
Methamphetamine-Induced Blood Pressure Sensitization Correlates with Morphological Alterations within A1/C1 Catecholamine Neurons
 , , ,
, , ,  ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. METH-Induced SBP Sensitization
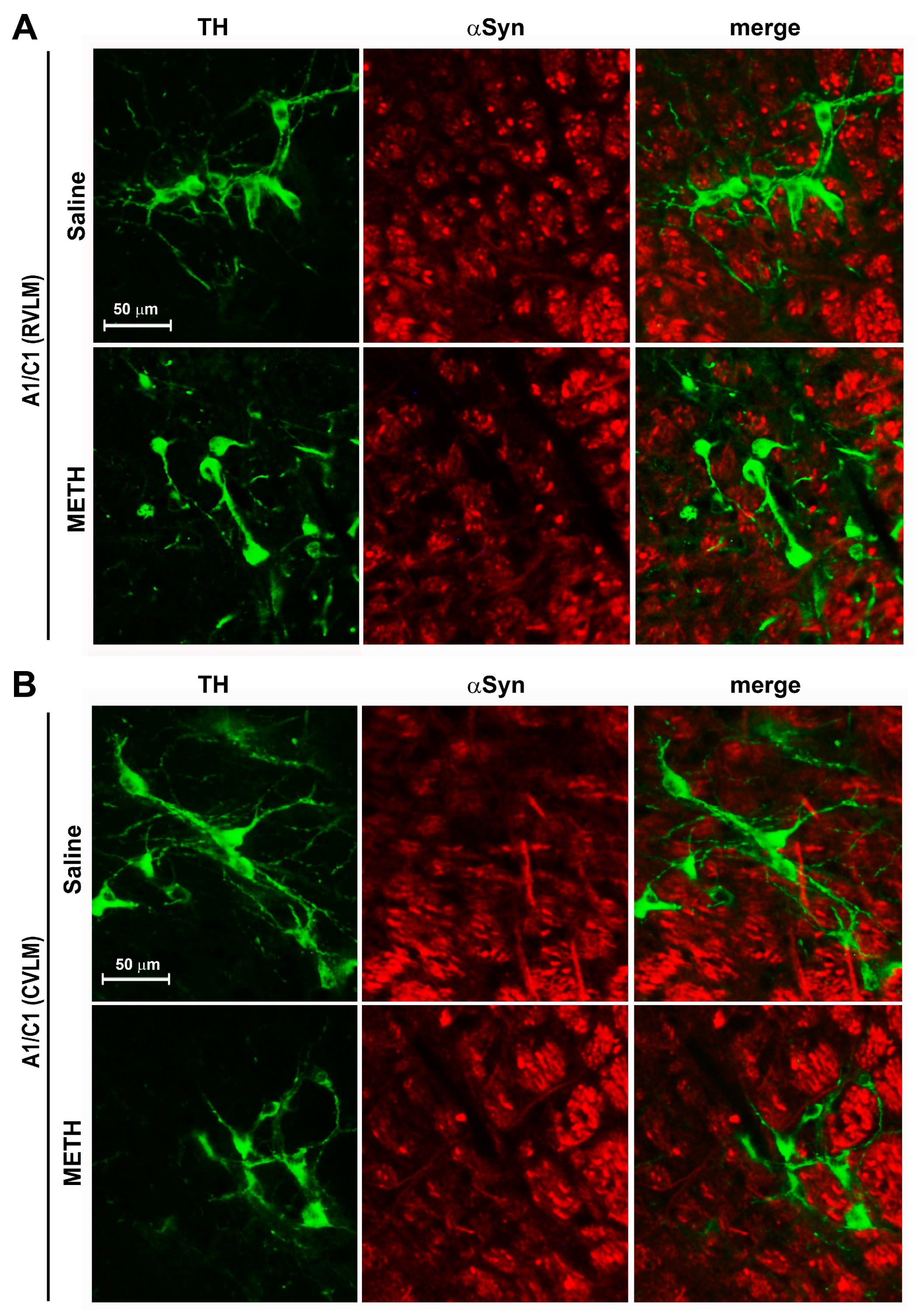
2.2. METH Sensitization Is Concomitant with Reduced Catecholamine Cell Density in A1/C1 of the Rostral and Caudal Ventrolateral Medulla (RVLM and CVLM)
2.3. METH Treatment Did Not Induce Neurodegeneration/Neurotoxicity within the RVLM and CVLM
2.4. METH Reduces Free Radicals within A1/C1 of the RVLM and CVLM
2.5. METH Increases Heat Shock Protein 70 (HSP70) within A1/C1 of Both the RVLM and CVLM
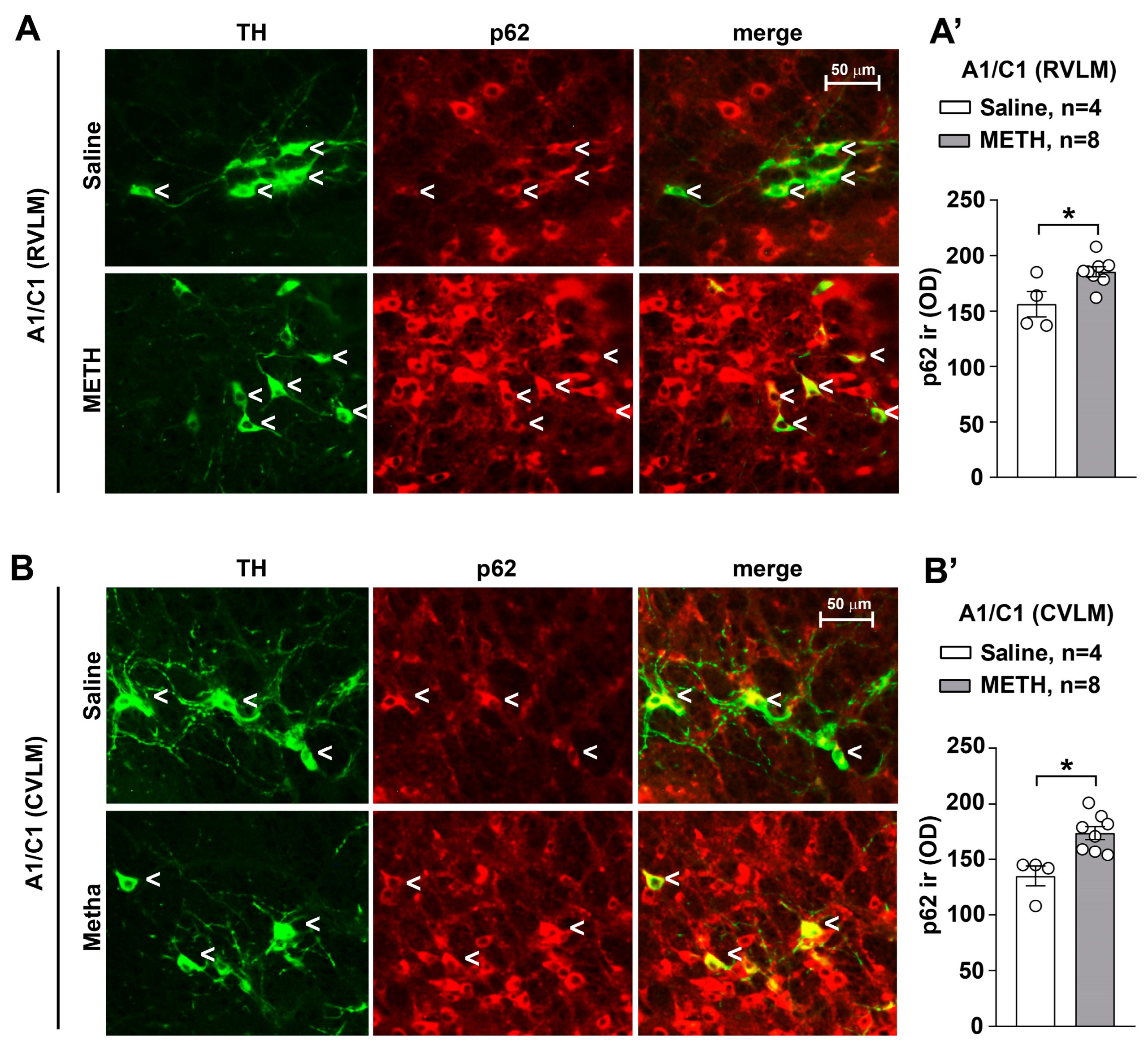
2.6. METH Increases Sequestosome/p62 within A1/C1 of Both the RVLM and CVLM
2.7. Phospho-cJun (p-cJun) Decreases Following a METH-Sensitization Protocol Leading to Increased Blood Pressure
3. Discussion
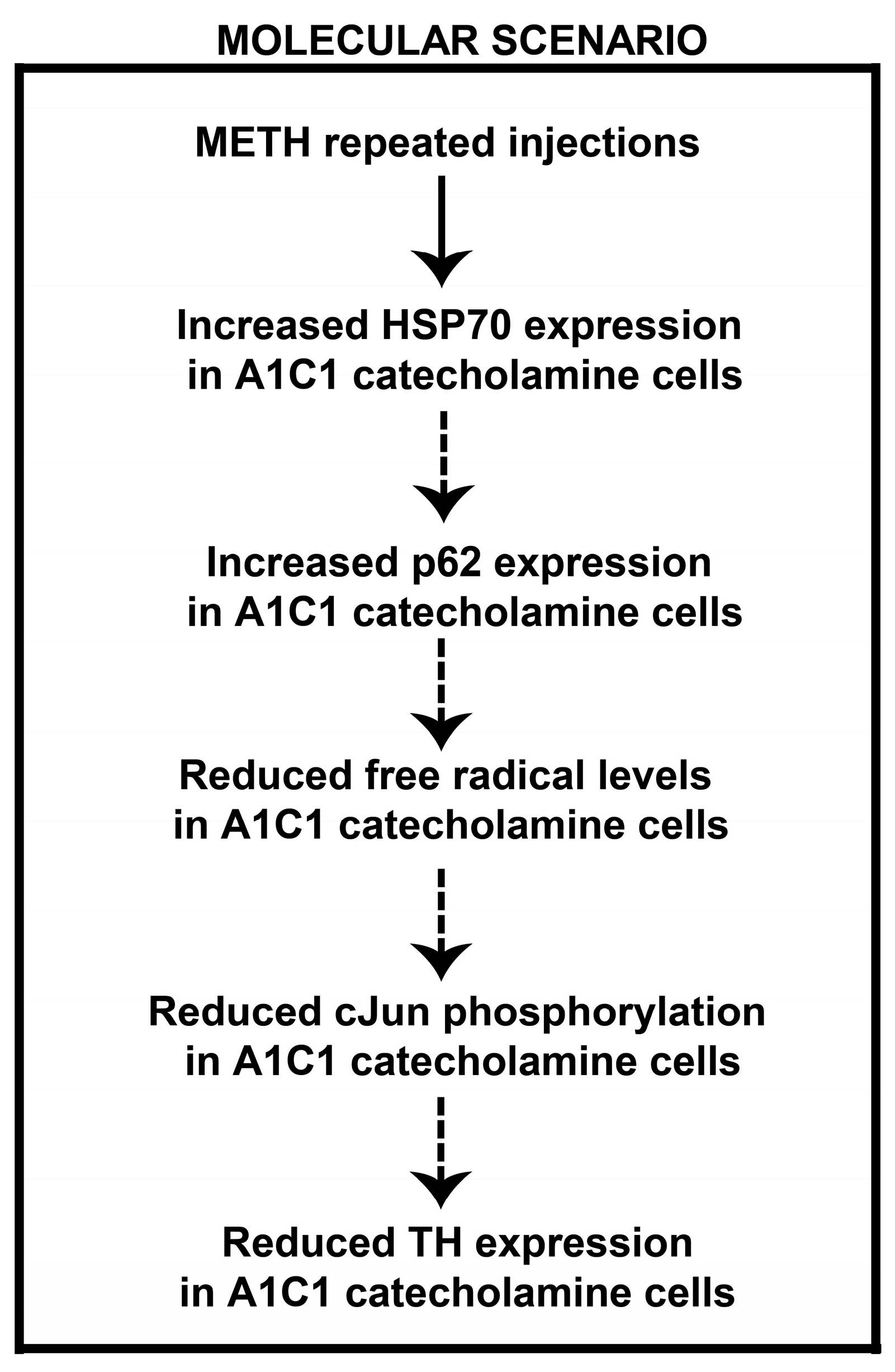
Appendix: A Working Hypothesis
4. Materials and Methods
4.1. Materials
4.2. Animals and Housing
4.3. Experimental Strategies
4.4. Blood Pressure Measurement
4.5. Vascular Reactivity
4.6. Immunohistochemical Analysis in Medullary Catecholamine Nuclei
4.7. TH-Positive Cell Counting in A1/C1, A2/C2 and AP Medullary Catecholamine Nuclei
4.8. Fluro-Jade B Staining in the RVLM/CVLM
4.9. Immunohistochemical Analysis in Mesenteric Arteries
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferrucci, M.; Pasquali, L.; Paparelli, A.; Ruggieri, S.; Fornai, F. Pathways of methamphetamine toxicity. Ann. N. Y. Acad. Sci. 2008, 1139, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Moszczynska, A.; Callan, S.P. Molecular, behavioral, and physiological consequences of methamphetamine neurotoxicity: Implications for treatment. J. Pharmacol. Exp. Therap. 2017, 362, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Xu, E.; Liu, J.; Liu, H.; Wang, X.; Xiong, H. Role of microglia in methamphetamine-induced neurotoxicity. Int. J. Physiol. Pathophysiol. Pharmacol. 2017, 9, 84–100. [Google Scholar] [PubMed]
- Limanaqi, F.; Busceti, C.L.; Celli, R.; Biagioni, F.; Fornai, F. Autophagy as a gateway for the effects of methamphetamine: From neurotransmitter release and synaptic plasticity to psychiatric and neurodegenerative disorders. Prog. Neurobiol. 2021, 204, 102112. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Lu, J.; Chen, X.; Gao, Z.; Zhang, C.; Chen, C.; Qiao, D.; Wang, H. Methamphetamine exposure induces neuronal programmed necrosis by activating the receptor-interacting protein kinase 3 -related signalling pathway. FASEB J. 2021, 35, e21561. [Google Scholar] [CrossRef]
- Lenzi, P.; Biagioni, F.; Busceti, C.L.; Lazzeri, G.; Polzella, M.; Frati, A.; Ferrucci, M.; Fornai, F. Alterations of Mitochondrial Structure in Methamphetamine Toxicity. Int. J. Mol. Sci. 2022, 23, 8926. [Google Scholar] [CrossRef]
- Wagner, G.C.; Ricaurte, G.A.; Seiden, L.S.; Schuster, C.R.; Miller, R.J.; Westley, J. Long-lasting depletions of striatal dopamine and loss of dopamine uptake sites following repeated administration of methamphetamine. Brain Res. 1980, 181, 151–160. [Google Scholar] [CrossRef]
- Ricaurte, G.A.; Guillery, R.W.; Seiden, L.S.; Schuster, C.R.; Moore, R.Y. Dopamine nerve terminal degeneration produced by high doses of methylamphetamine in the rat brain. Brain Res. 1982, 235, 93–103. [Google Scholar] [CrossRef]
- Fornai, F.; Lenzi, P.; Gesi, M.; Soldani, P.; Ferrucci, M.; Lazzeri, G.; Capobianco, L.; Battaglia, G.; De Blasi, A.; Nicoletti, F.; et al. Methamphetamine produces neuronal inclusions in the nigrostriatal system and in PC12 cells. J. Neurochem. 2004, 88, 114–123. [Google Scholar] [CrossRef]
- Moratalla, R.; Khairnar, A.; Simola, N.; Granado, N.; García-Montes, J.R.; Porceddu, P.F.; Tizabi, Y.; Costa, G.; Morelli, M. Amphetamine-related drugs neurotoxicity in humans and in experimental animals: Main mechanisms. Prog. Neurobiolog. 2017, 155, 149–170. [Google Scholar] [CrossRef]
- Ferrucci, M.; Biagioni, F.; Busceti, C.L.; Vidoni, C.; Castino, R.; Isidoro, C.; Ryskalin, L.; Frati, A.; Puglisi-Allegra, S.; Fornai, F. Inhibition of autophagy in vivo extends methamphetamine toxicity to mesencephalic cell bodies. Pharmaceuticals 2021, 14, 1003. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, G.; Gesi, M.; Lenzi, P.; Busceti, C.L.; Soldani, P.; Orzi, F.; Rampello, L.; Nicoletti, F.; Ruggieri, S.; Fornai, F. Morphological and biochemical evidence that apomorphine rescues striatal dopamine terminals and prevents methamphetamine toxicity. Ann. N. Y. Acad Sci. 2002, 965, 254–266. [Google Scholar] [CrossRef]
- Battaglia, G.; Fornai, F.; Busceti, C.L.; Aloisi, G.; Cerrito, F.; De Blasi, A.; Melchiorri, D.; Nicoletti, F. Selective blockade of mGlu5 metabotropic glutamate receptors is protective against methamphetamine neurotoxicity. J. Neurosci. 2002, 22, 2135–2141. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.F.; Wearne, T.A.; Cornish, J.L.; Goodchild, A.K. Effects of acute and chronic systemic methamphetamine on respiratory, cardiovascular and metabolic function, and cardiorespiratory reflexes. J. Physiol. 2016, 594, 763–780. [Google Scholar] [CrossRef]
- Busceti, C.L.; Bucci, D.; De Lucia, M.; Ferrucci, M.; Scioli, M.; Carrizzo, A.; Nicoletti, F.; Vecchione, C.; Fornai, F. Different dose of methamphetamine are needed to produce locomotor or blood pressure sensitization in mice. Life 2024, 14, 723. [Google Scholar] [CrossRef] [PubMed]
- Vickers, M.L.; Chan, H.Y.; Elliott, S.; Ketheesan, S.; Ramineni, V.; Eriksson, L.; McMahon, K.; Oddy, B.; Scott, J.G. Stimulant medications in the management of bulimia nervosa and anorexia nervosa in patients with and without comorbid attention deficithyper-activity disorder: A systematic review. Eat Behav. 2024, 54, 101908. [Google Scholar] [CrossRef] [PubMed]
- Guyenet, P.G.; Stornetta, R.L.; Bochorishvili, G.; Depuy, S.D.; Burke, P.G.; Abbott, S.B. C1 neurons: The body’s EMTs. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R187–R204. [Google Scholar] [CrossRef] [PubMed]
- Biagioni, F.; Ferese, R.; Limanaqi, F.; Madonna, M.; Lenzi, P.; Gambardella, S.; Fornai, F. Methamphetamine persistently increases alpha-synuclein and suppresses gene promoter methylation within striatal neurons. Brain Res. 2019, 1719, 157–175. [Google Scholar] [CrossRef]
- Motohashi, H.; Yamamoto, M. Nrf2-Keap1 defines a physiologically important stress response mechanism. Trends Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef]
- Hayes, J.D.; McMahon, M. NRF2 and KEAP1 mutations: Permanent activation of an adaptive response in cancer. Trends Biochem. Sci. 2009, 34, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef]
- Fornai, F.; Lenzi, P.; Ferrucci, M.; Lazzeri, G.; di Poggio, A.B.; Natale, G.; Busceti, C.L.; Biagioni, F.; Giusiani, M.; Ruggieri, S.; et al. Occurrence of neuronal inclusions combined with increased nigral expression of alpha-synuclein within dopa-minergic neurons following treatment with amphetamine derivatives in mice. Brain Res. Bull. 2005, 65, 405–413. [Google Scholar] [CrossRef]
- Schiffrin, E.L. Reactivity of small blood vessels in hypertension: Relation with structural changes. State of the art lecture. Hypertension 1992, 19, II1–II9. [Google Scholar] [CrossRef]
- Naito, Y.; Yoshida, H.; Konishi, C.; Ohara, N. Differences in responses to norepinephrine and adenosine triphosphate in isolated perfused mesenteric vascular beds between normotensive and spontaneously hypertensive rats. J. Cardiovasc. Pharmacol. 1998, 32, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Tatchum-Talom, R.; Eyster, K.M.; Martin, D.S. Sexual dimorphism in angiotensin II-induced hypertension and vascular alterations. Can. J. Physiol. Pharmacol. 2005, 83, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.A.; Ruggiero, D.A.; Joh, T.H.; Park, D.H.; Reis, D.J. Rostral ventrolateral medulla: Selective projections to the thoracic autonomic cell column from the region containing C1 adrenaline neurons. J. Comp. Neurol. 1984, 228, 168–185. [Google Scholar] [CrossRef]
- Abbott, S.B.; Stornetta, R.L.; Socolovsky, C.S.; West, G.H.; Guyenet, P.G. Photostimulation of channelrhodopsin-2 expressing ventrolateral medullary neurons increases sympathetic nerve activity and blood pressure in rats. J. Physiol. 2009, 587, 5613–5631. [Google Scholar] [CrossRef]
- Marina, N.; Abdala, A.P.; Korsak, A.; Simms, A.E.; Allen, A.M.; Paton, J.F.; Gourine, A.V. Control of sympathetic vasomotor tone by catecholaminergic C1 neurones of the rostral ventrolateral medulla oblongata. Cardiovasc. Res. 2011, 91, 703–710. [Google Scholar] [CrossRef]
- Standish, A.; Enquist, L.W.; Escardo, J.A.; Schwaber, J.S. Central neuronal circuit innervating the rat heart defined by transneuronal transport of pseudorabies virus. J. Neurosci. 1995, 15, 1998–2012. [Google Scholar] [CrossRef]
- Rogers, R.C.; Hermann, G.E.; Travagli, R.A. Brainstem pathways responsible for oesophageal control of gastric motility and tone in the rat. J. Physiol. 1999, 514, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Card, J.P.; Sved, J.C.; Craig, B.; Raizada, M.; Vazquez, J.; Sved, A.F. Efferent projections of rat rostroventrolateral medulla C1 catecholamine neurons: Implications for the central control of cardiovascular regulation. J. Comp. Neurol. 2006, 499, 840–859. [Google Scholar] [CrossRef] [PubMed]
- Olofsson, P.S.; Rosas-Ballina, M.; Levine, Y.A.; Tracey, K.J. Rethinking inflammation: Neural circuits in the regulation of immunity. Immunol. Rev. 2012, 248, 188–204. [Google Scholar] [CrossRef] [PubMed]
- DePuy, S.D.; Stornetta, R.L.; Bochorishvili, G.; Deisseroth, K.; Witten, I.; Coates, M.; Guyenet, P.G. Glutamatergic Neurotransmission between the C1 Neurons and the Parasympathetic Preganglionic Neurons of the Dorsal Motor Nucleus of the Vagus. J. Neurosci. 2013, 33, 1486–1497. [Google Scholar] [CrossRef]
- Stornetta, R.L.; Sevigny, C.P.; Guyenet, P.G. Vesicular glutamate transporter DNPI/VGLUT2 mRNA is present in C1 and several other groups of brainstem catecholaminergic neurons. J. Comp. Neurol. 2002, 444, 191–206. [Google Scholar] [CrossRef]
- Morrison, S.F.; Ernsberger, P.; Milner, T.A.; Callaway, J.; Gong, A.; Reis, D.J. A glutamate mechanism in the intermediolateral nucleus mediates sympathoexcitatory responses to stimulation of the rostral ventrolateral medulla. Prog. Brain Res. 1989, 81, 159–169. [Google Scholar] [CrossRef]
- Abbott, S.B.; Kanbar, R.; Bochorishvili, G.; Coates, M.B.; Stornetta, R.L.; Guyenet, P.G. C1 neurons excite locus coeruleus and A5 noradrenergic neurons along with sympathetic outflow in rats. J. Physiol. 2012, 590, 2897–2915. [Google Scholar] [CrossRef]
- Lorang, D.; Amara, S.G.; Simerly, R.B. Cell-type-specific expression of catecholamine transporters in the rat brain. J. Neurosci. 1994, 14, 4903–4914. [Google Scholar] [CrossRef]
- Sved, A.F. PNMT-containing catecholaminergic neurons are not necessarily adrenergic. Brain Res. 1989, 481, 113–118. [Google Scholar] [CrossRef]
- Armstrong, D.M.; Ross, C.A.; Pickel, V.M.; Joh, T.H.; Reis, D.J. Distribution of dopamine-, noradrenaline-, and adrenaline- containing cell bodies in the rat medulla oblongata: Demonstrated by the immunocytochemical localization of catecholamine biosynthetic enzymes. J. Comp. Neurol. 1982, 212, 173–187. [Google Scholar] [CrossRef]
- Sevigny, C.P.; Bassi, J.; Teschemacher, A.G.; Kim, K.S.; Williams, D.A.; Anderson, C.R.; Allen, A.M. C1 neurons in the rat rostral ventrolateral medulla differentially express vesicular monoamine transporter 2 in soma and axonal compartments. Eur. J. Neurosci. 2008, 28, 1536–1544. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.W.; Bayliss, D.A.; Guyenet, P.G. C1 neurons of neonatal rats: Intrinsic beating properties and 2-adrenergic receptors. Am. J. Physiol. 1995, 269, R1356–R1369. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.W.; Guyenet, P.G.; Bayliss, D.A. Voltage-dependent calcium currents in bulbospinal neurons of neonatal rat rostral ventrolateral medulla: Modulation by alpha2-adrenergic receptors. J. Neurophysiol. 1998, 79, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Engler, K.L.; Rudd, M.L.; Ryan, J.J.; Stewart, J.K.; Fischer-Stenger, K. Autocrine actions of macrophage-derived catecholamines on interleukin-1 beta 5. J. Neuroimmunol. 2005, 160, 87–91. [Google Scholar] [CrossRef]
- Sorriento, D.; Santulli, G.; Del Giudice, C.; Anastasio, A.; Trimarco, B.; Iaccarino, G. Endothelial cells are able to synthesize and release catecholamines both in vitro and in vivo. Hypertension 2012, 60, 129–136. [Google Scholar] [CrossRef]
- Chan, R.K.; Sawchenko, P.E. Organization and transmitter specificity of medullary neurons activated by sustained hypertension: Implications for understanding baroreceptor reflex circuitry. J. Neurosci. 1998, 18, 371–387. [Google Scholar] [CrossRef]
- Schreihofer, A.M.; Guyenet, P.G. The baroreflex and beyond: Control of sympathetic vasomotor tone by GABAergic neurons in the ventrolateral medulla. Clin. Exp. Pharmacol. Physiol. 2002, 29, 514–521. [Google Scholar] [CrossRef]
- Tucker, D.C.; Saper, C.B.; Ruggiero, D.A.; Reis, D.J. Organization of central adrenergic pathways. I. Relationships of ventrolateral medullary projections to the hypothalamus and spinal cord. J. Comp. Neurol. 1987, 259, 591–603. [Google Scholar] [CrossRef]
- Ruggiero, D.A.; Cravo, S.L.; Golanov, E.; Gomez, R.; Anwar, M.; Reis, D.J. Adrenergic and non-adrenergic spinal projections of a cardiovascularactive pressor area of medulla oblongata: Quantitative topographic analysis. Brain Res. 1994, 663, 107–120. [Google Scholar] [CrossRef]
- Otake, K.; Ruggiero, D.A.; Nakamura, Y. Adrenergic innervation of forebrain neurons that project to the paraventricular thalamic nucleus in the rat. Brain Res. 1995, 697, 17–26. [Google Scholar] [CrossRef]
- Minson, J.B.; Llewellyn-Smith, I.J.; Chalmers, J.P.; Pilowsky, P.M.; Arnolda, L.F. c-fos identifies GABA-synthesizing barosensitive neurons in caudal ventrolateral medulla. Neuroreport 1997, 8, 3015–3021. [Google Scholar] [CrossRef] [PubMed]
- Verberne, A.J.M.; Stornetta, R.L.; Guyenet, P.G. Properties of C1 and other ventrolateral medullary neurones with hypothalamic projections in the rat. J. Physiol. 1999, 517, 477–494. [Google Scholar] [CrossRef]
- Day, T.A.; Ferguson, A.V.; Renaud, L.P. Facilitatory influence of noradrenergic afferents on the excitability of rat paraventricular nucleus neurosecretory cells. J. Physiol. 1984, 355, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Kannan, H.; Yamashita, H.; Osaka, T. Paraventricular neurosecretory neurons: Synaptic inputs from the ventrolateral medulla in rats. Neurosci. Lett. 1984, 51, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Day, T.A.; Sibbald, J.R. Solitary nucleus excitation of supraoptic vasopressin cells via adrenergic afferents. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1988, 254, R711–R716. [Google Scholar] [CrossRef]
- Day, T.A.; Sibbald, J.R. A1 cell group mediates solitary nucleus excitation of supraoptic vasopressin cells. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1989, 257, R1020–R1026. [Google Scholar] [CrossRef]
- Tanaka, J.; Nishimura, J.; Kimura, F.; Nomura, M. Noradrenergic excitatory inputs to median preoptic neurones in rats. Neuroreport 1992, 3, 946–948. [Google Scholar] [CrossRef]
- Saphier, D. Electrophysiology and neuropharmacology of noradrenergic projections to rat PVN magnocellular neurons. Am. J. Physiol. 1993, 264, R891–R902. [Google Scholar] [CrossRef]
- Tanaka, M.; DeKloet, E.R.; De Wied, D.; Versteeg, D.H.G. Arginine8 vasopressin affects catecholamine metabolism in specific brain nuclei. Life Sci. 1997, 20, 1799. [Google Scholar] [CrossRef]
- Blessing, W.W.; Sved, A.F.; Reis, D.J. Destruction of noradrenergic neurons in rabbit brainstem elevates plasma vasopressin, causing hypertension. Science 1982, 217, 661–663. [Google Scholar] [CrossRef]
- Saper, C.B.; Reis, D.J.; Joh, T. Medullary catecholamine inputs to the anteroventral third ventricular cardiovascular regulatory region in the rat. Neurosci. Lett. 1983, 42, 285–291. [Google Scholar] [CrossRef]
- Ciriello, J.; Caverson, M. Direct pathway from neurons in the ventrolateral medulla relaying cardiovascular afferent information to the supraoptic nucleos in the cat. Brain Res. Rev. 1984, 292, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Chiba, T.; Murata, Y. Afferent and efferent connections of the medial preoptic area in the rat: A WGA-HRP study. Brain Res. Bull. 1985, 14, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Galaz, C.; Dyer, R.G.; Herbison, A.E. Analysis of brainstem A1 and A2 noradrenergic inputs to the preoptic area using microdialysis in the rat. Brain Res. 1994, 636, 227–232. [Google Scholar] [CrossRef]
- Pedrino, G.R.; Maurino, I.; de Almeida Colombari, D.S.; Cravo, S.L. Role of catecholaminergic neurones of the caudal ventrolateral medulla in cardiovascular responses induced by acute changes in circulating volume in rats. Exp. Physiol. 2006, 91, 995–1005. [Google Scholar] [CrossRef]
- Colombari, D.S.; Pedrino, G.R.; Freiria-Oliveira, A.H.; Korim, W.S.; Maurino, I.C.; Cravo, S.L. Lesions of medullary catecholaminergic neurons increase salt intake in rats. Brain Res. Bull. 2008, 76, 572–578. [Google Scholar] [CrossRef]
- Papas, S.; Ferguson, A.V. Electrophysiological evidence of baroreceptor input to area postrema. Am. J. Physiol. 1991, 261, R9–R13. [Google Scholar] [CrossRef]
- Gerstberger, R.; Fahrenholz, F. Autoradiographic localization of V1 vasopressin binding sites in rat brain and kidney. Eur. J. Pharmacol. 1989, 167, 105–116. [Google Scholar] [CrossRef]
- Lenkei, Z.; Palkovits, M.; Corvol, P.; Llorens-Cortes, C. Expression of angiotensin type-1 (AT1) and type-2 (AT2) receptor mRNAs in the adult rat brain: A functional neuroanatomical review. Front. Neuroendocrinol. 1997, 18, 383–439. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Hara, Y.; Anrather, J.; Speth, R.C.; Iadecola, C.; Pickel, V.M. Angiotensin II subtype 1A (AT1A) receptors in the rat sensory vagal complex: Subcellular localization and association with endogenous angiotensin. Neuroscience 2003, 122, 21–36. [Google Scholar] [CrossRef]
- Cai, Y.; Hay, M.; Bishop, V.S. Stimulation of area postrema by vasopressin and angiotensin II molulates neuronal activity in the nucleus tractus solitarius. Brain Res. 1994, 647, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Bishop, V.S.; Sanderford, M.G. Angiotensin II modulation of the arterial baroreflex: Role of the area postrema. Clin. Exp. Pharmacol. Physiol. 2000, 27, 428–431. [Google Scholar] [CrossRef] [PubMed]
- Cox, B.F.; Hay, M.; Bishop, V.S. Neurons in area postrema mediate vasopressin-induced enhancement of the baroreflex. Am. J. Physiol. 1990, 258, H1943–H1946. [Google Scholar] [CrossRef]
- Xue, B.; Gole, H.; Pamidimukkala, J.; Hay, M. Role of the area postrema in angiotensin II modulation of baroreflex controlof heart rate in conscious mice. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H1003–H1007. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, A.V.; Bains, J.S. Electrophysiology of the circumventricular organs. Front. Neuroendocrinol. 1996, 17, 440–475. [Google Scholar] [CrossRef]
- Smith, P.M.; Lowes, V.L.; Ferguson, A.V. Circulating vasopressin influences area postrema neurons. Neuroscience 1994, 59, 185–194. [Google Scholar] [CrossRef]
- Braga, V.A.; Medeiros, I.A.; Ribeiro, T.P.; França-Silva, M.S.; Botelho-Ono, M.S.; Guimarães, D.D. Angiotensin-II-induced reactive oxygen species along the SFO-PVN-RVLM pathway: Implications in neurogenic hypertension. Braz. J. Med. Biol. Res. 2011, 44, 871–876. [Google Scholar] [CrossRef]
- Bjørkøy, G.; Lamark, T.; Brech, A.; Outzen, H.; Perander, M.; Overvatn, A.; Stenmark, H.; Johansen, T. p6 2/SQSTM1 forms protein aggregates degraded by autophagy and has a protective effect on huntingtin-induced cell death. J. Cell Biol. 2005, 171, 603–614. [Google Scholar] [CrossRef]
- Komatsu, M.; Waguri, S.; Koike, M.; Sou, Y.S.; Ueno, T.; Hara, T.; Mizushima, N.; Iwata, J.; Ezaki, J.; Murata, S.; et al. Homeostatic levels of p62 control cytoplasmic inclusion body formation in autophagy-deficient mice. Cell 2007, 131, 1149–1163. [Google Scholar] [CrossRef]
- Pankiv, S.; Clausen, T.H.; Lamark, T.; Brech, A.; Bruun, J.A.; Outzen, H.; Øvervatn, A.; Bjørkøy, G.; Johansen, T. p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J. Biol. Chem. 2007, 282, 24131–24145. [Google Scholar] [CrossRef]
- Ichimura, Y.; Kumanomidou, T.; Sou, Y.S.; Mizushima, T.; Ezaki, J.; Ueno, T.; Kominami, E.; Yamane, T.; Tanaka, K.; Komatsu, M. p62/SQSTM1 functions as a signaling hub and an autophagy adaptor. J. Biol. Chem. 2008, 283, 22847–22857. [Google Scholar] [CrossRef] [PubMed]
- Lamark, T.; Kirkin, V.; Dikic, I.; Johansen, T. NBR1 and p62 as cargo receptors for selective autophagy of ubiquitinated targets. Cell Cycle 2009, 8, 1986–1990. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Lamark, T.; Sjøttem, E.; Larsen, K.B.; Awuh, J.A.; Øvervatn, A.; McMahon, M.; Hayes, J.D.; Johansen, T. p62/SQSTM1 is a target gene for transcription factor NRF2 and creates a positive feedback loop by inducing antioxidant response element-driven gene transcription. J. Biol. Chem. 2010, 285, 22576–22591. [Google Scholar] [CrossRef] [PubMed]
- Carrizzo, A.; Lenzi, P.; Procaccini, C.; Damato, A.; Biagioni, F.; Ambrosio, M.; Amodio, G.; Remondelli, P.; Del Giudice, C.; Izzo, R.; et al. Pentraxin 3 induces vascular endothelial dysfunction through a P-selectin/matrix metalloproteinase-1 pathway. Circulation 2015, 131, 1495–1505. [Google Scholar] [CrossRef]
- Schiattarella, G.G.; Cattaneo, F.; Carrizzo, A.; Paolillo, R.; Boccella, N.; Ambrosio, M.; Damato, A.; Pironti, G.; Franzone, A.; Russo, G.; et al. Akap1 regulates vascular function and endothelial cells behavior. Hypertension 2018, 71, 507–517. [Google Scholar] [CrossRef]
- Zacchigna, L.; Vecchione, C.; Notte, A.; Cordenonsi, M.; Dupont, S.; Maretto, S.; Cifelli, G.; Ferrari, A.; Maffei, A.; Fabbro, C.; et al. Emilin1 links TGF-beta maturation to blood pressure homeostasis. Cell 2006, 124, 929–942. [Google Scholar] [CrossRef]
- Schmued, L.C.; Albertson, C.; Slikker, W., Jr. Fluoro-Jade B: A novel fluorochrome for the sensitive and reliable histochemical localization of neuronal degeneration. Brain Res. 1997, 751, 37–46. [Google Scholar] [CrossRef]
- Schmued, L.C.; Hopkins, K.J. Fluoro-Jade B: A high affinity fluorescent marker for the localization of neuronal degeneration. Brain Res. 2000, 874, 123–130. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Busceti, C.L.; Bucci, D.; Damato, A.; De Lucia, M.; Venturini, E.; Ferrucci, M.; Lazzeri, G.; Puglisi-Allegra, S.; Scioli, M.; Carrizzo, A.; et al. Methamphetamine-Induced Blood Pressure Sensitization Correlates with Morphological Alterations within A1/C1 Catecholamine Neurons. Int. J. Mol. Sci. 2024, 25, 10282. https://doi.org/10.3390/ijms251910282
Busceti CL, Bucci D, Damato A, De Lucia M, Venturini E, Ferrucci M, Lazzeri G, Puglisi-Allegra S, Scioli M, Carrizzo A, et al. Methamphetamine-Induced Blood Pressure Sensitization Correlates with Morphological Alterations within A1/C1 Catecholamine Neurons. International Journal of Molecular Sciences. 2024; 25(19):10282. https://doi.org/10.3390/ijms251910282
Chicago/Turabian StyleBusceti, Carla Letizia, Domenico Bucci, Antonio Damato, Massimiliano De Lucia, Eleonora Venturini, Michela Ferrucci, Gloria Lazzeri, Stefano Puglisi-Allegra, Mariarosaria Scioli, Albino Carrizzo, and et al. 2024. "Methamphetamine-Induced Blood Pressure Sensitization Correlates with Morphological Alterations within A1/C1 Catecholamine Neurons" International Journal of Molecular Sciences 25, no. 19: 10282. https://doi.org/10.3390/ijms251910282
APA StyleBusceti, C. L., Bucci, D., Damato, A., De Lucia, M., Venturini, E., Ferrucci, M., Lazzeri, G., Puglisi-Allegra, S., Scioli, M., Carrizzo, A., Nicoletti, F., Vecchione, C., & Fornai, F. (2024). Methamphetamine-Induced Blood Pressure Sensitization Correlates with Morphological Alterations within A1/C1 Catecholamine Neurons. International Journal of Molecular Sciences, 25(19), 10282. https://doi.org/10.3390/ijms251910282