measles virus. Combining the measles virus with a cGAS inhibitor enhanced tumor cell killing. The study highlights the need for personalized treatment strategies due to PDAC’s heterogeneity [76]. These findings collectively illustrate the diverse strategies and promising outcomes of oncolytic viruses in preclinical models of pancreatic cancer. Table 1 presents a non-exhaustive list of preclinical studies based on oncolytic viruses (OVs) in pancreatic cancer.

Table 1.
Preclinical studies investigating various oncolytic viruses as potential therapies for pancreatic cancer.
2.2. Preclinical Trials That Combine OV with Immunotherapy
Programmed cell death-1 (PD-1), programmed cell death-ligand 1 (PD-L1), and cytotoxic T lymphocyte-associated antigen-4 (CTLA-4) are key in regulating immune responses to prevent autoimmunity. Immune checkpoint inhibitors (ICIs) that block these receptors enhance T-cell responses against tumors. OVs not only cause direct tumor cell lysis but also stimulate antitumor immune responses, making them excellent candidates for combination with ICIs and adoptive cell therapy (ACT). OVs can induce ICD, releasing tumor antigens and damage-associated molecular patterns (DAMPs) that promote dendritic cell maturation and T-cell activation. This synergistic combination can potentially overcome monotherapy limitations and improve therapeutic outcomes [86,87]. Recently, numerous research groups have explored the combined use of OV therapy and immunotherapy to treat a variety of cancers, including pancreatic cancers. In this section, we discuss several studies that explored different OVs in combination with immunotherapy or immunomodulatory therapies.
Liu et al. developed an HSV1 OV expressing murine OX40 ligand (OV-mOX40L) [88]. OX40 ligand stimulates T-cell proliferation and activation. OV-mOX40L increased tumor-infiltrating CD4+ T cells, reduced CTL exhaustion and regulatory T cells, reprogrammed pro-inflammatory macrophages and neutrophils, and decreased CAF in mice. Combination of OV-mOX40L with antiIL6 (BE0046) and antiPD-1 (BE0146) agents significantly prolonged survival in PDAC mice, suggesting a promising multimodal therapeutic strategy for PDAC [88].
Similarly, NKT cell activation therapy with an oncolytic vesicular stomatitis virus (VSVΔM51) engineered to express IL-15 (VSV-IL-15) was evaluated in a mouse PDAC model [89]. Here, C57BL/6 mice bearing Panc02 tumors were treated with either VSV-GFP or VSV-IL-15 along with α-GalCer-loaded dendritic cells for NKT cell activation. A group of mice additionally received antiPD-1 antibodies after NKT cell activation. The combination treatment led to better tumor control, validating the superiority of this approach for pancreatic cancer treatment [89].
2.3. OV-Based Clinical Studies in Pancreatic Cancer
The safety, feasibility, and clinical activity of several oncolytic OVs have been evaluated in phase 1 or 2 trials as single agents and in combination with chemotherapy and immunotherapy in patients with pancreatic cancer. The majority of these trials utilized modified oncolytic adenoviruses, while one trial used oncolytic reoviruses. Several selected clinical studies are discussed here.
ONYX-015 (dl1520) adenovirus: ONYX-015 is an adenovirus with a deletion of 55 kDa in the E1B gene that enables selective replication and increased lysis of cancer cells that carry p53 mutations. ONYX-015 (dl1520) was administered directly in primary tumors in patients with unresectable pancreatic cancer in a phase-I (NCT00006106) dose-escalation trial [90]. Multiple doses of the virus were very well tolerated by study subjects up to the maximum doses tested (2 × 1012 particles) [90]. A follow-up phase-I/II trial (NCT00006106) assessed the feasibility, tolerability, and efficacy of ONYX-015 with gemcitabine in 21 patients with unresectable/metastatic pancreatic cancer [91]. Treatment involved eight sessions of ONYX-015 injected via endoscopic ultrasound (EUS) into the primary tumor with the final four sessions in combination with gemcitabine. Overall, the study highlighted the feasibility of transgastric EUS-guided delivery for biological agents and the manageable safety profile of ONYX-015 [91]. Although ONYX-015 showed limited efficacy, these trials demonstrated the feasibility, tolerance, and safety of adenoviral delivery. This success laid the groundwork for future clinical evaluations with more advanced OVs [92].
VCN-01 adenovirus: A phase I trial (NCT02045602) evaluated the safety, maximum tolerated dose (MTD), and recommended phase II dose of VCN-01 for advanced pancreatic adenocarcinoma [58]. VCN-01 is an OAd designed to replicate in cancer cells with defective RB1 pathways and express hyaluronidase. Patients received escalating doses of VCN-01 combined with nab-paclitaxel and gemcitabine. The trial found no dose-limiting toxicities and achieved a 50% overall response rate. The study concluded that VCN-01, in combination with chemotherapy, is feasible, safe, and promising for treating pancreatic adenocarcinoma [93].
Ad5-yCD/mutTKSR39rep-hIL-12 adenovirus: We conducted a phase I trial [93] for metastatic pancreatic cancer using a replication-competent adenovirus Ad5-yCD/mutTKSR39rep-hIL12 that expresses two suicide genes (yeast cytosine deaminase/mutant S39R thymidine kinase from HSV) and human interleukin-12 (IL-12) (NCT03281382). The trial’s primary endpoints were the MTD and dose-limiting toxicities up to day 21. Most of the adverse events were mild, and MTD was not reached. The presence of Ad5-vector DNA indicated viral replication, while an increase in serum levels of IL-12, IFN-γ, and CXCL10 confirmed immune activation/potentiation [93]. Long-term follow-up showed that patients in the highest dose cohort had a higher median overall survival (18.4 months) compared to 4.8 months and 3.5 months in the lower dose cohorts [58].
LOAd703 adenovirus: LOAd703 encodes immunostimulatory genes TMZ-CD40L and 4-1BBL. A nonrandomized phase I/II trial (NCT02705196) assessed the safety and feasibility of LOAd703 with chemotherapy for advanced PDAC [94]. Twenty-one patients received standard chemotherapy (nab-paclitaxel and gemcitabine) and intratumoral LOAd703 injections. The treatment was generally well-tolerated, with common side effects being fever, fatigue, and elevated liver enzymes. No MTD was identified, confirming that the highest dose evaluated was safe. Immunological responses included increased adenovirus-specific T cells and CD8+ effector memory cells. About half (44%) of evaluable patients showed objective responses [94]. Arm 2 of this trial (LOKON001), which includes atezolizumab in addition to nab-paclitaxel plus gemcitabine and LOAd703, is ongoing.
Pelareorep (Reolysin) reovirus: A randomized phase II study (NCT01280058) evaluated pelareorep, an oncolytic reovirus, in combination with carboplatin and paclitaxel in treatment-naive metastatic pancreatic adenocarcinoma patients [95]. Pelareorep was found to be safe; however, it did not improve PFS when combined with carboplatin/paclitaxel (compared to chemo alone). Further targeting of immunosuppressive mediators may enhance oncolytic virotherapy [95].
Recombinant Human Adenovirus Type 5 (H101): Malignant ascites, a complication from peritoneal spread of malignancies, lacks effective treatments. In a phase II trial (NCT04771676), oncolytic adenovirus H101 virotherapy extended the median time to repeat paracentesis to 45 days, showed tolerable toxicity, and enhanced CD8+ T-cell and macrophage immune responses, suggesting the potential for combination with antiPD(L)1 therapy [96].
2.4. Clinical Studies Combining Oncolytic Viruses with Immunotherapy in Pancreatic Cancer
Reolysin was combined with chemotherapy and ICI (pembrolizumab) in patients with relapsed metastatic adenocarcinoma of the pancreas (NCT00998322) in an open-label phase 1b study [97]. Eleven patients received Reolysin, pembrolizumab, and either 5-FU/LV, gemcitabine, or irinotecan, every 3 weeks until disease progression or unacceptable toxicity. The primary endpoint was safety, with secondary objectives including tumor response and immune analysis. Grade 3 or 4 adverse events occurred in 73% of patients, including abdominal pain, anemia, biliary obstruction, and neutropenia. Among five evaluable patients, one achieved a partial response, and two had stable disease. On-treatment biopsies showed reovirus infection in cancer cells and immune infiltrates. Seven patients died due to disease progression [97].
A phase I study from the same group (NCT02620423) evaluated pelareorep (Reolysin) combined with pembrolizumab and chemotherapy in patients with advanced pancreatic cancer [98]. The study aimed to determine safety and efficacy in patients who had progressed after first-line treatment. Eleven patients received pelareorep, pembrolizumab, and either 5-fluorouracil, gemcitabine, or irinotecan. The treatment was well-tolerated, with most adverse events being mild flu-like symptoms. Disease control was achieved in three patients, out of which one had a partial response and two had stable disease. On-treatment biopsies showed reovirus replication, and T-cell receptor sequencing revealed the emergence of new T-cell clones. Patients with clinical benefit showed high peripheral clonality and changes in immune gene expression [98].
MEM-288 is an oncolytic adenovirus vector engineered to selectively replicate in cancer cells and stimulate an antitumor immune response. It encodes human interferon beta (IFNβ) and a recombinant CD40-ligand (MEM40) for immunotherapy against cancer [99]. An ongoing phase I clinical trial of MEM-288 (NCT05076760) consists of two parts. First is an open-label monotherapy dose escalation part that aims to determine the MTD and safety of MEM-288 in patients with various advanced cancers, including pancreatic cancer, advanced/metastatic NSCLC, cutaneous squamous-cell carcinoma (cSCC), Merkel cell, melanoma, triple-negative breast cancer (TNBC), or head and neck cancer, who have progressed following antiPD-1/PD-L1 therapy. Primary objectives include safety, tolerability, and MTD; secondary objectives focus on efficacy measures such as response rates and progression-free survival. The second part of the study is combination therapy with nivolumab, an antiPD-1 therapy. This part will evaluate the efficacy and safety of MEM-288 concurrently with nivolumab in patients with advanced/metastatic NSCLC who have relapsed after initial treatment with antiPD-1/PD-L1 with or without concurrent chemotherapy. The primary goal is to determine the overall response rate, with secondary objectives assessing safety, disease control, and other efficacy outcomes. MEM-288 is administered intratumorally every three weeks, with a maximum of six doses, while nivolumab is given intravenously at a dose of 360 mg every three weeks, with optional maintenance for up to two years [99].
VG161 is an injectable oncolytic HSV-1 therapy that incorporates recombinant human IL-12, IL-15, and PD-L1B [100]. A Chinese multicenter, open-label, phase I/II clinical trial currently recruiting has a single-arm design to assess the safety, tolerability, and preliminary effectiveness of VG161 in combination with the PD-1 inhibitor nivolumab in HSV-seropositive patients with advanced pancreatic cancer. The trial utilizes a standard 3 + 3 dose-escalation design to evaluate the safety of the combination therapy and to determine the recommended phase 2 dose (RP2D) for further efficacy testing. The first cycle of treatment is monitored up to Day 28 for dose-limiting toxicity (DLT). For the efficacy evaluation, a Simon two-stage design is implemented to further explore the preliminary effectiveness of the combination therapy at the established safe dose [100]. These clinical studies collectively illustrate the evolving landscape of OV therapies in pancreatic cancer and highlight advancements in the field that aim at improved patient outcomes. A summary of the clinical trials using OVs in pancreatic cancer therapy is presented in Table 2.
Table 2.
Oncolytic virus-based clinical trials that are actively recruiting/not yet recruiting/completed in locally advanced or metastatic or relapsed pancreatic cancer.
3. Challenges and Future Directions
OVs hold significant potential for transforming pancreatic cancer treatment, but several challenges must be addressed to fully realize their therapeutic benefits. Key considerations include viral selectivity, systemic toxicity, route of administration, host immune response, fibrotic tumor microenvironment, patient selection, and tumor heterogeneity.
A key future challenge is overcoming the obstacle posed by the hypoxic environment within solid tumors, which can hinder viral replication. Pipiya et al. studied the effects of hypoxia in lung, pancreatic, prostate, and colon cancer cells and demonstrated that while hypoxia reduced viral protein production, it did not affect mRNA levels. This underscores the need to address hypoxia-induced limitations to improve the efficacy of oncolytic therapies [101]. Widespread distribution of primary cellular receptors reduces viral selectivity, while systemic toxicity limits the dose that can be safely administered [102]. Although direct intratumoral administration is an invasive process, and thought to be less effective in treating distant metastases, the success of our trials indicates the superiority of this approach [58,103] over intravenous administration that carries risks of liver tropism, which can diminish the virus’s availability in the bloodstream and may cause systemic toxicity [102]. The host immune system poses another significant hurdle, as preexisting or treatment-induced antiviral antibodies, along with complement activation, antiviral cytokines, and macrophages, can neutralize OVs before they reach the tumor [104,105,106]. Although OVs can enhance local oncolysis, the strong immune response they trigger also raises the risk of systemic reactions which may reduce the overall efficiency and safety of the therapy [107,108]. The dense fibrotic tissue characteristic of pancreatic cancer further complicates treatment by hindering the penetration and spread of both OVs and traditional chemotherapies [102]. Components of the ECM such as heparan sulfate and collagen, along with fibrosis, necrosis, and interstitial hydrostatic pressure, create physical barriers that impede the diffusion of OVs to tumor sites [109,110]. Addressing these barriers by modifying the ECM is crucial for improving OV delivery and effectiveness. Patient selection and tumor heterogeneity add another layer of complexity as not all patients respond positively to OV therapy. The variability in tumor types, stages, and inherent patient differences makes it challenging to determine the most suitable OV treatment [110]. Identifying clinical biomarkers is essential for predicting which patients are likely to benefit, thereby improving treatment outcomes [111,112]. Recent studies have also explored the metabolic remodeling effects of OVs. Mahalingam et al. showed that oncolytic vaccinia virus could alter the metabolic profile of tumor cells by reducing glycolysis and inducing metabolic stress, leading to increased immunogenic cell death. By impairing energy production pathways, OVs not only target cancer cells directly but also make the tumor more susceptible to immune-mediated destruction [113]. Thus, exploring OV-mediated metabolic remodeling offers a promising avenue for enhancing OV efficacy and combining it with other therapies, such as metabolic inhibitors or immunotherapies.
Technological advancements, including optimizing viral delivery to tumor sites [114] and combining OVs with targeted nanoparticles [115,116,117], will reduce immune clearance and further enhance therapeutic efficacy. Promising results from multiple clinical trials that combined OVs with chemotherapy, radiotherapy, immune-modulatory and/or immune checkpoint inhibitors (Table 2) [118,119] are poised to become a crucial component for clinical management in advanced pancreatic cancer. Moreover, integrating artificial intelligence (AI) and machine learning (ML) into early screening and detection will help with personalized medicine and significantly advance OV therapy [120,121].
4. Conclusions
OVs represent a promising approach to cancer immunotherapy, with the potential to overcome some of the limitations associated with traditional cancer treatments. By selectively targeting cancer cells and stimulating antitumor immune responses, OVs offer a novel and versatile strategy for treating hard-to-manage solid cancers, including advanced pancreatic cancer. Continued research and clinical development are essential to realize the full therapeutic potential of OVs in cancer management.
Author Contributions
S.T.: original draft and editing; A.R.B.: editing; S.S.: review, feedback; F.S.: review, feedback; S.N.: conceptualization, supervision, editing, feedback. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by NCI R21 (1R21CA252010-01A1), HFHS Research Administration Start Up, HFHS Proposal Development Award, Game on Cancer Award and HFHS-Radiation Oncology Start Up to Shyam Nyati.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| AI | Artificial Intelligence |
| CAFs | Cancer-Associated Fibroblasts |
| CAR | Chimeric Antigen Receptor |
| CAR T | Chimeric Antigen Receptor T |
| CD | Cytosine Deaminase |
| CTLA-4 | Cytotoxic T Lymphocyte-Associated Antigen 4 |
| DAMPs | Damage-Associated Molecular Patterns |
| DCN | Decorin |
| DCs | Dendritic Cells |
| DNA | Deoxyribonucleic Acid |
| ECM | Extracellular Matrix |
| EMA | European Medicines Agency |
| EUS | Endoscopic Ultrasound |
| FDA | Food and Drug Administration |
| FGF2 | Fibroblast Growth Factor 2 |
| GM-CSF | Granulocyte-Macrophage Colony-Stimulating Factor |
| HMGB1 | High Mobility Group Box 1 |
| hMSCs | Human Bone Marrow-Derived Mesenchymal Stromal Cells |
| HSV | Herpes Simplex Viruses |
| ICAM | Intercellular Adhesion Molecule |
| ICIs | Immune Checkpoint Inhibitors |
| FN | Interferon |
| IFN-β | Interferon Beta |
| IFN-γ | Interferon Gamma |
| IL | Interleukin |
| IL-2 | Interleukin-2 |
| IL-6 | Interleukin-6 |
| IL-7 | Interleukin-7 |
| IL-12 | Interleukin-12 |
| MTD | Maximum Tolerated Dose |
| ML | Machine Learning |
| miRTS | MicroRNA Target Sites |
| MV | Measles Virus |
| MVMP | Minute Virus of Mice Prototype |
| NDV | Newcastle Disease Virus |
| NT | Neurotensin |
| OAd | Oncolytic Adenovirus |
| OS | Overall Survival |
| OV | Oncolytic Virus |
| PAMPs | Pathogen-Associated Molecular Patterns |
| PD-1 | Programmed Cell Death Protein 1 |
| PD-L1 | Programmed Death-Ligand 1 |
| PDAC | Pancreatic Ductal Adenocarcinoma |
| PEG | Polyethylene Glycol |
| PFS | Progression-Free Survival |
| RNA | Ribonucleic Acid |
| SAE | Significant Adverse Events |
| SBV | Sindbis Virus |
| SCC | Squamous-Cell Carcinoma |
| SFV | Semliki Forest Virus |
| SMAC | Second Mitochondria-Derived Activator of Caspase |
| SVV | Seneca Valley Virus |
| TILs | Tumor-Infiltrating Lymphocytes |
| TKR | Tumor Necrosis Factor-Alpha |
| TLR | Toll-Like Receptor |
| TME | Tumor Microenvironment |
| TNF-α | Tumor Necrosis Factor-Alpha |
| TRAIL | Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand |
| TK | Thymidine Kinase |
| VV | Vaccinia Virus |
| VSV | Vesicular Stomatitis Virus |
| VVLΔTK-IL-10 | Vaccinia Virus with Deleted Thymidine Kinase and Interleukin-10 Genes |
References
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA Cancer J. Clin. 2024, 74, 12–49. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Feng, Z.; Miao, R.; Liu, X.; Liu, C.; Liu, Z. Prognosis and survival analysis of patients with pancreatic cancer: Retrospective experience of a single institution. World J. Surg. Oncol. 2022, 20, 11. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Carvalho, T.M.A.; Di Molfetta, D.; Greco, M.R.; Koltai, T.; Alfarouk, K.O.; Reshkin, S.J.; Cardone, R.A. Tumor Microenvironment Features and Chemoresistance in Pancreatic Ductal Adenocarcinoma: Insights into Targeting Physicochemical Barriers and Metabolism as Therapeutic Approaches. Cancers 2021, 13, 6135. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Puckett, Y.; Garfield, K. Pancreatic Cancer. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2024. Available online: https://www.ncbi.nlm.nih.gov/books/NBK518996/ (accessed on 9 August 2024).
- Słodkowski, M.; Wroński, M.; Karkocha, D.; Kraj, L.; Śmigielska, K.; Jachnis, A. Current Approaches for the Curative-Intent Surgical Treatment of Pancreatic Ductal Adenocarcinoma. Cancers 2023, 15, 2584. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sinn, M.; Bahra, M.; Liersch, T.; Gellert, K.; Messmann, H.; Bechstein, W.; Waldschmidt, D.; Jacobasch, L.; Wilhelm, M.; Rau, B.M.; et al. CONKO-005: Adjuvant Chemotherapy With Gemcitabine Plus Erlotinib Versus Gemcitabine Alone in Patients After R0 Resection of Pancreatic Cancer: A Multicenter Randomized Phase III Trial. J. Clin. Oncol. 2017, 35, 3330–3337. [Google Scholar] [CrossRef] [PubMed]
- Conroy, T.; Desseigne, F.; Ychou, M.; Bouché, O.; Guimbaud, R.; Bécouarn, Y.; Adenis, A.; Raoul, J.L.; Gourgou-Bourgade, S.; de la Fouchardière, C.; et al. Groupe Tumeurs Digestives of Unicancer; PRODIGE Intergroup. FOLFIRINOX versus gemcitabine for metastatic pancreatic cancer. N. Engl. J. Med. 2011, 364, 1817–1825. [Google Scholar] [CrossRef] [PubMed]
- Von Hoff, D.D.; Ervin, T.; Arena, F.P.; Chiorean, E.G.; Infante, J.; Moore, M.; Seay, T.; Tjulandin, S.A.; Ma, W.W.; Saleh, M.N.; et al. Increased survival in pancreatic cancer with nab-paclitaxel plus gemcitabine. N. Engl. J. Med. 2013, 369, 1691–1703. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liu, Y.; Li, F.; Gao, F.; Xing, L.; Qin, P.; Liang, X.; Zhang, J.; Qiao, X.; Lin, L.; Zhao, Q.; et al. Periostin promotes the chemotherapy resistance to gemcitabine in pancreatic cancer. Tumour Biol. 2016, 37, 15283–15291. [Google Scholar] [CrossRef] [PubMed]
- Hennig, A.; Baenke, F.; Klimova, A.; Drukewitz, S.; Jahnke, B.; Brückmann, S.; Secci, R.; Winter, C.; Schmäche, T.; Seidlitz, T.; et al. Detecting drug resistance in pancreatic cancer organoids guides optimized chemotherapy treatment. J. Pathol. 2022, 257, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Burris, H.A., 3rd; Moore, M.J.; Andersen, J.; Green, M.R.; Rothenberg, M.L.; Modiano, M.R.; Cripps, M.C.; Portenoy, R.K.; Storniolo, A.M.; Tarassoff, P.; et al. Improvements in survival and clinical benefit with gemcitabine as first-line therapy for patients with advanced pancreas cancer: A randomized trial. J. Clin. Oncol. 1997, 15, 2403–2413. [Google Scholar] [CrossRef] [PubMed]
- Karasic, T.B.; O’Hara, M.H.; Loaiza-Bonilla, A.; Reiss, K.A.; Teitelbaum, U.R.; Borazanci, E.; De Jesus-Acosta, A.; Redlinger, C.; Burrell, J.A.; Laheru, D.A.; et al. Effect of Gemcitabine and nab-Paclitaxel With or Without Hydroxychloroquine on Patients With Advanced Pancreatic Cancer: A Phase 2 Randomized Clinical Trial. JAMA Oncol. 2019, 5, 993–998. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kuruppu, D.; Tanabe, K.K. Viral oncolysis by herpes simplex virus and other viruses. Cancer Biol. Ther. 2005, 4, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Alemany, R. Viruses in cancer treatment. Clin. Transl. Oncol. 2013, 15, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Li, D.; Zhu, X. Cancer immunotherapy: Pros, cons and beyond. Biomed. Pharmacother. 2020, 124, 109821. [Google Scholar] [CrossRef] [PubMed]
- Hemminki, O.; Dos Santos, J.M.; Hemminki, A. Oncolytic viruses for cancer immunotherapy. J. Hematol. Oncol. 2020, 13, 84. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Thoidingjam, S.; Sriramulu, S.; Freytag, S.; Brown, S.L.; Kim, J.H.; Chetty, I.J.; Siddiqui, F.; Movsas, B.; Nyati, S. Oncolytic virus-based suicide gene therapy for cancer treatment: A perspective of the clinical trials conducted at Henry Ford Health. Transl. Med. Commun. 2023, 8, 11. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ochiai, H.; Campbell, S.A.; Archer, G.E.; Chewning, T.A.; Dragunsky, E.; Ivanov, A.; Gromeier, M.; Sampson, J.H. Targeted therapy for glioblastoma multiforme neoplastic meningitis with intrathecal delivery of an oncolytic recombinant poliovirus. Clin. Cancer Res. 2006, 12, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Marintcheva, B. Chapter 9—Virus-based therapeutic approaches. In Harnessing the Power of Viruses; Marintcheva, B., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 243–276. [Google Scholar]
- Bai, Y.; Hui, P.; Du, X.; Su, X. Updates to the antitumor mechanism of oncolytic virus. Thorac. Cancer. 2019, 10, 1031–1035. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Marelli, G.; Howells, A.; Lemoine, N.R.; Wang, Y. Oncolytic Viral Therapy and the Immune System: A Double-Edged Sword Against Cancer. Front. Immunol. 2018, 9, 866. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kaufman, H.L.; Kohlhapp, F.J.; Zloza, A. Oncolytic viruses: A new class of immunotherapy drugs. Nat. Rev. Drug Discov. 2015, 14, 642–662. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, X.; Shen, Y.; Wan, X.; Hu, X.; Cai, W.Q.; Wu, Z.; Xin, Q.; Liu, X.; Gui, J.; Xin, H.Y.; et al. Oncolytic virotherapy evolved into the fourth generation as tumor immunotherapy. J. Transl. Med. 2023, 21, 500. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, L.; Wang, W.; Wang, R.; Zhang, N.; Shang, H.; Bi, Y.; Chen, D.; Zhang, C.; Li, L.; Yin, J.; et al. Reshaping the Immune Microenvironment by Oncolytic Herpes Simplex Virus in Murine Pancreatic Ductal Adenocarcinoma. Mol. Ther. 2021, 29, 744–761. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mantwill, K.; Klein, F.G.; Wang, D.; Hindupur, S.V.; Ehrenfeld, M.; Holm, P.S.; Nawroth, R. Concepts in Oncolytic Adenovirus Therapy. Int. J. Mol. Sci. 2021, 22, 10522. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Andtbacka, R.H.; Kaufman, H.L.; Collichio, F.; Amatruda, T.; Senzer, N.; Chesney, J.; Delman, K.A.; Spitler, L.E.; Puzanov, I.; Agarwala, S.S.; et al. Talimogene Laherparepvec Improves Durable Response Rate in Patients With Advanced Melanoma. J. Clin. Oncol. 2015, 33, 2780–2788. [Google Scholar] [CrossRef] [PubMed]
- Kaur, B.; Cripe, T.P.; Chiocca, E.A. “Buy one get one free”: Armed viruses for the treatment of cancer cells and their microenvironment. Curr. Gene Ther. 2009, 9, 341–355. [Google Scholar] [CrossRef]
- Elsedawy, N.B.; Russell, S.J. Oncolytic vaccines. Expert. Rev. Vaccines 2013, 12, 1155–1172. [Google Scholar] [CrossRef] [PubMed]
- Larson, C.; Oronsky, B.; Scicinski, J.; Fanger, G.R.; Stirn, M.; Oronsky, A.; Reid, T.R. Going viral: A review of replication-selective oncolytic adenoviruses. Oncotarget 2015, 6, 19976–19989. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chen, L.; Zuo, M.; Zhou, Q.; Wang, Y. Oncolytic virotherapy in cancer treatment: Challenges and optimization prospects. Front. Immunol. 2023, 14, 1308890. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jiang, B.; Zhou, L.; Lu, J.; Wang, Y.; Liu, C.; You, L.; Guo, J. Stroma-Targeting Therapy in Pancreatic Cancer: One Coin With Two Sides? Front. Oncol. 2020, 10, 576399. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bell, J.; McFadden, G. Viruses for tumor therapy. Cell Host Microbe 2014, 15, 260–265. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fukuhara, H.; Ino, Y.; Todo, T. Oncolytic virus therapy: A new era of cancer treatment at dawn. Cancer Sci. 2016, 107, 1373–1379. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yoo, S.Y.; Badrinath, N.; Lee, H.L.; Heo, J.; Kang, D.H. A Cancer-Favoring, Engineered Vaccinia Virus for Cholangiocarcinoma. Cancers 2019, 11, 1667. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ebelt, N.D.; Zamloot, V.; Manuel, E.R. Targeting desmoplasia in pancreatic cancer as an essential first step to effective therapy. Oncotarget. 2020, 11, 3486–3488. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Erkan, M.; Michalski, C.W.; Rieder, S.; Reiser-Erkan, C.; Abiatari, I.; Kolb, A.; Giese, N.A.; Esposito, I.; Friess, H.; Kleeff, J. The activated stroma index is a novel and independent prognostic marker in pancreatic ductal adenocarcinoma. Clin. Gastroenterol. Hepatol. 2008, 6, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Ullman, N.A.; Burchard, P.R.; Dunne, R.F.; Linehan, D.C. Immunologic Strategies in Pancreatic Cancer: Making Cold Tumors Hot. J. Clin. Oncol. 2022, 40, 2789–2805. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Murakami, T.; Hiroshima, Y.; Matsuyama, R.; Homma, Y.; Hoffman, R.M.; Endo, I. Role of the tumor microenvironment in pancreatic cancer. Ann. Gastroenterol. Surg. 2019, 3, 130–137. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, L.; Chard Dunmall, L.S.; Cheng, Z.; Wang, Y. Remodeling the tumor microenvironment by oncolytic viruses: Beyond oncolysis of tumor cells for cancer treatment. J. Immunother. Cancer. 2022, 10, e004167. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Platanias, L.C. Mechanisms of type-I- and type-II-interferon-mediated signalling. Nat. Rev. Immunol. 2005, 5, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.S.; Liu, Z.; Kowalsky, S.; Feist, M.; Kalinski, P.; Lu, B.; Storkus, W.J.; Bartlett, D.L. Oncolytic Immunotherapy: Conceptual Evolution, Current Strategies, and Future Perspectives. Front. Immunol. 2017, 8, 555. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Breitbach, C.J.; Lichty, B.D.; Bell, J.C. Oncolytic Viruses: Therapeutics With an Identity Crisis. eBioMedicine 2016, 9, 31–36. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Berkey, S.E.; Thorne, S.H.; Bartlett, D.L. Oncolytic Virotherapy and the Tumor Microenvironment. Adv. Exp. Med. Biol. 2017, 1036, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Angarita, F.A.; Acuna, S.A.; Ottolino-Perry, K.; Zerhouni, S.; McCart, J.A. Mounting a strategic offense: Fighting tumor vasculature with oncolytic viruses. Trends Mol. Med. 2013, 19, 378–392. [Google Scholar] [CrossRef] [PubMed]
- Breitbach, C.J.; Arulanandam, R.; De Silva, N.; Thorne, S.H.; Patt, R.; Daneshmand, M.; Moon, A.; Ilkow, C.; Burke, J.; Hwang, T.H.; et al. Oncolytic vaccinia virus disrupts tumor-associated vasculature in humans. Cancer Res. 2013, 73, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Ilkow, C.S.; Marguerie, M.; Batenchuk, C.; Mayer, J.; Ben Neriah, D.; Cousineau, S.; Falls, T.; Jennings, V.A.; Boileau, M.; Bellamy, D.; et al. Reciprocal cellular cross-talk within the tumor microenvironment promotes oncolytic virus activity. Nat. Med. 2015, 21, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.H.; Choi, I.K.; Lee, H.S.; Yan, H.H.; Son, M.K.; Ahn, H.M.; Hong, J.; Yun, C.O.; Hong, S.S. Oncolytic adenovirus expressing relaxin (YDC002) enhances therapeutic efficacy of gemcitabine against pancreatic cancer. Cancer Lett. 2017, 396, 155–166, Erratum in: Cancer Lett. 2017, 404, 93. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.K.; Lee, Y.S.; Yoo, J.Y.; Yoon, A.R.; Kim, H.; Kim, D.S.; Seidler, D.G.; Kim, J.H.; Yun, C.O. Effect of decorin on overcoming the extracellular matrix barrier for oncolytic virotherapy. Gene Ther. 2010, 17, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hong, J.; Jung, B.K.; Oh, E.; Yun, C.O. Oncolytic Ad co-expressing decorin and Wnt decoy receptor overcomes chemoresistance of desmoplastic tumor through degradation of ECM and inhibition of EMT. Cancer Lett. 2019, 459, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Al-Zaher, A.A.; Moreno, R.; Fajardo, C.A.; Arias-Badia, M.; Farrera, M.; de Sostoa, J.; Rojas, L.A.; Alemany, R. Evidence of Anti-tumoral Efficacy in an Immune Competent Setting with an iRGD-Modified Hyaluronidase-Armed Oncolytic Adenovirus. Mol. Ther. Oncolytics 2018, 8, 62–70. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bazan-Peregrino, M.; Garcia-Carbonero, R.; Laquente, B.; Álvarez, R.; Mato-Berciano, A.; Gimenez-Alejandre, M.; Morgado, S.; Rodríguez-García, A.; Maliandi, M.V.; Riesco, M.C.; et al. VCN-01 disrupts pancreatic cancer stroma and exerts antitumor effects. J. Immunother. Cancer. 2021, 9, e003254. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rahal, A.; Musher, B. Oncolytic viral therapy for pancreatic cancer. J. Surg. Oncol. 2017, 116, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Dock, G. The Influence of Complicating Diseases Upon Leukaemia; Lea Brothers & Company: Cambridge, UK, 1904. [Google Scholar]
- Zhang, S.; Rabkin, S.D. The discovery and development of oncolytic viruses: Are they the future of cancer immunotherapy? Expert Opin. Drug. Discov. 2021, 16, 391–410. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Martuza, R.L.; Malick, A.; Markert, J.M.; Ruffner, K.L.; Coen, D.M. Experimental therapy of human glioma by means of a genetically engineered virus mutant. Science 1991, 252, 854–856. [Google Scholar] [CrossRef] [PubMed]
- Bommareddy, P.K.; Peters, C.; Saha, D.; Rabkin, S.D.; Kaufman, H.L. Oncolytic Herpes Simplex Viruses as a Paradigm for the Treatment of Cancer. Annu. Rev. Cancer Biol. 2018, 2, 155–173. [Google Scholar] [CrossRef]
- Nasar, R.T.; Uche, I.K.; Kousoulas, K.G. Targeting Cancers with oHSV-Based Oncolytic Viral Immunotherapy. Curr. Issues Mol. Biol. 2024, 46, 5582–5594. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Barton, K.N.; Siddiqui, F.; Pompa, R.; Freytag, S.O.; Khan, G.; Dobrosotskaya, I.; Ajlouni, M.; Zhang, Y.; Cheng, J.; Movsas, B.; et al. Phase I trial of oncolytic adenovirus-mediated cytotoxic and interleukin-12 gene therapy for the treatment of metastatic pancreatic cancer. Mol. Ther. Oncolytics 2020, 20, 94–104. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- OBryan, S.M.; Mathis, J.M. Oncolytic Virotherapy for Breast Cancer Treatment. Curr. Gene Ther. 2018, 18, 192–205. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Asija, S.; Chatterjee, A.; Goda, J.S.; Yadav, S.; Chekuri, G.; Purwar, R. Oncolytic immunovirotherapy for high-grade gliomas: A novel and an evolving therapeutic option. Front. Immunol. 2023, 14, 1118246. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Dai, B.; Roife, D.; Kang, Y.; Gumin, J.; Rios Perez, M.V.; Li, X.; Pratt, M.; Brekken, R.A.; Fueyo-Margareto, J.; Lang, F.F.; et al. Preclinical Evaluation of Sequential Combination of Oncolytic Adenovirus Delta-24-RGD and Phosphatidylserine-Targeting Antibody in Pancreatic Ductal Adenocarcinoma. Mol. Cancer Ther. 2017, 16, 662–670. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Na, Y.; Choi, J.W.; Kasala, D.; Hong, J.; Oh, E.; Li, Y.; Jung, S.J.; Kim, S.W.; Yun, C.O. Potent antitumor effect of neurotensin receptor-targeted oncolytic adenovirus co-expressing decorin and Wnt antagonist in an orthotopic pancreatic tumor model. J. Control Release 2015, 220 Pt B, 766–782. [Google Scholar] [CrossRef] [PubMed]
- Na, Y.; Nam, J.P.; Hong, J.; Oh, E.; Shin, H.C.; Kim, H.S.; Kim, S.W.; Yun, C.O. Systemic administration of human mesenchymal stromal cells infected with polymer-coated oncolytic adenovirus induces efficient pancreatic tumor homing and infiltration. J. Control Release 2019, 305, 75–88. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Brugada-Vilà, P.; Cascante, A.; Lázaro, M.Á.; Castells-Sala, C.; Fornaguera, C.; Rovira-Rigau, M.; Albertazzi, L.; Borros, S.; Fillat, C. Oligopeptide-modified poly(beta-amino ester)s-coated AdNuPARmE1A: Boosting the efficacy of intravenously administered therapeutic adenoviruses. Theranostics 2020, 10, 2744–2758. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Salzwedel, A.O.; Han, J.; LaRocca, C.J.; Shanley, R.; Yamamoto, M.; Davydova, J. Combination of interferon-expressing oncolytic adenovirus with chemotherapy and radiation is highly synergistic in hamster model of pancreatic cancer. Oncotarget 2018, 9, 18041–18052. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Watanabe, K.; Luo, Y.; Da, T.; Guedan, S.; Ruella, M.; Scholler, J.; Keith, B.; Young, R.M.; Engels, B.; Sorsa, S.; et al. Pancreatic cancer therapy with combined mesothelin-redirected chimeric antigen receptor T cells and cytokine-armed oncolytic adenoviruses. JCI Insight 2018, 3, e99573. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Raimondi, G.; Gea-Sorlí, S.; Otero-Mateo, M.; Fillat, C. Inhibition of miR-222 by Oncolytic Adenovirus-Encoded miRNA Sponges Promotes Viral Oncolysis and Elicits Antitumor Effects in Pancreatic Cancer Models. Cancers 2021, 13, 3233. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Doerner, J.; Sallard, E.; Zhang, W.; Solanki, M.; Liu, J.; Ehrke-Schulz, E.; Zirngibl, H.; Lieber, A.; Ehrhardt, A. Novel Group C Oncolytic Adenoviruses Carrying a miRNA Inhibitor Demonstrate Enhanced Oncolytic Activity In Vitro and In Vivo. Mol. Cancer Ther. 2022, 21, 460–470. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hashimoto, M.; Kuroda, S.; Kanaya, N.; Kadowaki, D.; Yoshida, Y.; Sakamoto, M.; Hamada, Y.; Sugimoto, R.; Yagi, C.; Ohtani, T.; et al. Long-term activation of anti-tumor immunity in pancreatic cancer by a p53-expressing telomerase-specific oncolytic adenovirus. Br. J. Cancer 2024, 130, 1187–1195. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ge, Y.; Lei, W.; Ma, Y.; Wang, Y.; Wei, B.; Chen, X.; Ru, G.; He, X.; Mou, X.; Wang, S. Synergistic antitumor effects of CDK inhibitor SNS-032 and an oncolytic adenovirus co-expressing TRAIL and Smac in pancreatic cancer. Mol. Med. Rep. 2017, 15, 3521–3528. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Carew, J.S.; Espitia, C.M.; Zhao, W.; Kelly, K.R.; Coffey, M.; Freeman, J.W.; Nawrocki, S.T. Reolysin is a novel reovirus-based agent that induces endoplasmic reticular stress-mediated apoptosis in pancreatic cancer. Cell Death Dis. 2013, 4, e728. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Baertsch, M.A.; Leber, M.F.; Bossow, S.; Singh, M.; Engeland, C.E.; Albert, J.; Grossardt, C.; Jäger, D.; von Kalle, C.; Ungerechts, G. MicroRNA-mediated multi-tissue detargeting of oncolytic measles virus. Cancer Gene Ther. 2014, 21, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Chard, L.S.; Lemoine, N.R.; Wang, Y. New role of Interleukin-10 in enhancing the antitumor efficacy of oncolytic vaccinia virus for treatment of pancreatic cancer. Oncoimmunology 2015, 4, e1038689. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, R.; Chen, J.; Wang, W.; Zhao, Z.; Wang, H.; Liu, S.; Li, F.; Wan, Y.; Yin, J.; Wang, R.; et al. CD40L-armed oncolytic herpes simplex virus suppresses pancreatic ductal adenocarcinoma by facilitating the tumor microenvironment favorable to cytotoxic T cell response in the syngeneic mouse model. J. Immunother. Cancer 2022, 10, e003809. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Vienne, M.; Lopez, C.; Lulka, H.; Nevot, A.; Labrousse, G.; Dusetti, N.; Buscail, L.; Cordelier, P. Minute virus of mice shows oncolytic activity against pancreatic cancer cells exhibiting a mesenchymal phenotype. Mol. Ther. Oncol. 2024, 32, 200780. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Schäfer, T.E.; Knol, L.I.; Haas, F.V.; Hartley, A.; Pernickel, S.C.S.; Jády, A.; Finkbeiner, M.S.C.; Achberger, J.; Arelaki, S.; Modic, Ž.; et al. Biomarker screen for efficacy of oncolytic virotherapy in patient-derived pancreatic cancer cultures. eBioMedicine 2024, 105, 105219. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chard, L.S.; Maniati, E.; Wang, P.; Zhang, Z.; Gao, D.; Wang, J.; Cao, F.; Ahmed, J.; El Khouri, M.; Hughes, J.; et al. A vaccinia virus armed with interleukin-10 is a promising therapeutic agent for treatment of murine pancreatic cancer. Clin. Cancer Res. 2015, 21, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Rovira-Rigau, M.; Raimondi, G.; Marín, M.Á.; Gironella, M.; Alemany, R.; Fillat, C. Bioselection Reveals miR-99b and miR-485 as Enhancers of Adenoviral Oncolysis in Pancreatic Cancer. Mol. Ther. 2019, 27, 230–243. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Man, Y.K.S.; Davies, J.A.; Coughlan, L.; Pantelidou, C.; Blázquez-Moreno, A.; Marshall, J.F.; Parker, A.L.; Halldén, G. The Novel Oncolytic Adenoviral Mutant Ad5-3Δ-A20T Retargeted to αvβ6 Integrins Efficiently Eliminates Pancreatic Cancer Cells. Mol. Cancer Ther. 2018, 17, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Chu, L.; Yuan, S.; Yang, Y.; Yang, Y.; Xu, B.; Zhang, K.; Liu, X.Y.; Wang, R.; Fang, L.; et al. RGD-modified oncolytic adenovirus-harboring shPKM2 exhibits a potent cytotoxic effect in pancreatic cancer via autophagy inhibition and apoptosis promotion. Cell Death Dis. 2017, 8, e2835. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Han, Z.; Lee, S.; Je, S.; Eom, C.Y.; Choi, H.J.; Song, J.J.; Kim, J.H. Survivin silencing and TRAIL expression using oncolytic adenovirus increase anti-tumorigenic activity in gemcitabine-resistant pancreatic cancer cells. Apoptosis 2016, 21, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Cataldi, M.; Shah, N.R.; Felt, S.A.; Grdzelishvili, V.Z. Breaking resistance of pancreatic cancer cells to an attenuated vesicular stomatitis virus through a novel activity of IKK inhibitor TPCA-1. Virology 2015, 485, 340–354. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Singh, H.M.; Leber, M.F.; Bossow, S.; Engeland, C.E.; Dessila, J.; Grossardt, C.; Zaoui, K.; Bell, J.C.; Jäger, D.; von Kalle, C.; et al. MicroRNA-sensitive oncolytic measles virus for chemovirotherapy of pancreatic cancer. Mol. Ther. Oncolytics 2021, 21, 340–355. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, J.; Bonifati, S.; Hristov, G.; Marttila, T.; Valmary-Degano, S.; Stanzel, S.; Schnölzer, M.; Mougin, C.; Aprahamian, M.; Grekova, S.P.; et al. Synergistic combination of valproic acid and oncolytic parvovirus H-1PV as a potential therapy against cervical and pancreatic carcinomas. EMBO Mol. Med. 2013, 5, 1537–1555. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Dai, M.H.; Liu, S.L.; Chen, N.G.; Zhang, T.P.; You, L.; QZhang, F.; Chou, T.C.; Szalay, A.A.; Fong, Y.; Zhao, Y.P. Oncolytic vaccinia virus in combination with radiation shows synergistic antitumor efficacy in pancreatic cancer. Cancer Lett. 2014, 344, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Seidel, J.A.; Otsuka, A.; Kabashima, K. Anti-PD-1 and Anti-CTLA-4 Therapies in Cancer: Mechanisms of Action, Efficacy, and Limitations. Front. Oncol. 2018, 8, 86. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Palanivelu, L.; Liu, C.H.; Lin, L.T. Immunogenic cell death: The cornerstone of oncolytic viro-immunotherapy. Front. Immunol. 2023, 13, 1038226. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liu, S.; Li, F.; Ma, Q.; Du, M.; Wang, H.; Zhu, Y.; Deng, L.; Gao, W.; Wang, C.; Liu, Y.; et al. OX40L-Armed Oncolytic Virus Boosts T-cell Response and Remodels Tumor Microenvironment for Pancreatic Cancer Treatment. Theranostics 2023, 13, 4016–4029. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nelson, A.; Gebremeskel, S.; Lichty, B.D.; Johnston, B. Natural killer T cell immunotherapy combined with IL-15-expressing oncolytic virotherapy and PD-1 blockade mediates pancreatic tumor regression. J. Immunother. Cancer 2022, 10, e003923. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mulvihill, S.; Warren, R.; Venook, A.; Adler, A.; Randlev, B.; Heise, C.; Kirn, D. Safety and feasibility of injection with an E1B-55 kDa gene-deleted, replication-selective adenovirus (ONYX-015) into primary carcinomas of the pancreas: A phase I trial. Gene Ther. 2001, 8, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Hecht, J.R.; Bedford, R.; Abbruzzese, J.L.; Lahoti, S.; Reid, T.R.; Soetikno, R.M.; Kirn, D.H.; Freeman, S.M. A phase I/II trial of intratumoral endoscopic ultrasound injection of ONYX-015 with intravenous gemcitabine in unresectable pancreatic carcinoma. Clin. Cancer Res. 2003, 9, 555–561. [Google Scholar] [PubMed]
- Nattress, C.B.; Halldén, G. Advances in oncolytic adenovirus therapy for pancreatic cancer. Cancer Lett. 2018, 434, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Carbonero, R.; Bazan-Peregrino, M.; Gil-Martín, M.; Álvarez, R.; Macarulla, T.; Riesco-Martinez, M.C.; Verdaguer, H.; Guillén-Ponce, C.; Farrera-Sal, M.; Moreno, R.; et al. Phase I, multicenter, open-label study of intravenous VCN-01 oncolytic adenovirus with or without nab-paclitaxel plus gemcitabine in patients with advanced solid tumors. J. Immunother. Cancer 2022, 10, e003255. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Musher, B.L.; Rowinsky, E.K.; Smaglo, B.G.; Abidi, W.; Othman, M.; Patel, K.; Jawaid, S.; Jing, J.; Brisco, A.; Leen, A.M.; et al. LOAd703, an oncolytic virus-based immunostimulatory gene therapy, combined with chemotherapy for unresectable or metastatic pancreatic cancer (LOKON001): Results from arm 1 of a non-randomised, single-centre, phase 1/2 study. Lancet Oncol. 2024, 25, 488–500. [Google Scholar] [CrossRef] [PubMed]
- Noonan, A.M.; Farren, M.R.; Geyer, S.M.; Huang, Y.; Tahiri, S.; Ahn, D.; Mikhail, S.; Ciombor, K.K.; Pant, S.; Aparo, S.; et al. Randomized Phase 2 Trial of the Oncolytic Virus Pelareorep (Reolysin) in Upfront Treatment of Metastatic Pancreatic Adenocarcinoma. Mol. Ther. 2016, 24, 1150–1158. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, Y.; Qian, L.; Chen, K.; Gu, S.; Meng, Z.; Wang, J.; Li, Y.; Wang, P. Oncolytic adenovirus in treating malignant ascites: A phase II trial and longitudinal single-cell study. Mol. Ther. 2024, 32, 2000–2020. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mahalingam, D.; Fountzilas, C.; Moseley, J.L.; Noronha, N.; Cheetham, K.; Dzugalo, A.; Nuovo, G.; Gutierrez, A.; Arora, S.P. A study of REOLYSIN in combination with pembrolizumab and chemotherapy in patients (pts) with relapsed metastatic adenocarcinoma of the pancreas (MAP). J. Clin. Oncol. 2017, 35, e15753. [Google Scholar] [CrossRef]
- Mahalingam, D.; Wilkinson, G.A.; Eng, K.H.; Fields, P.; Raber, P.; Moseley, J.L.; Cheetham, K.; Coffey, M.; Nuovo, G.; Kalinski, P.; et al. Pembrolizumab in Combination with the Oncolytic Virus Pelareorep and Chemotherapy in Patients with Advanced Pancreatic Adenocarcinoma: A Phase Ib Study. Clin. Cancer Res. 2020, 26, 71–81. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Saltos, A.N.; Arrowood, C.; Beasley, G.; Ronald, J.; El-Haddad, G.; Guerra-Guevara, L.; Ready, N.E. A phase 1 first-in-human study of interferon beta (IFNβ) and membrane-stable CD40L expressing oncolytic virus (MEM-288) in solid tumors including non–small-cell lung cancer (NSCLC). J. Clin. Oncol. 2023, 41 (Suppl. 16), 2569. [Google Scholar] [CrossRef]
- Shen, Y.; Qin, A.; Qiu, Y.; Jin, X.; Song, W.; Fang, T.; Liang, T. 766 A clinical trial to evaluate the safety, tolerability and preliminary efficacy of VG161 in combination with Nivolumab in patients with advanced pancreatic cancer. J. ImmunoTherapy Cancer 2023, 11 (Suppl. 1), A861. [Google Scholar]
- Pipiya, T.; Sauthoff, H.; Huang, Y.Q.; Chang, B.; Cheng, J.; Heitner, S.; Chen, S.; Rom, W.N.; Hay, J.G. Hypoxia reduces adenoviral replication in cancer cells by downregulation of viral protein expression. Gene Ther. 2005, 12, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Taylor, I.P.; Lopez, J.A. Oncolytic adenoviruses and the treatment of pancreatic cancer: A review of clinical trials. J. Cancer Res. Clin. Oncol. 2023, 149, 8117–8129. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bhatnagar, A.R.; Siddiqui, F.; Khan, G.; Pompa, R.; Kwon, D.; Nyati, S. Long-Term Follow-Up of Phase I Trial of Oncolytic Adenovirus-Mediated Cytotoxic and Interleukin-12 Gene Therapy for Treatment of Metastatic Pancreatic Cancer. Biomedicines 2024, 12, 1065. [Google Scholar] [CrossRef]
- Groeneveldt, C.; van den Ende, J.; van Montfoort, N. Preexisting immunity: Barrier or bridge to effective oncolytic virus therapy? Cytokine Growth Factor. Rev. 2023, 70, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, S.; Ghorbani, E.; Khazaei, M.; Avan, A.; Ryzhikov, M.; Azadmanesh, K.; Hassanian, S.M. Interferon-Mediated Tumor Resistance to Oncolytic Virotherapy. J. Cell Biochem. 2017, 118, 1994–1999. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, C.; Iankov, I.D.; Anderson, S.K.; Aderca, I.; Leontovich, A.A.; Maurer, M.J.; Oberg, A.L.; Schroeder, M.A.; Giannini, C.; Greiner, S.M.; et al. Constitutive Interferon Pathway Activation in Tumors as an Efficacy Determinant Following Oncolytic Virotherapy. J. Natl. Cancer Inst. 2018, 110, 1123–1132. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liu, M.; Zhang, R.; Huang, H.; Liu, P.; Zhao, X.; Wu, H.; He, Y.; Xu, R.; Qin, X.; Cheng, Z.; et al. Erythrocyte-Leveraged Oncolytic Virotherapy (ELeOVt): Oncolytic Virus Assembly on Erythrocyte Surface to Combat Pulmonary Metastasis and Alleviate Side Effects. Adv. Sci. 2024, 11, e2303907. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bounassar-Filho, J.P.; Boeckler-Troncoso, L.; Cajigas-Gonzalez, J.; Zavala-Cerna, M.G. SARS-CoV-2 as an Oncolytic Virus Following Reactivation of the Immune System: A Review. Int. J. Mol. Sci. 2023, 24, 2326. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yaacov, B.; Lazar, I.; Tayeb, S.; Frank, S.; Izhar, U.; Lotem, M.; Perlman, R.; Ben-Yehuda, D.; Zakay-Rones, Z.; Panet, A. Extracellular matrix constituents interfere with Newcastle disease virus spread in solid tissue and diminish its potential oncolytic activity. J. Gen. Virol. 2012, 93 Pt 8, 1664–1672. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Zhang, Z.; Chen, Y.; Xu, J.; Wang, J.; Wang, Z. Enhancing cancer therapy: The integration of oncolytic virus therapy with diverse treatments. Cancer Cell Int. 2024, 24, 242. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhu, Z.; McGray, A.J.R.; Jiang, W.; Lu, B.; Kalinski, P.; Guo, Z.S. Improving cancer immunotherapy by rationally combining oncolytic virus with modulators targeting key signaling pathways. Mol. Cancer 2022, 21, 196. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kaufman, H.L. Can Biomarkers Guide Oncolytic Virus Immunotherapy? Clin. Cancer Res. 2021, 27, 3278–3279. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, D.; Fountzilas, C.; Moseley, J.; Noronha, N.; Tran, H.; Chakrabarty, R.; Selvaggi, G.; Coffey, M.; Thompson, B.; Sarantopoulos, J. A phase II study of REOLYSIN® (pelareorep) in combination with carboplatin and paclitaxel for patients with advanced malignant melanoma. Cancer Chemother. Pharmacol. 2017, 79, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Huang, J.; Tong, A.; Yang, H. Oncolytic Viruses for Cancer Therapy: Barriers and Recent Advances. Mol. Ther. Oncolytics 2019, 15, 234–247. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tian, Y.; Xie, D.; Yang, L. Engineering strategies to enhance oncolytic viruses in cancer immunotherapy. Signal Transduct. Target. Ther. 2022, 7, 117. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fang, C.; Xiao, G.; Wang, T.; Song, L.; Peng, B.; Xu, B.; Zhang, K. Emerging Nano-/Biotechnology Drives Oncolytic Virus-Activated and Combined Cancer Immunotherapy. Research 2023, 6, 0108. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ajam-Hosseini, M.; Akhoondi, F.; Doroudian, M. Nano based-oncolytic viruses for cancer therapy. Crit. Rev. Oncol. Hematol. 2023, 185, 103980. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jiang, Z.; Zhang, Y.; Huang, X.; Liu, Q. Efficacy and Safety of Oncolytic Viruses in Randomized Controlled Trials: A Systematic Review and Meta-Analysis. Cancers 2020, 12, 1416. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lichty, B.D.; Breitbach, C.J.; Stojdl, D.F.; Bell, J.C. Going viral with cancer immunotherapy. Nat. Rev. Cancer. 2014, 14, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Kourou, K.; Exarchos, T.P.; Exarchos, K.P.; Karamouzis, M.V.; Fotiadis, D.I. Machine learning applications in cancer prognosis and prediction. Comput. Struct. Biotechnol. J. 2014, 13, 8–17. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kaviarasan, V.; Ragunath, B.; Veerabathiran, R. Chapter 4—Oncolytic virus cancer therapeutic options and integration of artificial intelligence into virus cancer research. In Oncogenic Viruses; Ennaji, M.M., Ed.; Academic Press: Cambridge, MA, USA, 2023; pp. 61–80. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).