

Effects of Corm Treatment with Cold Plasma and Electromagnetic Field on Growth and Production of Saffron Metabolites in Crocus sativus

 , , , , ,
, , , , ,  , , and
, , and 
Abstract
1. Introduction
2. Results
2.1. Effects on Sprouting Kinetics, Seedling Growth, and Number of Leaf Trichomes
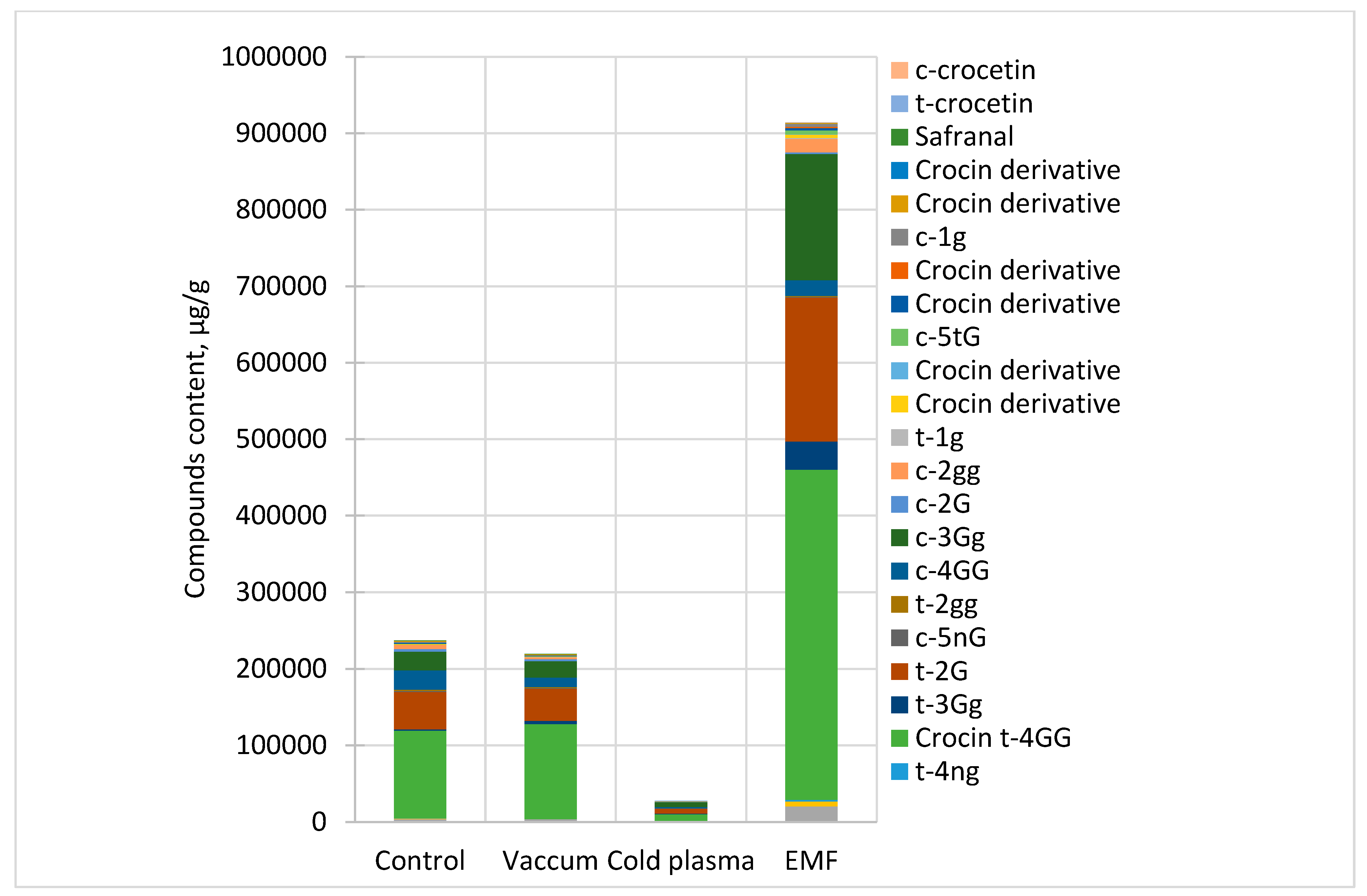
2.2. Effects on Amounts of Secondary Metabolites in the Stigma of C. sativus
3. Discussion
4. Materials and Methods
4.1. Plant Material and Seedling Cultivation
4.2. Corm Treatment by CP, Vacuum, and EMF
4.3. Analysis of Sprouting and Plant Morphometric Parameters
4.4. Sample Preparation and HPLC-DAD Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Attri, P.; Ishikawa, K.; Okumura, T.; Koga, K.; Shiratani, M. Plasma agriculture from laboratory to farm: A review. Processes 2020, 8, 1002. [Google Scholar] [CrossRef]
- Adhikari, B.; Adhikari, M.; Park, G. The effects of plasma on plant growth, development, and sustainability. Appl. Sci. 2020, 10, 6045. [Google Scholar] [CrossRef]
- Waskow, A.; Howling, A.; Furno, I. Mechanisms of plasma-seed treatments as a potential seed processing technology. Front. Phys. 2021, 9, 617345. [Google Scholar] [CrossRef]
- Mildaziene, V.; Ivankov, A.; Sera, B.; Baniulis, D. Biochemical and Physiological Plant Processes Affected by Seed Treatment with Non-Thermal Plasma. Plants 2022, 11, 856. [Google Scholar] [CrossRef]
- Kaur, S.; Vian, A.; Chandel, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Sensitivity of plants to high frequency electromagnetic radiation: Cellular mechanisms and morphological changes. Rev. Environ. Sci. Biotechnol. 2021, 20, 55–74. [Google Scholar] [CrossRef]
- Bukhari, S.I.; Manzoor, M.; Dhar, M.K. A comprehensive review of the pharmacological potential of Crocus sativus and its bioactive apocarotenoids. Biomed. Pharmacother. 2018, 98, 733–745. [Google Scholar] [CrossRef]
- Mykhailenko, O.; Desenko, V.; Ivanauskas, L.; Georgiyants, V. Standard operating procedure of Ukrainian saffron cultivation according with Good Agricultural and Collection Practices to assure quality and traceability. Ind. Crops Prod. 2020, 151, 112376–112387. [Google Scholar] [CrossRef]
- Ashraf, N.; Jain, D.; Vishwakarma, R.A. Identification, cloning and characterization of an ultrapetala transcription factor CsULT1 from Crocus: A novel regulator of apocarotenoid biosynthesis. BMC Plant Biol. 2015, 15, 25. [Google Scholar] [CrossRef]
- Butnariu, M.; Quispe, C.; Herrera-Bravo, J.; Sharifi-Rad, J.; Singh, L.; Aborehab, N.M.; Bouyahya, A.; Venditti, A.; Sen, S.; Acharya, K.; et al. The pharmacological activities of Crocus sativus L.: A review based on the mechanisms and therapeutic opportunities of its phytoconstituents. Oxid. Med. Cell. Longev. 2022, 2022, 8214821. [Google Scholar] [CrossRef]
- ISO 3632-1:2011; Spices—Saffron (Crocus sativus L.). Specification (Edition 2, 2011). International Organization for Standardization: Genève, Switzerland, 2011; 6p.
- Brinckmann, J.A. Geographical indications for medicinal plants: Globalization, climate change, quality and market implications for geo-authentic botanicals. World J. Tradit. Chin. Med. 2015, 1, 16–23. [Google Scholar] [CrossRef]
- D’Archivio, A.A.; Giannitto, A.; Maggi, M.A.; Ruggieri, F. Geographical classification of Italian saffron (Crocus sativus L.) based on chemical constituents determined by high-performance liquid-chromatography and by using linear discriminant analysis. Food Chem. 2016, 212, 110–116. [Google Scholar] [CrossRef]
- Rubio-Moraga, A.; Ahrazem, O.; Pérez-Clemente, R.M.; Gómez-Cadenas, A.; Yoneyama, K.; López-Ráez, J.A.; Molina, R.V.; Gómez-Gómez., L. Apical dominance in saffron and the involvement of the branching enzymes CCD7 and CCD8 in the control of bud sprouting. BMC Plant Biol. 2014, 14, 171. [Google Scholar] [CrossRef] [PubMed]
- Mir, J.I.; Ahmed, N.; Singh, D.B.; Khan, M.H.; Zffer, S.; Shafi, W. Breeding and biotechnological opportunities in saffron crop improvement. Afri. J. Agri. Res. 2015, 10, 1970–1974. [Google Scholar]
- Mildažiene, V.; Paužaite, G.; Nauciene, Z.; Malakauskiene, A.; Žukiene, R.; Januškaitiene, I.; Jakštas, V.; Ivanauskas, L.; Filatova, I.; Lyushkevich, V. Pre-sowing seed treatment with cold plasma and electromagnetic field increases secondary metabolite content in purple coneflower (Echinacea purpurea) leaves. Plasma Process. Polym. 2018, 14, 1700059. [Google Scholar] [CrossRef]
- Mildaziene, V.; Pauzaite, G.; Nauciene, Z.; Zukiene, R.; Malakauskiene, A.; Norkeviciene, E.; Stukonis, V.; Slepetiene, A.; Olsauskaite, V.; Padarauskas, A.; et al. Effect of seed treatment with cold plasma and electromagnetic field on red clover germination, growth and content of major isoflavones. J. Phys. D Appl. Phys. 2020, 53, 26. [Google Scholar] [CrossRef]
- Mildaziene, V.; Ivankov, A.; Pauzaite, G.; Nauciene, Z.; Zukiene, R.; Degutyte-Fomins, L.; Pukalskas, A.; Venskutonis, P.R.; Filatova, I.; Lyuskevich, V. Seed treatment with cold plasma and electromagnetic field induces changes in red clover root growth dynamics, flavonoid exudation, and activates nodulation. Plasma Proc. Polym. 2020, 18, 2000160. [Google Scholar] [CrossRef]
- Ivankov, A.; Nauciene, Z.; Degutyte-Fomins, L.; Zukiene, R.; Januskaitiene, I.; Malakauskiene, A.; Jakstas, V.; Ivanauskas, L.; Romanovskaja, D.; Slepetiene, A.; et al. Changes in agricultural performance of common buckwheat induced by seed treatment with cold plasma and electromagnetic field. Appl. Sci. 2021, 11, 4391. [Google Scholar] [CrossRef]
- Holc, M.; Primc, G.; Iskra, J.; Titan, P.; Kovač, J.; Mozetič, M.; Junkar, I. Effect of Oxygen Plasma on Sprout and Root Growth, Surface Morphology and Yield of Garlic. Plants 2019, 8, 462. [Google Scholar] [CrossRef]
- Simpson, M.G. Plant Morphology. In Plant Systematics, 2nd ed.; Elsevier Inc.: Oxford, UK, 2010; pp. 451–513. [Google Scholar]
- Akbarian, M.; Shahidi, F.; Varidi, M.J.; Koocheki, A.; Roshanak, S. Effect of cold plasma on microbial and chemical properties of saffron. Saffron Agron. Technol. 2019, 7, 425–439. [Google Scholar]
- Luque de Castroa, M.D.; Quiles-Zafra, R. Appropriate use of analytical terminology – examples drawn from research on saffron. Talanta Open 2020, 2, 100005. [Google Scholar] [CrossRef]
- Richards, F.J.A. Flexible Growth Function for Empirical Use. J. Exp. Bot. 1959, 10, 290–300. [Google Scholar] [CrossRef]
- Muravnik, L.E. The structural peculiarities of the leaf glandular Trichomes: A review. In Plant Cell and Tissue Differentiation and Secondary Metabolites: Fundamentals and Applications; Ramawat, K.G., Ekiert, H.M., Goyal, S., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 63–97. [Google Scholar]
- Zarinkamar, F.; Tajik, S.; Soleimanpour, S. Effects of altitude on anatomy and concentration of crocin, picrocrocin and safranal in Crocus sativus L. Aust. J. Crop Sci. 2011, 5, 831–838. [Google Scholar]
- Mykhailenko, O.; Bezruk, I.; Ivanauskas, L.; Georgiyants, V. Comparative Analysis of the Major Metabolites of Ukrainian Saffron Samples by HPLC. Plant Foods Hum. Nutr. 2021, 76, 394–396. [Google Scholar] [CrossRef]
- Sánchez, A.M.; Carmona, M.; Ordoudi, S.A.; Tsimidou, M.Z.; Alonso, G.L. Kinetics of individual crocetin ester degradation in aqueous extracts of saffron (Crocus sativus L.) upon thermal treatment in the dark. J. Agric. Food Chem. 2008, 56, 1627–1637. [Google Scholar] [CrossRef] [PubMed]
- Ivankov, A.; Nauciene, Z.; Zukiene, R.; Degutyte-Fomins, L.; Malakauskiene, A.; Kraujalis, P.; Venskutonis, P.R.; Filatova, I.; Lyushkevich, V.; Mildaziene, V. Changes in growth and production of non-psychotropic cannabinoids induced by pre-sowing treatment of hemp seeds with cold plasma, vacuum and electromagnetic field. Appl. Sci. 2020, 10, 8519. [Google Scholar] [CrossRef]
- Sirgedaitė-Šėžienė, V.; Lučinskaitė, I.; Mildažienė, V.; Ivankov, A.; Koga, K.; Shiratani, M.; Laužikė, K.; Baliuckas, V. Changes in Content of Bioactive Compounds and Antioxidant Activity Induced in Needles of Different Half-Sib Families of Norway Spruce (Picea abies (L.) H. Karst) by Seed Treatment with Cold Plasma. Antioxidants 2022, 11, 1558. [Google Scholar] [CrossRef]
- Gasperini, D.; Howe, G.A. Phytohormones in a universe of regulatory metabolites: Lessons from jasmonate. Plant Physiol. 2024, 195, 135–154. [Google Scholar] [CrossRef]
- Lacchini, E.; Goossens, A. Combinatorial control of plant specialized metabolism: Mechanisms, functions, and consequences. Annu. Rev. Cell Dev. Biol. 2020, 36, 291–313. [Google Scholar] [CrossRef]
- Cazzonelli, C.I.; Pogson, B.J. Source to sink: Regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010, 15, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Rao, S.; Zhou, X.; Li, L. Plant carotenoids: Recent advances and future perspectives. Mol. Hortic. 2022, 2, 3. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pizá, M.C.; Prevosto, L.; Zilli, C.; Cejas, E.; Kelly, H.; Balestrasse, K. Effects of non–thermal plasmas on seed-borne Diaporthe/Phomopsis complex and germination parameters of soybean seeds. Innov. Food Sci. Emerg. 2018, 49, 82–91. [Google Scholar] [CrossRef]
- Mildažiene, V.; Aleknaviciute, V.; Žukiene, R.; Paužaite, G.; Nauciene, Z.; Filatova, I.; Lyushkevich, V.; Haimi, P.; Tamošiune, I.; Baniulis, D. Treatment of Common sunflower (Helianthus annus L.) seeds with radio-frequency electromagnetic field and cold plasma induces changes in seed phytohormone balance, seedling development and leaf protein expression. Sci. Rep. 2019, 9, 6437. [Google Scholar] [CrossRef]
- Degutyte-Fomins, L.; Paužaite, G.; Žukiene, R.; Mildažiene, V.; Koga, K.; Shiratani, M. Relationship between cold plasma treatment-induced changes in radish seed germination and phytohormone balance. Jpn. J. Appl. Phys. 2020, 59, SH1001. [Google Scholar] [CrossRef]
- Attri, P.; Ishikawa, K.; Okumura, T.; Koga, K.; Shiratani, M.; Mildaziene, M. Impact of seed color and storage time on the radish seed germination and sprout growth in plasma agriculture. Sci. Rep. 2021, 11, 2539. [Google Scholar] [CrossRef]
- Šerá, B.; Vanková, R.; Rohácek, K.; Šerý, M. Gliding arc plasma treatment of maize (Zea mays L.) grains promotes seed germination and early growth, affecting hormone pools, but not significantly photosynthetic parameters. Agronomy 2021, 11, 2066. [Google Scholar] [CrossRef]
- Iranbakhsh, A.; Ardebili, Z.O.; Molaei, H.; Ardebili, N.O.; Amini, M. Cold plasma up-regulated expressions of WRKY1 transcription factor and genes involved in biosynthesis of cannabinoids in hemp (Cannabis sativa L.). Plasma Chem. Plasma Process. 2020, 40, 527–537. [Google Scholar] [CrossRef]
- Ghasempour, M.; Iranbakhsh, A.; Ebadi, M.; Oraghi Ardebili, Z. Seed priming with cold plasma improved seedling performance, secondary metabolism, and expression of deacetylvindoline O-acetyltransferase gene in Catharanthus roseus. Contrib. Plasm. Phys. 2020, 60, e201900159. [Google Scholar] [CrossRef]
- Ghaemi, M.; Majd, A.; Iranbakhsh, A. Transcriptional responses following seed priming with cold plasma and electromagnetic field in Salvia nemorosa L. J. Theor. Appl. Phys. 2020, 14, 323–328. [Google Scholar] [CrossRef]
- Seddighinia, F.S.; Iranbakhsh, A.; Ardebili, Z.O.; Soleimanpour, S. Seed-priming with cold plasma and supplementation of nutrient solution with carbon nanotube enhanced carotenoid contents and the expression of psy and pds in Bitter melon (Momordica charantia). J. Appl. Bot. Food Qual. 2021, 94, 7–14. [Google Scholar]
- Leprince, O.; Pellizzaro, A.; Berriri, S.; Buitink, J. Late seed maturation: Drying without dying. J. Exp. Bot. 2016, 68, 827–841. [Google Scholar] [CrossRef]
- Sano, N.; Rajjou, L.; North, H.M.; Debeaujon, I.; Marion-Poll, A.; Seo, M. Staying alive: Molecular aspects of seed longevity. Plant Cell Physiol. 2016, 57, 660–674. [Google Scholar] [CrossRef] [PubMed]
- Renau-Morata, B.; Nebauer, S.G.; Sánchez, M.; Molina, R.V. Effect of corm size, water stress and cultivation conditions on photosynthesis and biomass partitioning during the vegetative growth of saffron (Crocus sativus L.). Ind. Crops Prod. 2012, 39, 40–46. [Google Scholar] [CrossRef]
- Barjasteh, A.; Lamichhane, P.; Dehghani, Z.; Kaushik, N.; Gupta, R.; Choi, E.H.; Kaushik, N.K. Recent Progress of Non-thermal Atmospheric Pressure Plasma for Seed Germination and Plant Development: Current Scenario and Future Landscape. J. Plant Growth Regul. 2023, 42, 5417–5432. [Google Scholar] [CrossRef]
- Takaku, Y.; Suzuki, H.; Ohta, I.; Tsutsui, T.; Matsumoto, H.; Shimomura, M.; Hariyama, T. A ‘NanoSuit’ surface shield successfully protects organisms in high vacuum: Observations on living organisms in an FE-SEM. Proc. R. Soc. B 2015, 282, 20142857. [Google Scholar] [CrossRef]
- Zhang, H.; Ma, X.; Li, W.; Niu, D.; Wang, Z.; Yan, X.; Yang, X.; Yang, Y.; Cui, H. Genome-wide characterization of NtHD-ZIP IV: Different roles in abiotic stress response and glandular Trichome induction. BMC Plant Biol. 2019, 19, 444. [Google Scholar] [CrossRef] [PubMed]
- Kjær, A.; Grevsen, K.; Jensen, M. Effect of external stress on density and size of glandular trichomes in full-grown Artemisia annua, the source of anti-malarial artemisinin. AoB Plants 2012, 2012, pls018. [Google Scholar] [CrossRef]
- Amini, M.; Ghoranneviss, M.; Abdijadid, S. Effect of cold plasma on crocin esters and volatile compounds of saffron. Food Chem. 2017, 235, 290–293. [Google Scholar] [CrossRef]
- Darvish, H.; Ramezan, Y.; Khani, M.R.; Kamkari, A. Effect of low-pressure cold plasma processing on decontamination and quality attributes of Saffron (Crocus sativus L.). Food Sci. Nutr. 2022, 10, 1082–2090. [Google Scholar] [CrossRef]
- Mykhailenko, O.; Petrikaitė, V.; Korinek, M.; El-Shazly, M.; Chen, B.-H.; Yen, C.-H.; Hsieh, C.-F.; Bezruk, I.; Dabrišiūtė, A.; Ivanauskas, L.; et al. Bio-guided bioactive profiling and HPLC-DAD fingerprinting of Ukrainian saffron (Crocus sativus stigma): Moving from Correlation toward Causation. BMC Complement. Med. Ther. 2021, 21, 203–218. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indice/Treatment Group | Control | Vacuum3 | CP3 | CP5 | EMF5 |
|---|---|---|---|---|---|
| Vi (%) | 100.0 ± 0.0 | 100.0 ± 0.0 | 100.0 ± 0.0 | 100.0 ± 0.0 | 100.0 ± 0.0 |
| Me (days) | 25.6 ± 0.5 | 25.1 ± 0.4 | 24.5 ± 1.1 | 29.5 ± 0.5 * | 25.3 ± 1.1 |
| Qu (days) | 3.6 ± 0.3 | 2.6 ± 0.1 * | 3.3 ± 0.2 | 3.0 ± 0.1 | 3.2 ± 0.4 |
| Bud Number per Corm | Control | Vacuum3 | CP3 | CP5 | EMF5 |
|---|---|---|---|---|---|
| 1 | 16.0 | 9.5 | 14.3 | 33.3 | 9.5 |
| 2 | 12.0 | 23.8 | 38.1 | 33.3 | 28.6 |
| 3 | 36.0 | 19.0 | 23.8 | 28.6 | 38.1 |
| 4 | 28.0 | 28.6 | 14.3 | 4.8 | 19.0 |
| 5 | 4.0 | 9.5 | 9.5 | 0.0 | 4.8 |
| 6 | 4.0 | 4.8 | 0.0 | 0.0 | 0.0 |
| 7 | 0.0 | 4.8 | 0.0 | 0.0 | 0.0 |
| Morphometric Parameter | Control (6) | Vacuum3 (6) | CP3 (4) | EMF5 (7) |
|---|---|---|---|---|
| Flower length, cm | 7.90 ± 0.00 | 8.00 ± 0.50 | 4.50 ± 0.50 * | 10.00 ± 0.00 * |
| Flower fresh weight, g | 0.43 ± 0.05 | 0.46 ± 0.05 | 0.22 ± 0.03 | 0.52 ± 0.07 |
| Flower dry weight, g | 0.05 ± 0.002 | 0.05 ± 0.003 | 0.02 ± 0.001 * | 0.07 ± 0.01 |
| Pistil length, cm | 4.10 ± 0.20 | 4.07 ± 0.23 | 3.80 ± 0.18 | 3.87 ± 0.19 |
| Pistil fresh weight, g | 0.03 ± 0.003 | 0.03 ± 0.003 | 0.02 ± 0.002 | 0.03 ± 0.009 |
| Pistil dry weight, g | 0.004 ± 0.0005 | 0.007 ± 0.002 | 0.004 ± 0.0007 | 0.004 ± 0.0003 |
| Compound | No Glucose Moieties | Abbreviation ** | tR (min) | UV/Vis, λmax, nm | MW, g·mol−1 | MolFor |
|---|---|---|---|---|---|---|
| trans-crocetin (tri-β-D-glucosyl)(β-D-gentiobiosyl) ester | Crocin 5 | t-5tG | 33.190 | 262, 332sh, 443, 468 | 1138 | C50H74O29 |
| trans-crocetin (β-D-neapolitanosyl)(β-D-gentiobiosyl) ester | - | t-5nG | 34.928 | 234, 300sh, 418, 439 | 1138 | C50H74O29 |
| trans-crocetin (β-D-neapolitanosyl)(β-D-glucosyl) ester | - | t-4ng | 35.630 | 263, 441, 466 | 976 | C44H64O24 |
| trans-crocetin di(β-D-gentiobiosyl)ester | Crocin 4 | t-4GG | 37.825 | 261, 331sh, 440, 466 | 976 | C44H64O24 |
| trans-crocetin (β-D-gentiobiosyl)(β-D-glucosyl) ester | Crocin 3 | t-3Gg | 39.608 | 261, 327sh, 440, 466 | 814 | C38H54O19 |
| trans-crocetin mono(β-D-gentiobiosyl) ester | Crocin 2 | t-2G | 40.424 | 261, 331sh, 440, 466 | 652 | C33H44O14 |
| cis-crocetin (β-D-neapolitanosyl)(β-D-gentiobiosyl) ester | - | c-5nG | 42.119 | 242, 260, 414, 436 | 1139 | C50H74O29 |
| trans-crocetin di(β-D-glucosyl) ester | Crocin 2′ | t-2gg | 43.147 | 261, 322, 439, 465 | 652 | C33H44O14 |
| cis-crocetin di(β-D-gentiobiosyl)ester | cis-Crocin 4 | c-4GG | 47.084 | 262, 326, 435, 457 | 976 | C44H64O24 |
| cis-crocetin (β-D-gentiobiosyl)(β-D-glucosyl) ester | cis-Crocin 3 | c-3Gg | 48.252 | 258, 325, 435, 460 | 814 | C38H54O19 |
| cis-crocetin mono(β-D-gentiobiosyl) ester | cis-Crocin 2 | c-2G | 49.511 | 262, 327, 433, 456 | 652 | C33H44O14 |
| cis-crocetin di(β-D-glucosyl) ester | cis-Crocin 2′ | c-2gg | 49.711 | 262, 326, 434, 456sh | 652 | C33H44O14 |
| trans-crocetin mono(β-D-glucosyl) ester | Crocin 1 | t-1g | 50.592 | 258, 434, 459 | 490 | C26H34O9 |
| Crocin derivative | - | - | 52.256 | 258, 434, 459 | 479~ | C44H64O24 |
| Crocin derivative | - | - | 52.685 | 258, 327, 434 | 479~ | C44H64O24 |
| cis-crocetin (tri-β-D-glucosyl)(β-D-gentiobiosyl) ester | cis-Crocin 5 | c-5tG | 53.893 | 263, 332sh, 445, 469 | 1138 | C50H74O29 |
| Crocin derivative | - | - | 54.780 | 259, 323, 428, 452 | 479~ | C44H64O24 |
| Crocin derivative | - | - | 55.140 | 245sh, 332, 431, 454 | 479~ | C44H64O24 |
| cis-crocetin mono(β-D-glucosyl) ester | cis-Crocin 1 | c-1g | 55.614 | 259, 323, 428, 452 | 490 | C26H34O9 |
| Crocin derivative | - | - | 57.965 | 258, 324sh, 434, 460 | 479~ | C44H64O24 |
| Crocin derivative | - | - | 61.007 | 249, 326, 430, 453 | 479~ | C44H64O24 |
| trans-crocetin | - | t-crocetin | 64.716 | 258, 434, 459 | 328 | C20H24O4 |
| cis-crocetin | - | c-crocetin | 67.827 | 255, 318sh, 427, 453 | 328 | C20H24O4 |
| Code 1 | Compound | Rt. min | Control | Vacuum3 | CP3 | EMF5 |
|---|---|---|---|---|---|---|
| A | Picrocrocin | 16.49 | 720.6 ± 88.4 | 965.6 ± 9.2 * | 395.4 ± 2.4 * | 847.3 ± 6.2 * |
| B | Rutin | 20.81 | 889.0 ± 147.2 | 762.6 ± 21.0 | 171.0 ± 2.0 * | 750.0 ± 33.1 |
| C | trans-crocetin (tri-β-D-glucosyl)(β-D-gentiobiosyl) ester | 33.19 | 2180.6 ± 19.4 | 2085.0 ± 130.1 | 591.2 ± 3.2 * | 18,898.4 ± 17,021.5 ** |
| D | trans-crocetin (β-D-neapolitanosyl)(β-D-gentiobiosyl) ester | 34.93 | 867.2 ± 90.4 | 786.6 ± 6.5 | 76.4 ± 1.6 * | 6356.2 ± 251.4 ** |
| E | trans-crocetin (β-D-neapolitanosyl)(β-D-glucosyl) ester | 35.63 | 357.0 ± 83.0 | - | - | 2034.7 ± 692.2 ** |
| F | trans-crocetin di(β-D-gentiobiosyl)ester * | 37.83 | 114,319.0 ± 513.6 | 124,350.4 ± 996.2 | 8949.1 ± 38.2 * | 431,173.0 ± 965.0 * |
| G | trans-crocetin (β-D-gentiobiosyl)(β-D-glucosyl) ester | 39.61 | 1714.2 ± 90.6 | 4225.0 ± 71.9 * | 886.8 ± 9.2 * | 37,187.7 ± 144.9 * |
| H | trans-crocetin mono(β-D-gentiobiosyl) ester | 40.42 | 49,042.2 ± 1983.0 | 42,026.4 ± 251.0 * | 6399.4 ± 101.8 * | 188,232.6 ± 1415.9 ** |
| I | cis-crocetin (β-D-neapolitanosyl)(β-D-gentiobiosyl) ester | 42.12 | 1200.8 ± 24.6 | 575.0 ± 49.6 * | 131.6 ± 14.8 * | 781.9 ± 13.9 * |
| J | trans-crocetin di(β-D-glucosyl) ester | 43.15 | 1735.6 ± 127.6 | 1363.8 ± 90.5 * | 207.0 ± 1.4 * | 1197.8 ± 34.7 * |
| K | cis-crocetin di(β-D-gentiobiosyl)ester | 47.08 | 25,398.0 ± 195.0 | 12,675.6 ± 87.6 | 1899.8 ± 30.6 * | 20,536.6 ± 74.9 * |
| L | cis-crocetin (β-D-gentiobiosyl)(β-D-glucosyl) ester | 48.25 | 23,894.2 ± 137.8 | 21,464.2 ± 285.2 * | 6270.6 ± 43.0 * | 164,859.6 ± 14,739.5 ** |
| M | cis-crocetin mono(β-D-gentiobiosyl) ester | 49.51 | 3594.4 ± 162.4 | 1878.0 ± 19.0 * | 192.2 ± 30.4 * | 2530.7 ± 3.5 * |
| O | cis-crocetin di(β-D-glucosyl) ester | 49.71 | 5529.8 ± 296.0 | 2812.1 ± 39.8 * | 309.4 ± 19.2 * | 17,151.2 ± 123.7 ** |
| P | trans-crocetin mono(β-D-glucosyl) ester | 50.59 | - | 257.9 ± 14.9 * | 78.6 ± 5.8 * | 1446.8 ± 124.9 * |
| Q | Crocin derivative | 52.26 | 792.6 ± 56.6 | 400.5 ± 7.1 * | 69.0 ± 0.18 * | 4345.0 ± 401.2 ** |
| R | Crocin derivative | 52.69 | - | - | 61.6 ± 1.8 | 511.5 ± 33.4 |
| S | cis-crocetin (tri-β-D-glucosyl)(β-D-gentiobiosyl) ester | 53.89 | 251.4 ± 12.2 | 458.2 ± 11.8 * | 95.6 ± 2.8 * | 4876.2 ± 404.6 * |
| T | Crocin derivative | 54.78 | 1289.6 ± 36.6 | 734.4 ± 3.0 * | 171.8 ± 7.6 * | 3238.4 ± 328.5 * |
| U | Crocin derivative | 55.14 | - | - | - | 1531.6 ± 84.3 |
| V | cis-crocetin mono(β-D-glucosyl) ester | 55.61 | 1297.2 ± 16.0 | 737.6 ± 6.4 * | 167.4 ± 9.2 * | 3810.5 ± 157.4 ** |
| W | Crocin derivative | 57.97 | 1528.4 ± 85.6 | 1115.2 ± 55.7 * | 144.6 ± 33.0 * | 1374.1 ± 18.1 * |
| X | Crocin derivative | 61.01 | - | - | - | 200.4 ± 23.6 |
| Y | Safranal | 62.28 | 8.4 ± 1.8 | 10.3 ± 0.5 | 1.60 ± 0.04 * | 16.3 ± 0.2 ** |
| Z | trans-crocetin | 64.72 | - | - | 197.7 ± 1.2 * | 225.4 ± 9.1 * |
| AA | cis-crocetin | 67.83 | - | - | - | 204.6 ± 8.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mildažienė, V.; Žūkienė, R.; Fomins, L.D.; Naučienė, Z.; Minkutė, R.; Jarukas, L.; Drapak, I.; Georgiyants, V.; Novickij, V.; Koga, K.; et al. Effects of Corm Treatment with Cold Plasma and Electromagnetic Field on Growth and Production of Saffron Metabolites in Crocus sativus. Int. J. Mol. Sci. 2024, 25, 10412. https://doi.org/10.3390/ijms251910412
Mildažienė V, Žūkienė R, Fomins LD, Naučienė Z, Minkutė R, Jarukas L, Drapak I, Georgiyants V, Novickij V, Koga K, et al. Effects of Corm Treatment with Cold Plasma and Electromagnetic Field on Growth and Production of Saffron Metabolites in Crocus sativus. International Journal of Molecular Sciences. 2024; 25(19):10412. https://doi.org/10.3390/ijms251910412
Chicago/Turabian StyleMildažienė, Vida, Rasa Žūkienė, Laima Degutytė Fomins, Zita Naučienė, Rima Minkutė, Laurynas Jarukas, Iryna Drapak, Victoriya Georgiyants, Vitalij Novickij, Kazunori Koga, and et al. 2024. "Effects of Corm Treatment with Cold Plasma and Electromagnetic Field on Growth and Production of Saffron Metabolites in Crocus sativus" International Journal of Molecular Sciences 25, no. 19: 10412. https://doi.org/10.3390/ijms251910412
APA StyleMildažienė, V., Žūkienė, R., Fomins, L. D., Naučienė, Z., Minkutė, R., Jarukas, L., Drapak, I., Georgiyants, V., Novickij, V., Koga, K., Shiratani, M., & Mykhailenko, O. (2024). Effects of Corm Treatment with Cold Plasma and Electromagnetic Field on Growth and Production of Saffron Metabolites in Crocus sativus. International Journal of Molecular Sciences, 25(19), 10412. https://doi.org/10.3390/ijms251910412