
The Potential Related Genes and Mechanisms Involved in Improving the Treadmill Exercise Ability of APP/PS1 Mice

Abstract
:1. Introduction
2. Results
2.1. Treadmill Exercise Improved Memory and Reduced the Anxiety Behavior of AD Mice
2.2. Treadmill Exercise Improved the Exercise Ability and Alleviated the Skeletal Muscle Atrophy in AD Mice
2.3. Treadmill Exercise Increased the Expression of Genes and Proteins Related to Skeletal Muscle Protein Synthesis in the AD Mice
2.4. Treadmill Exercise Increased the Expressions of Genes and Proteins Related to Skeletal Muscle Protein Degradation in the AD Mice
2.5. Treadmill Exercise Altered Gene Expression in the Skeletal Muscle of AD Mice
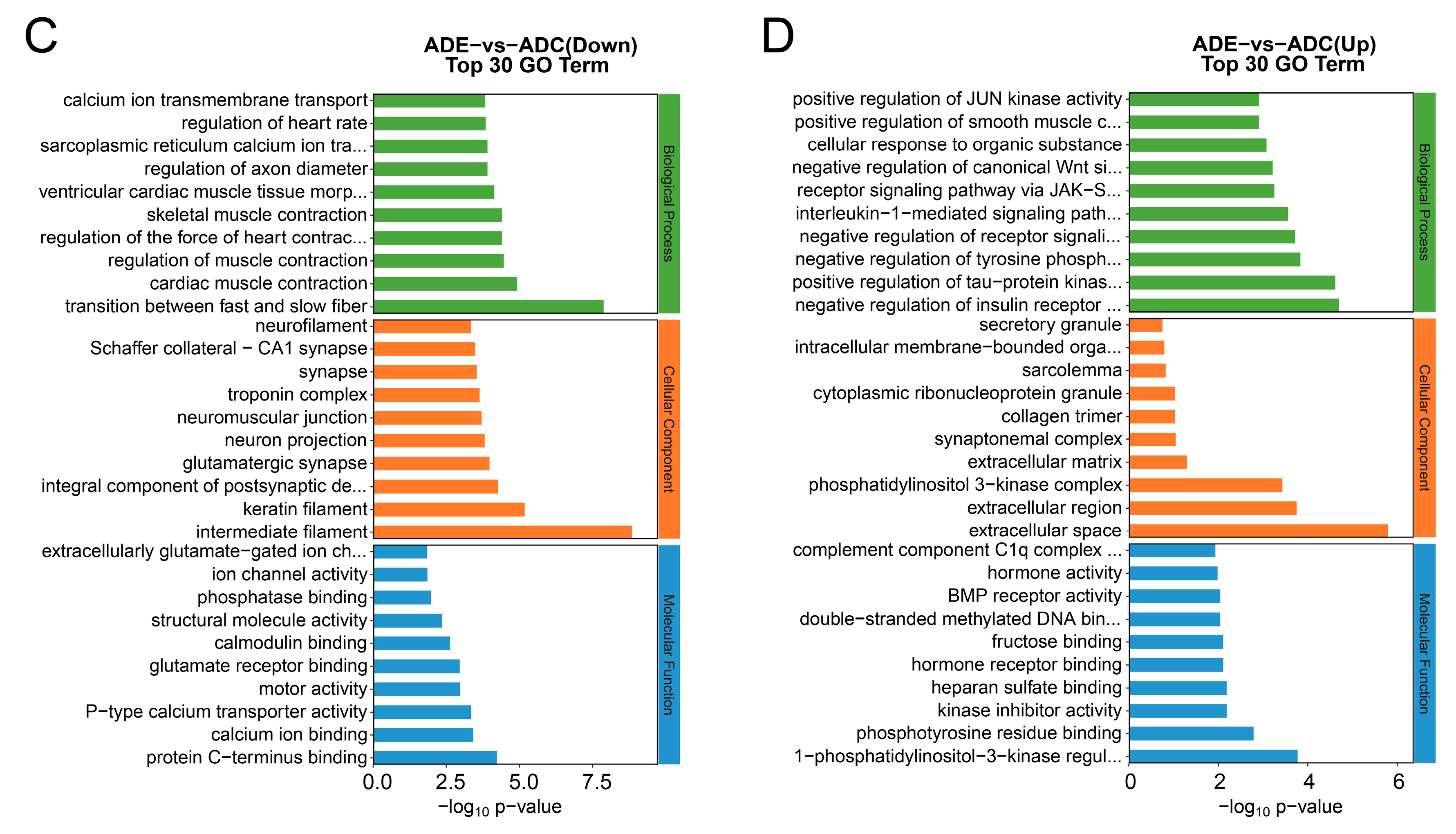
2.6. GO Enrichment Analysis of the DEGs
2.7. KEGG Enrichment Analysis of the DEGs
2.8. Transcription Factor Analysis
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Groups
4.2. Exercise Intervention
4.3. Morris Water Maze
4.4. Open Field Test
4.5. Exercise Ability Test
4.6. Tissue Collection
4.7. Experimental Reagents, Instruments, and Methods
4.7.1. HE Staining and Masson Staining
4.7.2. Wheat Germ Agglutinin (WGA) Staining
4.7.3. Transmission Electron Microscopy
4.7.4. Real-Time PCR
4.7.5. Western Blotting
4.7.6. RNA-Seq
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bellenguez, C.; Kucukali, F.; Jansen, I.E.; Kleineidam, L.; Moreno-Grau, S.; Amin, N.; Naj, A.C.; Campos-Martin, R.; Grenier-Boley, B.; Andrade, V.; et al. New insights into the genetic etiology of Alzheimer’s disease and related dementias. Nat. Genet. 2022, 54, 412–436. [Google Scholar] [CrossRef] [PubMed]
- Ren, R.; Qi, J.; Lin, S.; Liu, X.; Yin, P.; Wang, Z.; Tang, R.; Wang, J.; Huang, Q.; Li, J.; et al. The China Alzheimer Report 2022. Gen. Psychiatry 2022, 35, e100751. [Google Scholar] [CrossRef]
- Scheltens, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chetelat, G.; Teunissen, C.E.; Cummings, J.; Van Der Flier, W.M. Alzheimer’s disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef] [PubMed]
- Joe, E.; Ringman, J.M. Cognitive symptoms of Alzheimer’s disease: Clinical management and prevention. BMJ 2019, 367, l6217. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Provenzano, F.A.; Small, S.A.; Alzheimer’s Disease Neuroimaging Initiative. A deep learning MRI approach outperforms other biomarkers of prodromal Alzheimer’s disease. Alzheimers Res. Ther. 2022, 14, 45. [Google Scholar] [CrossRef]
- Liu, Q.; Xie, T.; Xi, Y.; Li, L.; Mo, F.; Liu, X.; Liu, Z.; Gao, J.M.; Yuan, T. Sesamol Attenuates Amyloid Peptide Accumulation and Cognitive Deficits in APP/PS1 Mice: The Mediating Role of the Gut-Brain Axis. J. Agric. Food Chem. 2021, 69, 12717–12729. [Google Scholar] [CrossRef]
- Yang, J.Y.; Baek, S.E.; Yoon, J.W.; Kim, H.S.; Kwon, Y.; Yeom, E. Nesfatin-1 ameliorates pathological abnormalities in Drosophila hTau model of Alzheimer’s disease. Biochem. Biophys. Res. Commun. 2024, 727, 150311. [Google Scholar] [CrossRef]
- Yuan, S.; Wang, Y.; Yang, J.; Tang, Y.; Wu, W.; Meng, X.; Jian, Y.; Lei, Y.; Liu, Y.; Tang, C.; et al. Treadmill exercise can regulate the redox balance in the livers of APP/PS1 mice and reduce LPS accumulation in their brains through the gut-liver-kupffer cell axis. Aging 2024, 16, 1374–1389. [Google Scholar] [CrossRef]
- Salinas-Rodriguez, A.; Palazuelos-Gonzalez, R.; Rivera-Almaraz, A.; Manrique-Espinoza, B. Longitudinal association of sarcopenia and mild cognitive impairment among older Mexican adults. J. Cachexia Sarcopenia Muscle 2021, 12, 1848–1859. [Google Scholar] [CrossRef]
- Hu, Y.; Peng, W.; Ren, R.; Wang, Y.; Wang, G. Sarcopenia and mild cognitive impairment among elderly adults: The first longitudinal evidence from CHARLS. J. Cachexia Sarcopenia Muscle 2022, 13, 2944–2952. [Google Scholar] [CrossRef]
- Brisendine, M.H.; Nichenko, A.S.; Bandara, A.B.; Willoughby, O.S.; Amiri, N.; Weingrad, Z.; Specht, K.S.; Bond, J.M.; Addington, A.; Jones, R.G., 3rd; et al. Neuromuscular Dysfunction Precedes Cognitive Impairment in a Mouse Model of Alzheimer’s Disease. Function 2024, 5, zqad066. [Google Scholar] [CrossRef] [PubMed]
- Jun, L.; Robinson, M.; Geetha, T.; Broderick, T.L.; Babu, J.R. Prevalence and Mechanisms of Skeletal Muscle Atrophy in Metabolic Conditions. Int. J. Mol. Sci. 2023, 24, 2973. [Google Scholar] [CrossRef] [PubMed]
- Longo, S.; Messi, M.L.; Wang, Z.M.; Meeker, W.; Delbono, O. Accelerated sarcopenia precedes learning and memory impairments in the P301S mouse model of tauopathies and Alzheimer’s disease. J. Cachexia Sarcopenia Muscle 2024, 15, 1358–1375. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lim, Y. The Potential Role of Myokines/Hepatokines in the Progression of Neuronal Damage in Streptozotocin and High-Fat Diet-Induced Type 2 Diabetes Mellitus Mice. Biomedicines 2022, 10, 1521. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wong, P.Y.; Chow, S.K.H.; Cheung, W.H.; Wong, R.M.Y. Does the regulation of skeletal muscle influence cognitive function? A scoping review of pre-clinical evidence. J. Orthop. Transl. 2023, 38, 76–83. [Google Scholar] [CrossRef]
- Wang, M.; Hua, Y.; Bai, Y. A review of the application of exercise intervention on improving cognition in patients with Alzheimer’s disease: Mechanisms and clinical studies. Rev. Neurosci. 2024; ahead of print. [Google Scholar] [CrossRef]
- Nicola, L.; Loo, S.J.Q.; Lyon, G.; Turknett, J.; Wood, T.R. Does resistance training in older adults lead to structural brain changes associated with a lower risk of Alzheimer’s dementia? A narrative review. Ageing Res. Rev. 2024, 98, 102356. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 2015, 96, e52434. [Google Scholar]
- Sartori, R.; Romanello, V.; Sandri, M. Mechanisms of muscle atrophy and hypertrophy: Implications in health and disease. Nat. Commun. 2021, 12, 330. [Google Scholar] [CrossRef]
- Abe, K. Dynamic Activity Changes in Transcription Factors: Unlocking the Mechanisms Regulating Physiological Changes in the Brain. Neurosci. Res. 2024; ahead of print. [Google Scholar] [CrossRef]
- Butt, T.H.; Tobiume, M.; Re, D.B.; Kariya, S. Physical Exercise Counteracts Aging-Associated White Matter Demyelination Causing Cognitive Decline. Aging Dis. 2024, 15, 2136–2148. [Google Scholar] [CrossRef] [PubMed]
- Augusto-Oliveira, M.; Arrifano, G.P.; Leal-Nazare, C.G.; Santos-Sacramento, L.; Lopes-Araujo, A.; Royes, L.F.F.; Crespo-Lopez, M.E. Exercise Reshapes the Brain: Molecular, Cellular, and Structural Changes Associated with Cognitive Improvements. Mol. Neurobiol. 2023, 60, 6950–6974. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, T. Preventive Strategies for Cognitive Decline and Dementia: Benefits of Aerobic Physical Activity, Especially Open-Skill Exercise. Brain Sci. 2023, 13, 521. [Google Scholar] [CrossRef] [PubMed]
- Lopez, D.C.; White, Z.J.; Hall, S.E. Anxiety in Alzheimer’s disease rats is independent of memory and impacted by genotype, age, sex, and exercise. Alzheimers Dement. 2024, 20, 3543–3550. [Google Scholar] [CrossRef] [PubMed]
- Mehla, J.; Deibel, S.H.; Karem, H.; Hossain, S.; Lacoursiere, S.G.; Sutherland, R.J.; Mohajerani, M.H.; Mcdonald, R.J. Dramatic impacts on brain pathology, anxiety, and cognitive function in the knock-in APP(NL-G-F) mouse model of Alzheimer disease following long-term voluntary exercise. Alzheimers Res. Ther. 2022, 14, 143. [Google Scholar] [CrossRef]
- Tieland, M.; Trouwborst, I.; Clark, B.C. Skeletal muscle performance and ageing. J. Cachexia Sarcopenia Muscle 2018, 9, 3–19. [Google Scholar] [CrossRef]
- Kerr, N.R.; Mossman, C.W.; Chou, C.H.; Bunten, J.M.; Kelty, T.J.; Childs, T.E.; Rector, R.S.; Arnold, W.D.; Grisanti, L.A.; Du, X.; et al. Hindlimb immobilization induces insulin resistance and elevates mitochondrial ROS production in the hippocampus of female rats. J. Appl. Physiol. 2024, 137, 512–526. [Google Scholar] [CrossRef]
- Li, F.; Bian, D.; Bai, T.; Jin, H.; Sun, X.; Lu, J.; Zhu, L.; Wu, W.; Zhou, J.; Shi, Y.; et al. Cognitive impairment is associated with sarcopenia mainly related to attention and calculation in hospitalized Chinese elderly men. Asia Pac. J. Clin. Nutr. 2022, 31, 534–542. [Google Scholar]
- Wu, B.; Lyu, Y.B.; Cao, Z.J.; Wei, Y.; Shi, W.Y.; Gao, X.; Zhou, J.H.; Kraus, V.B.; Zhao, F.; Chen, X.; et al. Associations of Sarcopenia, Handgrip Strength and Calf Circumference with Cognitive Impairment among Chinese Older Adults. Biomed. Environ. Sci. 2021, 34, 859–870. [Google Scholar]
- Fang, W.Y.; Tseng, Y.T.; Lee, T.Y.; Fu, Y.C.; Chang, W.H.; Lo, W.W.; Lin, C.L.; Lo, Y.C. Triptolide prevents LPS-induced skeletal muscle atrophy via inhibiting NF-kappaB/TNF-alpha and regulating protein synthesis/degradation pathway. Br. J. Pharmacol. 2021, 178, 2998–3016. [Google Scholar] [CrossRef]
- O’neill, B.T.; Lee, K.Y.; Klaus, K.; Softic, S.; Krumpoch, M.T.; Fentz, J.; Stanford, K.I.; Robinson, M.M.; Cai, W.; Kleinridders, A.; et al. Insulin and IGF-1 receptors regulate FoxO-mediated signaling in muscle proteostasis. J. Clin. Investig. 2016, 126, 3433–3446. [Google Scholar] [CrossRef] [PubMed]
- Di Ludovico, A.; La Bella, S.; Ciarelli, F.; Chiarelli, F.; Breda, L.; Mohn, A. Skeletal muscle as a pro- and anti-inflammatory tissue: Insights from children to adults and ultrasound findings. J. Ultrasound, 2024; ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, D.; Cao, G.; Shi, Q.; Zhu, J.; Zhang, M.; Cheng, H.; Wen, Q.; Xu, H.; Zhu, L.; et al. IL-27 signalling promotes adipocyte thermogenesis and energy expenditure. Nature 2021, 600, 314–318. [Google Scholar] [CrossRef]
- De La Monte, S.M.; Wands, J.R. Alzheimer’s disease is type 3 diabetes-evidence reviewed. J. Diabetes Sci. Technol. 2008, 2, 1101–1113. [Google Scholar] [CrossRef]
- Lopez-Cerdan, A.; Andreu, Z.; Hidalgo, M.R.; Soler-Saez, I.; De La Iglesia-Vaya, M.; Mikozami, A.; Guerini, F.R.; Garcia-Garcia, F. An integrated approach to identifying sex-specific genes, transcription factors, and pathways relevant to Alzheimer’s disease. Neurobiol. Dis. 2024, 199, 106605. [Google Scholar] [CrossRef] [PubMed]
- Corbett, B.F.; You, J.C.; Zhang, X.; Pyfer, M.S.; Tosi, U.; Iascone, D.M.; Petrof, I.; Hazra, A.; Fu, C.H.; Stephens, G.S.; et al. DeltaFosB Regulates Gene Expression and Cognitive Dysfunction in a Mouse Model of Alzheimer’s Disease. Cell Rep. 2017, 20, 344–355. [Google Scholar] [CrossRef]
- Qin, X.; Wang, Y.; Paudel, H.K. Inhibition of Early Growth Response 1 in the Hippocampus Alleviates Neuropathology and Improves Cognition in an Alzheimer Model with Plaques and Tangles. Am. J. Pathol. 2017, 187, 1828–1847. [Google Scholar] [CrossRef]
- Nagase, T.; Tohda, C. Skeletal muscle atrophy-induced hemopexin accelerates onset of cognitive impairment in Alzheimer’s disease. J. Cachexia Sarcopenia Muscle 2021, 12, 2199–2210. [Google Scholar] [CrossRef]
- Vegeto, E.; Villa, A.; Della Torre, S.; Crippa, V.; Rusmini, P.; Cristofani, R.; Galbiati, M.; Maggi, A.; Poletti, A. The Role of Sex and Sex Hormones in Neurodegenerative Diseases. Endocr. Rev. 2020, 41, 273–319. [Google Scholar] [CrossRef]
- Zalewska, T.; Pawelec, P.; Ziabska, K.; Ziemka-Nalecz, M. Sexual Dimorphism in Neurodegenerative Diseases and in Brain Ischemia. Biomolecules 2022, 13, 26. [Google Scholar] [CrossRef]
- Yang, J.; Yuan, S.; Jian, Y.; Lei, Y.; Hu, Z.; Yang, Q.; Yan, X.; Zheng, L.; Li, J.; Liu, W. Aerobic exercise regulates GPR81 signal pathway and mediates complement- microglia axis homeostasis on synaptic protection in the early stage of Alzheimer’s disease. Life Sci. 2023, 331, 122042. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Yang, J.; Jian, Y.; Lei, Y.; Yao, S.; Hu, Z.; Liu, X.; Tang, C.; Liu, W. Treadmill Exercise Modulates Intestinal Microbes and Suppresses LPS Displacement to Alleviate Neuroinflammation in the Brains of APP/PS1 Mice. Nutrients 2022, 14, 4134. [Google Scholar] [CrossRef] [PubMed]
- Jian, Y.; Yuan, S.; Yang, J.; Lei, Y.; Li, X.; Liu, W. Aerobic Exercise Alleviates Abnormal Autophagy in Brain Cells of APP/PS1 Mice by Upregulating AdipoR1 Levels. Int. J. Mol. Sci. 2022, 23, 9921. [Google Scholar] [CrossRef] [PubMed]
- Burch, N.; Arnold, A.S.; Item, F.; Summermatter, S.; Brochmann Santana Santos, G.; Christe, M.; Boutellier, U.; Toigo, M.; Handschin, C. Electric pulse stimulation of cultured murine muscle cells reproduces gene expression changes of trained mouse muscle. PLoS ONE 2010, 5, e10970. [Google Scholar] [CrossRef]
- Xiong, Y.; Wu, Z.; Zhang, B.; Wang, C.; Mao, F.; Liu, X.; Hu, K.; Sun, X.; Jin, W.; Kuang, S. Fndc5 loss-of-function attenuates exercise-induced browning of white adipose tissue in mice. FASEB J. 2019, 33, 5876–5886. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| IGF-1 | ACCTGCCTGGGTGTCCAAAT | CGATAGGGACGGGGACTTCT |
| PI3K | CGAAACAAAGCCGAGAACC | GCAATGTTTGACTTCGCCAT |
| Akt | CTTCTATGGTGCGGAGATTGT | ACAGCCCGAAGTCCGTTATCT |
| mTOR | TTCCTGAACAGCGAGCACAA | TGCCAAGACACAGTAGCGGA |
| Fbxo25 | AAGGTGTGACCCCTGTAGC | CCTCTTTTTGGCTGCGTATTCA |
| Fbxo32 | CAGCTTCGTGAGCGACCTC | GGCAGTCGAGAAGTCCAGTC |
| MAFbx | CCACTTCTCAGAGCGGCAGA | CTTCTTGGGTAACATCGCACA |
| Murf | CCACTTCTCAGAGCGGCAGA | CTTCTTGGGTAACATCGCACA |
| lc3a | GACCGCTGTAAGGAGGTGC | CTTGACCAACTCGCTCATGTTA |
| At9a | GTTCGCCCCCTTTAATAGTGC | TGAACTCCAACGTCAAGCGG |
| P62 | ATGTGAACATGGAGGGAAGA | GGAGTTCACCTGTAGATGGGT |
| Beclin | GAACTCTGGAGGTCTCGCT | CACCCAGGCTCGTTCTACC |
| GAPDH | CATGGCCTTCCGTGTTCCTA | CCTGCTTCACCACCTTCTTGAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Wu, X.; Wu, W.; Tang, Y.; Meng, X.; Peng, M.; Tang, C.; Zheng, L.; Liu, W. The Potential Related Genes and Mechanisms Involved in Improving the Treadmill Exercise Ability of APP/PS1 Mice. Int. J. Mol. Sci. 2024, 25, 10244. https://doi.org/10.3390/ijms251910244
Zhao Z, Wu X, Wu W, Tang Y, Meng X, Peng M, Tang C, Zheng L, Liu W. The Potential Related Genes and Mechanisms Involved in Improving the Treadmill Exercise Ability of APP/PS1 Mice. International Journal of Molecular Sciences. 2024; 25(19):10244. https://doi.org/10.3390/ijms251910244
Chicago/Turabian StyleZhao, Zhe, Xingqing Wu, Weijia Wu, Yingzhe Tang, Xiangyuan Meng, Mei Peng, Changfa Tang, Lan Zheng, and Wenfeng Liu. 2024. "The Potential Related Genes and Mechanisms Involved in Improving the Treadmill Exercise Ability of APP/PS1 Mice" International Journal of Molecular Sciences 25, no. 19: 10244. https://doi.org/10.3390/ijms251910244