
Impact of Epigenetics, Diet, and Nutrition-Related Pathologies on Wound Healing


Abstract
:1. Introduction
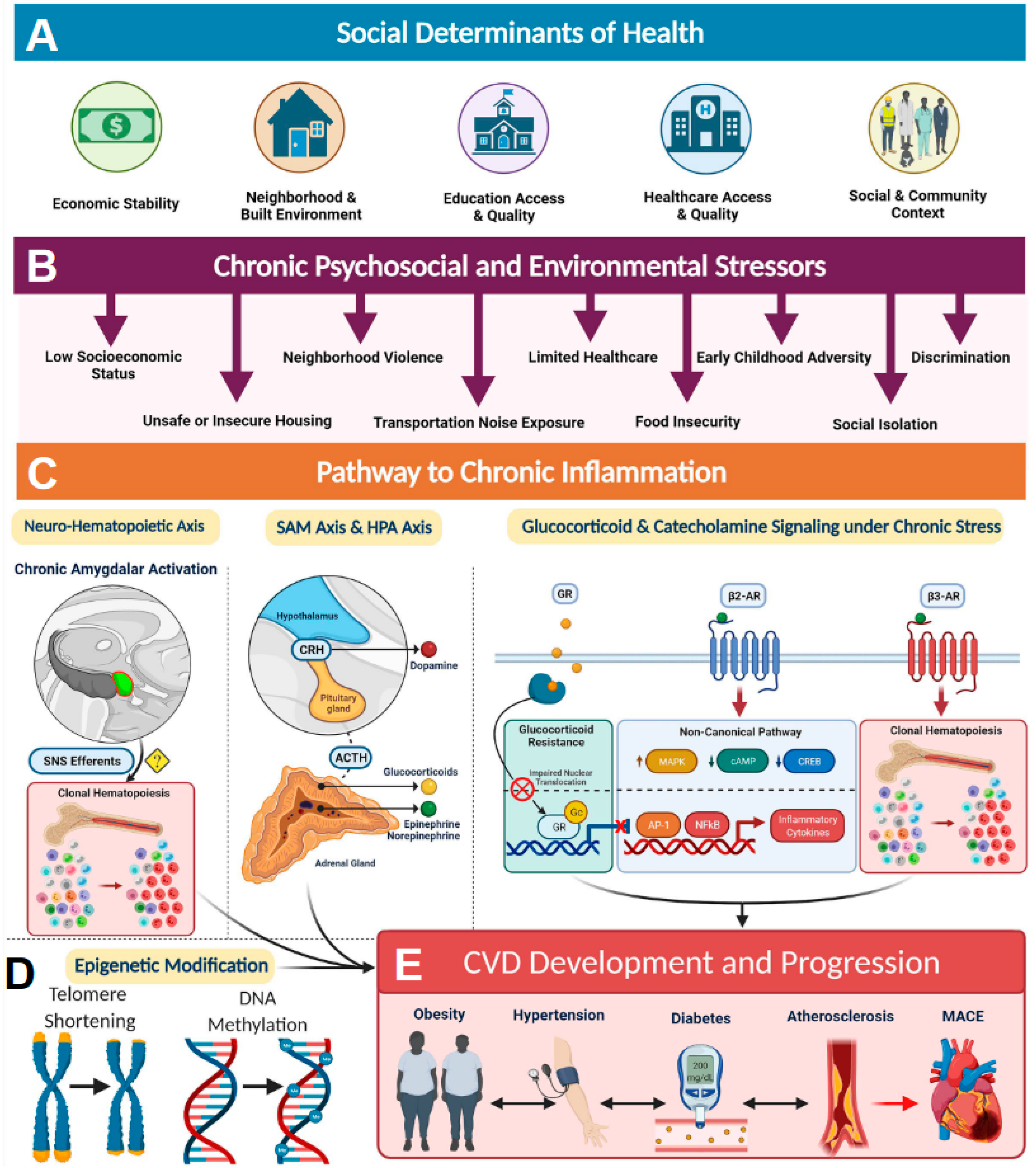
2. Socioeconomics and Health
3. Genomics and Health
3.1. Sociogenomics
3.2. Genomic Wide Association Studies (GWAS) in Diabetes and Wound Healing
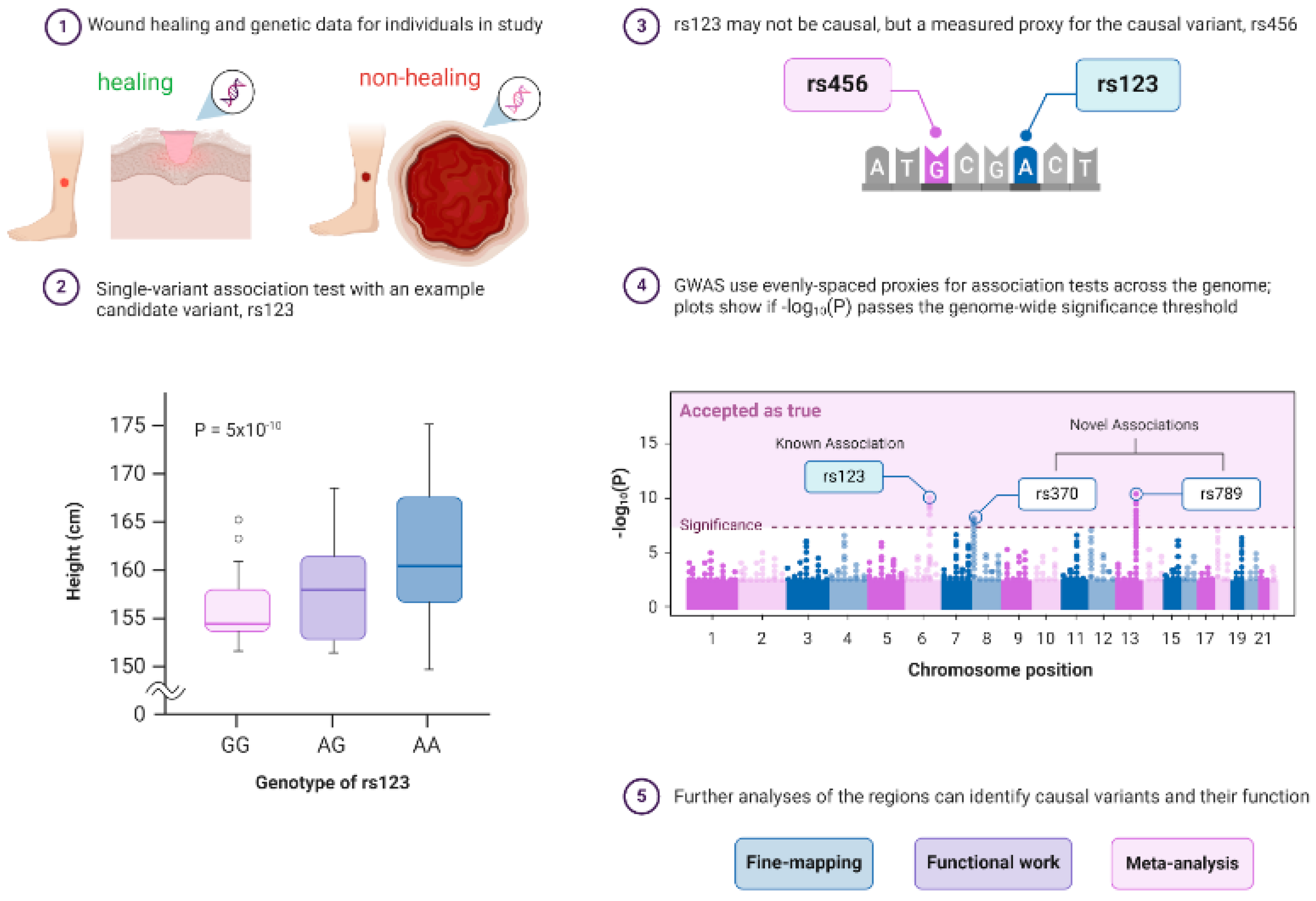
3.3. Epigenetics
4. The Process of Physiological Wound Healing
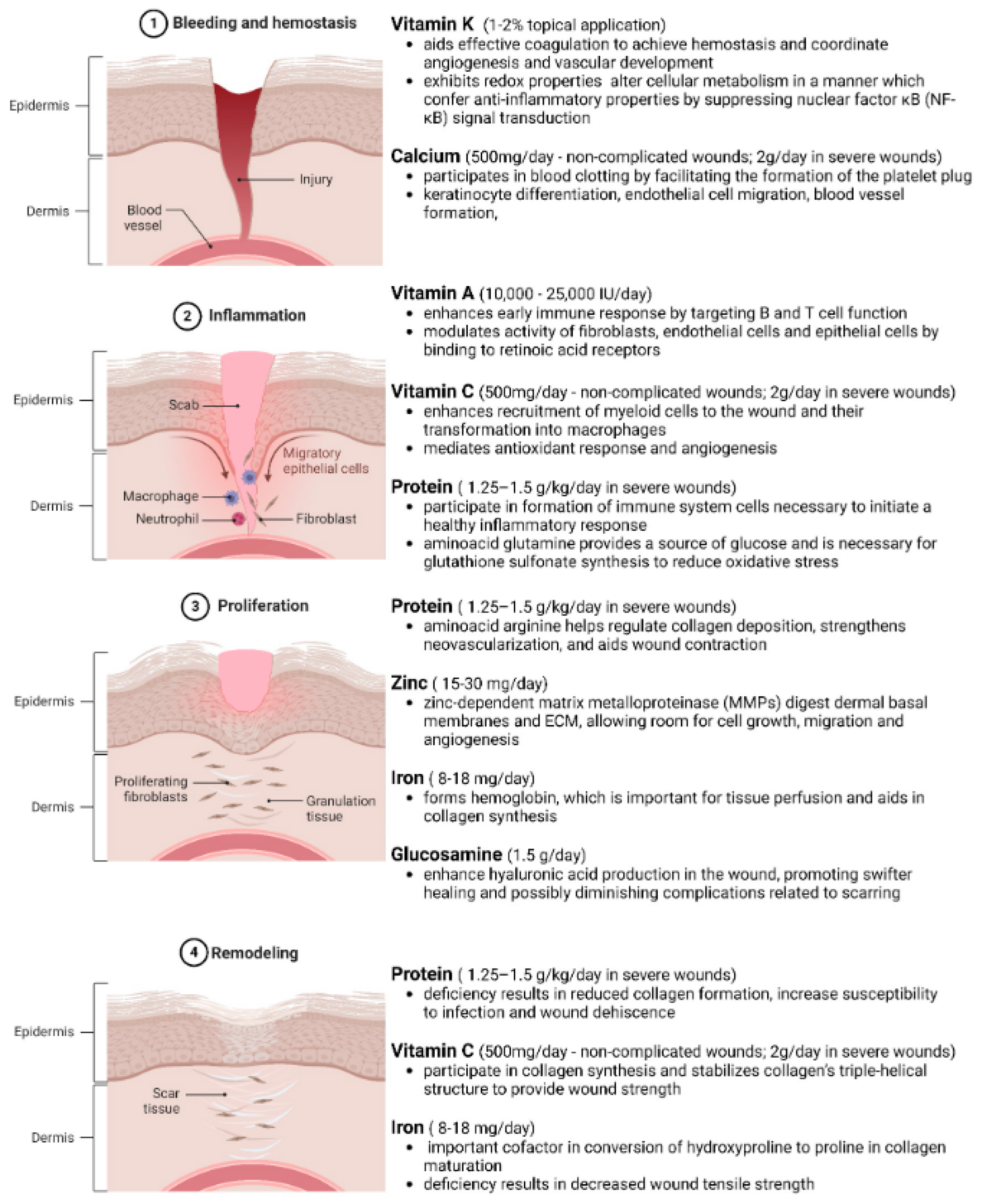
4.1. Genetics and Molecular Mechanisms of the Wound Healing Process
4.1.1. Genetics and Molecular Mechanisms in the Hemostasis Phase of Wound Repair
4.1.2. Genetics and Molecular Mechanisms in the Inflammation Phase of Wound Repair
4.1.3. Genetics and Molecular Mechanisms in the Proliferation Phase of Wound Repair
4.1.4. Genetics and Molecular Mechanisms in the Remodeling Phase of Wound Repair
5. Impact of Socioeconomic Status on the Healing Process
5.1. Pathophysiological Mechanisms of Altered Gene Expression in Chronic Wounds
5.1.1. The Conserved Transcriptional Response to Adversity (CRTA)
5.1.2. Diabetes and Impairment of Wound Healing
6. Impact of Nutrition on the Healing Process
6.1. Specific Genes and SNPs Associated with Nutritional Pathology and Poor Wound Healing
6.1.1. MTHFR
6.1.2. VDR
6.1.3. MMP-9
7. Macronutrients and Micronutrients in Wound Healing
7.1. Protein
7.2. Fatty Acids
7.3. Carbohydrates
7.4. Vitamins
7.4.1. Vitamin A
7.4.2. Vitamin B
7.4.3. Vitamin C and K
7.4.4. Vitamin D
7.4.5. Vitamin E
7.5. Mineral Nutrients
7.6. Clinical Evidence of Nutritional Supplementation on Wound Healing
8. Obesity, Oxygen, and Hypoxia
Exercise, Obesity, and Wound Healing
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sen, C.K. Human Wound and Its Burden: Updated 2022 Compendium of Estimates. Adv. Wound Care 2023, 12, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Sen, C.K. Human Wounds and Its Burden: An Updated Compendium of Estimates. Adv. Wound Care 2019, 8, 39–48. [Google Scholar] [CrossRef]
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of Obesity and Severe Obesity Among Adults: United States, 2017–2018. NCHS Data Brief. 2020, 360, 1–8. [Google Scholar]
- Sen, C.K.; Gordillo, G.M.; Roy, S.; Kirsner, R.; Lambert, L.; Hunt, T.K.; Gottrup, F.; Gurtner, G.C.; Longaker, M.T. Human skin wounds: A major and snowballing threat to public health and the economy. Wound Repair Regen. 2009, 17, 763–771. [Google Scholar] [CrossRef]
- Morton, L.M.; Phillips, T.J. Wound healing and treating wounds: Differential diagnosis and evaluation of chronic wounds. J. Am. Acad. Dermatol. 2016, 74, 589–605; quiz 605–606. [Google Scholar] [CrossRef]
- Serra, R.; Grande, R.; Butrico, L.; Rossi, A.; Settimio, U.F.; Caroleo, B.; Amato, B.; Gallelli, L.; de Franciscis, S. Chronic wound infections: The role of Pseudomonas aeruginosa and Staphylococcus aureus. Expert Rev. Anti Infect. Ther. 2015, 13, 605–613. [Google Scholar] [CrossRef]
- Walicka, M.; Raczynska, M.; Marcinkowska, K.; Lisicka, I.; Czaicki, A.; Wierzba, W.; Franek, E. Amputations of Lower Limb in Subjects with Diabetes Mellitus: Reasons and 30-Day Mortality. J. Diabetes Res. 2021, 2021, 8866126. [Google Scholar] [CrossRef]
- Gordois, A.; Scuffham, P.; Shearer, A.; Oglesby, A.; Tobian, J.A. The health care costs of diabetic peripheral neuropathy in the US. Diabetes Care 2003, 26, 1790–1795. [Google Scholar] [CrossRef] [PubMed]
- Girijala, R.L.; Bush, R.L. Review of Socioeconomic Disparities in Lower Extremity Amputations: A Continuing Healthcare Problem in the United States. Cureus 2018, 10, e3418. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Lin, C.W.; Yang, H.M.; Hung, S.Y.; Chen, I.W. Survival and associated risk factors in patients with diabetes and amputations caused by infectious foot gangrene. J. Foot Ankle Res. 2018, 11, 1. [Google Scholar] [CrossRef]
- Conger, R.D.; Conger, K.J.; Martin, M.J. Socioeconomic Status, Family Processes, and Individual Development. J. Marriage Fam. 2010, 72, 685–704. [Google Scholar] [CrossRef] [PubMed]
- Fiscella, K.; Williams, D.R. Health disparities based on socioeconomic inequities: Implications for urban health care. Acad. Med. 2004, 79, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Becker, G.; Newsom, E. Socioeconomic status and dissatisfaction with health care among chronically ill African Americans. Am. J. Public Health 2003, 93, 742–748. [Google Scholar] [CrossRef]
- Powell-Wiley, T.M.; Baumer, Y.; Baah, F.O.; Baez, A.S.; Farmer, N.; Mahlobo, C.T.; Pita, M.A.; Potharaju, K.A.; Tamura, K.; Wallen, G.R. Social Determinants of Cardiovascular Disease. Circ. Res. 2022, 130, 782–799. [Google Scholar] [CrossRef]
- Steptoe, A.; Zaninotto, P. Lower socioeconomic status and the acceleration of aging: An outcome-wide analysis. Proc. Natl. Acad. Sci. USA 2020, 117, 14911–14917. [Google Scholar] [CrossRef]
- Cohen, S.; Doyle, W.J.; Baum, A. Socioeconomic status is associated with stress hormones. Psychosom. Med. 2006, 68, 414–420. [Google Scholar] [CrossRef]
- Jones, C.; Gwenin, C. Cortisol level dysregulation and its prevalence-Is it nature’s alarm clock? Physiol. Rep. 2021, 8, e14644. [Google Scholar] [CrossRef] [PubMed]
- Sen, C.K.; Roy, S. Sociogenomic Approach to Wound Care: A New Patient-Centered Paradigm. Adv. Wound Care 2019, 8, 523–526. [Google Scholar] [CrossRef]
- French, S.A.; Tangney, C.C.; Crane, M.M.; Wang, Y.; Appelhans, B.M. Nutrition quality of food purchases varies by household income: The SHoPPER study. BMC Public Health 2019, 19, 231. [Google Scholar] [CrossRef]
- Viinikainen, J.; Bryson, A.; Bockerman, P.; Kari, J.T.; Lehtimaki, T.; Raitakari, O.; Viikari, J.; Pehkonen, J. Does better education mitigate risky health behavior? A mendelian randomization study. Econ. Hum. Biol. 2022, 46, 101134. [Google Scholar] [CrossRef]
- Garrett, B.E.; Martell, B.N.; Caraballo, R.S.; King, B.A. Socioeconomic Differences in Cigarette Smoking Among Sociodemographic Groups. Prev. Chronic Dis. 2019, 16, E74. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, J.C.; Browning, K.K. Smoking, chronic wound healing, and implications for evidence-based practice. J. Wound Ostomy Cont. Nurs. 2014, 41, 415–423; quiz E1-2. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.P.; Bovbjerg, D.H.; Marsland, A.L. Glucocorticoid resistance and beta2-adrenergic receptor signaling pathways promote peripheral pro-inflammatory conditions associated with chronic psychological stress: A systematic review across species. Neurosci. Biobehav. Rev. 2021, 128, 117–135. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Janicki-Deverts, D.; Doyle, W.J.; Miller, G.E.; Frank, E.; Rabin, B.S.; Turner, R.B. Chronic stress, glucocorticoid receptor resistance, inflammation, and disease risk. Proc. Natl. Acad. Sci. USA 2012, 109, 5995–5999. [Google Scholar] [CrossRef] [PubMed]
- Del Giacco, L.; Cattaneo, C. Introduction to genomics. Methods Mol. Biol. 2012, 823, 79–88. [Google Scholar] [PubMed]
- Goodarzi, M.O. Genetics of obesity: What genetic association studies have taught us about the biology of obesity and its complications. Lancet Diabetes Endocrinol. 2018, 6, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.F.; Mardis, E.R. The emerging clinical relevance of genomics in cancer medicine. Nat. Rev. Clin. Oncol. 2018, 15, 353–365. [Google Scholar] [CrossRef]
- Robinson, G.E.; Grozinger, C.M.; Whitfield, C.W. Sociogenomics: Social life in molecular terms. Nat. Rev. Genet. 2005, 6, 257–270. [Google Scholar] [CrossRef]
- Singh, K.; Rustagi, Y.; Abouhashem, A.S.; Tabasum, S.; Verma, P.; Hernandez, E.; Pal, D.; Khona, D.K.; Mohanty, S.K.; Kumar, M.; et al. Genome-wide DNA hypermethylation opposes healing in patients with chronic wounds by impairing epithelial-mesenchymal transition. J. Clin. Investig. 2022, 132, e157279. [Google Scholar] [CrossRef]
- Singh, K.; Pal, D.; Sinha, M.; Ghatak, S.; Gnyawali, S.C.; Khanna, S.; Roy, S.; Sen, C.K. Epigenetic Modification of MicroRNA-200b Contributes to Diabetic Vasculopathy. Mol. Ther. 2017, 25, 2689–2704. [Google Scholar] [CrossRef]
- Idaghdour, Y.; Czika, W.; Shianna, K.V.; Lee, S.H.; Visscher, P.M.; Martin, H.C.; Miclaus, K.; Jadallah, S.J.; Goldstein, D.B.; Wolfinger, R.D.; et al. Geographical genomics of human leukocyte gene expression variation in southern Morocco. Nat. Genet. 2010, 42, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Archie, E.A.; Altmann, J.; Alberts, S.C. Social status predicts wound healing in wild baboons. Proc. Natl. Acad. Sci. USA 2012, 109, 9017–9022. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, J.R.; Mutesa, L.; Uddin, M. Traumatic Stress Epigenetics. Curr. Behav. Neurosci. Rep. 2018, 5, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Juruena, M.F.; Eror, F.; Cleare, A.J.; Young, A.H. The Role of Early Life Stress in HPA Axis and Anxiety. Adv. Exp. Med. Biol. 2020, 1191, 141–153. [Google Scholar]
- Schroeder, K.; Schuler, B.R.; Kobulsky, J.M.; Sarwer, D.B. The association between adverse childhood experiences and childhood obesity: A systematic review. Obes. Rev. 2021, 22, e13204. [Google Scholar] [CrossRef] [PubMed]
- Nowacka-Chmielewska, M.; Liskiewicz, D.; Liskiewicz, A.; Marczak, L.; Wojakowska, A.; Jerzy Barski, J.; Malecki, A. Cerebrocortical proteome profile of female rats subjected to the western diet and chronic social stress. Nutr. Neurosci. 2022, 25, 567–580. [Google Scholar] [CrossRef]
- Witte, J.S. Genome-wide association studies and beyond. Annu. Rev. Public Health 2010, 31, 9–20. [Google Scholar] [CrossRef]
- Hunter-Zinck, H.; Shi, Y.; Li, M.; Gorman, B.R.; Ji, S.G.; Sun, N.; Webster, T.; Liem, A.; Hsieh, P.; Devineni, P.; et al. Genotyping Array Design and Data Quality Control in the Million Veteran Program. Am. J. Hum. Genet. 2020, 106, 535–548. [Google Scholar] [CrossRef]
- DeForest, N.; Majithia, A.R. Genetics of Type 2 Diabetes: Implications from Large-Scale Studies. Curr. Diab. Rep. 2022, 22, 227–235. [Google Scholar] [CrossRef]
- Diegelmann, R.F.; Evans, M.C. Wound healing: An overview of acute, fibrotic and delayed healing. Front. Biosci. 2004, 9, 283–289. [Google Scholar] [CrossRef]
- Stone, R.C.; Chen, V.; Burgess, J.; Pannu, S.; Tomic-Canic, M. Genomics of Human Fibrotic Diseases: Disordered Wound Healing Response. Int. J. Mol. Sci. 2020, 21, 8590. [Google Scholar] [CrossRef] [PubMed]
- Visscher, P.M.; Wray, N.R.; Zhang, Q.; Sklar, P.; McCarthy, M.I.; Brown, M.A.; Yang, J. 10 Years of GWAS Discovery: Biology, Function, and Translation. Am. J. Hum. Genet. 2017, 101, 5–22. [Google Scholar] [CrossRef]
- McCarthy, M.I.; Abecasis, G.R.; Cardon, L.R.; Goldstein, D.B.; Little, J.; Ioannidis, J.P.A.; Hirschhorn, J.N. Genome-wide association studies for complex traits: Consensus, uncertainty and challenges. Nat. Rev. Genet. 2008, 9, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Tammen, S.A.; Friso, S.; Choi, S.W. Epigenetics: The link between nature and nurture. Mol. Aspects Med. 2013, 34, 753–764. [Google Scholar] [CrossRef]
- Bhamidipati, T.; Kumar, M.; Verma, S.S.; Mohanty, S.K.; Kacar, S.; Reese, D.; Martinez, M.M.; Kamocka, M.M.; Dunn, K.W.; Sen, C.K.; et al. Epigenetic basis of diabetic vasculopathy. Front. Endocrinol. 2022, 13, 989844. [Google Scholar] [CrossRef] [PubMed]
- Bhamidipati, T.; Sinha, M.; Sen, C.K.; Singh, K. Laser Capture Microdissection in the Spatial Analysis of Epigenetic Modifications in Skin: A Comprehensive Review. Oxid. Med. Cell. Longev. 2022, 2022, 4127238. [Google Scholar] [CrossRef]
- Harkless, R.; Singh, K.; Christman, J.; McCarty, A.; Sen, C.; Jalilvand, A.; Wisler, J. Microvesicle-Mediated Transfer of DNA Methyltransferase Proteins Results in Recipient Cell Immunosuppression. J. Surg. Res. 2023, 283, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Wisler, J.R.; Singh, K.; McCarty, A.; Harkless, R.; Karpurapu, M.; Hernandez, E.; Mukherjee, D.; Abouhashem, A.S.; Christman, J.W.; Sen, C.K. Exosomal Transfer of DNA Methyl-Transferase mRNA Induces an Immunosuppressive Phenotype in Human Monocytes. Shock 2022, 57, 218–227. [Google Scholar] [CrossRef]
- Dakhlallah, D.A.; Wisler, J.; Gencheva, M.; Brown, C.M.; Leatherman, E.R.; Singh, K.; Brundage, K.; Karsies, T.; Dakhlallah, A.; Witwer, K.W.; et al. Circulating extracellular vesicle content reveals de novo DNA methyltransferase expression as a molecular method to predict septic shock. J. Extracell Vesicles 2019, 8, 1669881. [Google Scholar] [CrossRef]
- Wang, Y.; Hou, C.; Wisler, J.; Singh, K.; Wu, C.; Xie, Z.; Lu, Q.; Zhou, Z. Elevated histone H3 acetylation is associated with genes involved in T lymphocyte activation and glutamate decarboxylase antibody production in patients with type 1 diabetes. J. Diabetes Investig. 2019, 10, 51–61. [Google Scholar] [CrossRef]
- Ling, C.; Ronn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 29, 1028–1044. [Google Scholar] [CrossRef] [PubMed]
- Margo, G.M.; Manring, J.M. The current literature on inpatient psychotherapy. Hosp. Community Psychiatry 1989, 40, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.H.; Huang, B.S.; Horng, H.C.; Yeh, C.C.; Chen, Y.J. Wound healing. J. Chin. Med. Assoc. 2018, 81, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Westman, J.; Grinstein, S.; Marques, P.E. Phagocytosis of Necrotic Debris at Sites of Injury and Inflammation. Front. Immunol. 2019, 10, 3030. [Google Scholar] [CrossRef] [PubMed]
- Gharbia, F.Z.; Abouhashem, A.S.; Moqidem, Y.A.; Elbaz, A.A.; Abdellatif, A.; Singh, K.; Sen, C.K.; Azzazy, H.M.E. Adult skin fibroblast state change in murine wound healing. Sci. Rep. 2023, 13, 886. [Google Scholar] [CrossRef]
- Zhou, X.; Brown, B.A.; Siegel, A.P.; El Masry, M.S.; Zeng, X.; Song, W.; Das, A.; Khandelwal, P.; Clark, A.; Singh, K.; et al. Exosome-Mediated Crosstalk between Keratinocytes and Macrophages in Cutaneous Wound Healing. ACS Nano. 2020, 14, 12732–12748. [Google Scholar] [CrossRef]
- Nissinen, L.M.; Kahari, V.M. Collagen Turnover in Wound Repair—A Macrophage Connection. J. Invest. Dermatol. 2015, 135, 2350–2352. [Google Scholar] [CrossRef]
- Nagle, S.M.; Stevens, K.A.; Wilbraham, S.C. Wound Assessment; StatPearls: Treasure Island, FL, USA, 2024. [Google Scholar]
- Holinstat, M. Normal platelet function. Cancer Metastasis Rev. 2017, 36, 195–198. [Google Scholar] [CrossRef]
- Smith, S.A.; Travers, R.J.; Morrissey, J.H. How it all starts: Initiation of the clotting cascade. Crit. Rev. Biochem. Mol. Biol. 2015, 50, 326–336. [Google Scholar] [CrossRef]
- Kaneko, N.; Kurata, M.; Yamamoto, T.; Morikawa, S.; Masumoto, J. The role of interleukin-1 in general pathology. Inflamm. Regen. 2019, 39, 12. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Zelova, H.; Hosek, J. TNF-alpha signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. 2013, 62, 641–651. [Google Scholar] [CrossRef]
- Zhao, R.; Liang, H.; Clarke, E.; Jackson, C.; Xue, M. Inflammation in Chronic Wounds. Int. J. Mol. Sci. 2016, 17, 2085. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed]
- King, A.; Balaji, S.; Le, L.D.; Crombleholme, T.M.; Keswani, S.G. Regenerative Wound Healing: The Role of Interleukin-10. Adv. Wound Care 2014, 3, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Kanno, E.; Tanno, H.; Masaki, A.; Sasaki, A.; Sato, N.; Goto, M.; Shisai, M.; Yamaguchi, K.; Takagi, N.; Shoji, M.; et al. Defect of Interferon gamma Leads to Impaired Wound Healing through Prolonged Neutrophilic Inflammatory Response and Enhanced MMP-2 Activation. Int. J. Mol. Sci. 2019, 20, 5657. [Google Scholar] [CrossRef]
- Tonnesen, M.G.; Feng, X.; Clark, R.A. Angiogenesis in wound healing. J. Investig. Dermatol. Symp. Proc. 2000, 5, 40–46. [Google Scholar] [CrossRef]
- Okonkwo, U.A.; DiPietro, L.A. Diabetes and Wound Angiogenesis. Int. J. Mol. Sci. 2017, 18, 1419. [Google Scholar] [CrossRef]
- Gonzalez, A.C.; Costa, T.F.; Andrade, Z.A.; Medrado, A.R. Wound healing—A literature review. An. Bras. Dermatol. 2016, 91, 614–620. [Google Scholar] [CrossRef]
- Greenhalgh, D.G. The role of apoptosis in wound healing. Int. J. Biochem. Cell Biol. 1998, 30, 1019–1030. [Google Scholar] [CrossRef]
- Cole, S.W.; Hawkley, L.C.; Arevalo, J.M.; Sung, C.Y.; Rose, R.M.; Cacioppo, J.T. Social regulation of gene expression in human leukocytes. Genome Biol. 2007, 8, R189. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.W.; Conti, G.; Arevalo, J.M.; Ruggiero, A.M.; Heckman, J.J.; Suomi, S.J. Transcriptional modulation of the developing immune system by early life social adversity. Proc. Natl. Acad. Sci. USA 2012, 109, 20578–20583. [Google Scholar] [CrossRef] [PubMed]
- Algren, M.H.; Ekholm, O.; Nielsen, L.; Ersboll, A.K.; Bak, C.K.; Andersen, P.T. Social isolation, loneliness, socioeconomic status, and health-risk behaviour in deprived neighbourhoods in Denmark: A cross-sectional study. SSM Popul. Health 2020, 10, 100546. [Google Scholar] [CrossRef] [PubMed]
- Reiss, F.; Meyrose, A.K.; Otto, C.; Lampert, T.; Klasen, F.; Ravens-Sieberer, U. Socioeconomic status, stressful life situations and mental health problems in children and adolescents: Results of the German BELLA cohort-study. PLoS ONE 2019, 14, e0213700. [Google Scholar] [CrossRef]
- Leonard, T.; Hughes, A.E.; Pruitt, S.L. Understanding how low-socioeconomic status households cope with health shocks: An analysis of multi-sector linked data. Ann. Am. Acad. Pol. Soc. Sci. 2017, 669, 125–145. [Google Scholar] [CrossRef]
- Kapp, S.; Santamaria, N. The financial and quality-of-life cost to patients living with a chronic wound in the community. Int. Wound J. 2017, 14, 1108–1119. [Google Scholar] [CrossRef]
- Nazri, N.S.; Vanoh, D.; Leng, S.K. Malnutrition, low diet quality and its risk factors among older adults with low socio-economic status: A scoping review. Nutr. Res. Rev. 2021, 34, 107–116. [Google Scholar] [CrossRef]
- Stadelmann, W.K.; Digenis, A.G.; Tobin, G.R. Impediments to wound healing. Am. J. Surg. 1998, 176, 39S–47S. [Google Scholar] [CrossRef]
- Cole, S.W. The Conserved Transcriptional Response to Adversity. Curr. Opin. Behav. Sci. 2019, 28, 31–37. [Google Scholar] [CrossRef]
- American Diabetes, A. Economic Costs of Diabetes in the U.S. in 2017. Diabetes Care 2018, 41, 917–928. [Google Scholar] [CrossRef]
- Rustagi, Y.; Abouhashem, A.S.; Verma, P.; Verma, S.S.; Hernandez, E.; Liu, S.; Kumar, M.; Guda, P.R.; Srivastava, R.; Mohanty, S.K.; et al. Endothelial Phospholipase Cgamma2 Improves Outcomes of Diabetic Ischemic Limb Rescue Following VEGF Therapy. Diabetes 2022, 71, 1149–1165. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Sinha, M.; Pal, D.; Tabasum, S.; Gnyawali, S.C.; Khona, D.; Sarkar, S.; Mohanty, S.K.; Soto-Gonzalez, F.; Khanna, S.; et al. Cutaneous Epithelial to Mesenchymal Transition Activator ZEB1 Regulates Wound Angiogenesis and Closure in a Glycemic Status-Dependent Manner. Diabetes 2019, 68, 2175–2190. [Google Scholar] [CrossRef]
- Spampinato, S.F.; Caruso, G.I.; De Pasquale, R.; Sortino, M.A.; Merlo, S. The Treatment of Impaired Wound Healing in Diabetes: Looking among Old Drugs. Pharmaceuticals 2020, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Kim, T.H.; Choi, J.Y.; Kwon, Y.J.; Choi, D.H.; Kim, K.C.; Kim, M.J.; Hwang, H.K.; Lee, K.B. Predictors for Amputation in Patients with Diabetic Foot Wound. Vasc. Specialist Int. 2018, 34, 109–116. [Google Scholar] [CrossRef]
- Singh, K.; Agrawal, N.K.; Gupta, S.K.; Sinha, P.; Singh, K. Increased expression of TLR9 associated with pro-inflammatory S100A8 and IL-8 in diabetic wounds could lead to unresolved inflammation in type 2 diabetes mellitus (T2DM) cases with impaired wound healing. J. Diabetes Complicat. 2016, 30, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Agrawal, N.K.; Gupta, S.K.; Mohan, G.; Chaturvedi, S.; Singh, K. Genetic and epigenetic alterations in Toll like receptor 2 and wound healing impairment in type 2 diabetes patients. J. Diabetes Complicat. 2015, 29, 222–229. [Google Scholar] [CrossRef]
- Singh, K.; Singh, V.K.; Agrawal, N.K.; Gupta, S.K.; Singh, K. Genetic Alterations in Toll-Like Receptor 4 Signaling Pathway and Impairment of Wound Healing in Patients With Type 2 Diabetes. Int. J. Low Extrem. Wounds 2014, 13, 162–163. [Google Scholar] [CrossRef]
- Pal, D.; Ghatak, S.; Singh, K.; Abouhashem, A.S.; Kumar, M.; El Masry, M.S.; Mohanty, S.K.; Palakurti, R.; Rustagi, Y.; Tabasum, S.; et al. Identification of a physiologic vasculogenic fibroblast state to achieve tissue repair. Nat. Commun. 2023, 14, 1129. [Google Scholar] [CrossRef]
- Sinha, M.; Sen, C.K.; Singh, K.; Das, A.; Ghatak, S.; Rhea, B.; Blackstone, B.; Powell, H.M.; Khanna, S.; Roy, S. Direct conversion of injury-site myeloid cells to fibroblast-like cells of granulation tissue. Nat. Commun. 2018, 9, 936. [Google Scholar] [CrossRef]
- Anderson, K.; Hamm, R.L. Factors That Impair Wound Healing. J. Am. Coll. Clin. Wound Spec. 2012, 4, 84–91. [Google Scholar] [CrossRef]
- Singh, K.; Agrawal, N.K.; Gupta, S.K.; Mohan, G.; Chaturvedi, S.; Singh, K. Differential Expression of Matrix Metalloproteinase-9 Gene in Wounds of Type 2 Diabetes Mellitus Cases with Susceptible-1562C>T Genotypes and Wound Severity. Int. J. Low Extrem. Wounds 2014, 13, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Singh, K. Carcinogenesis and diabetic wound healing: Evidences of parallelism. Curr. Diabetes Rev. 2015, 11, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yang, C.; Wang, X.Y.; Zhou, L.Y.; Lao, G.J.; Liu, D.; Wang, C.; Hu, M.D.; Zeng, T.T.; Yan, L.; et al. MicroRNA-129 and -335 Promote Diabetic Wound Healing by Inhibiting Sp1-Mediated MMP-9 Expression. Diabetes 2018, 67, 1627–1638. [Google Scholar] [CrossRef] [PubMed]
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; Castruita-De la Rosa, C.; Ramirez-Acuna, J.M.; Perez-Romero, B.A.; Guerrero-Rodriguez, J.F.; Martinez-Avila, N.; Martinez-Fierro, M.L. The Roles of Matrix Metalloproteinases and Their Inhibitors in Human Diseases. Int. J. Mol. Sci. 2020, 21, 9739. [Google Scholar] [CrossRef]
- Chawla, M.; Gupta, R.; Das, B. Gut microbiome dysbiosis in malnutrition. Prog. Mol. Biol. Transl. Sci. 2022, 192, 205–229. [Google Scholar]
- de Oliveira, G.L.V.; Cardoso, C.R.B.; Taneja, V.; Fasano, A. Editorial: Intestinal Dysbiosis in Inflammatory Diseases. Front. Immunol. 2021, 12, 727485. [Google Scholar] [CrossRef]
- Patterson, G.T.; Osorio, E.Y.; Peniche, A.; Dann, S.M.; Cordova, E.; Preidis, G.A.; Suh, J.H.; Ito, I.; Saldarriaga, O.A.; Loeffelholz, M.; et al. Pathologic Inflammation in Malnutrition Is Driven by Proinflammatory Intestinal Microbiota, Large Intestine Barrier Dysfunction, and Translocation of Bacterial Lipopolysaccharide. Front. Immunol. 2022, 13, 846155. [Google Scholar] [CrossRef]
- Stechmiller, J.K. Understanding the role of nutrition and wound healing. Nutr. Clin. Pract. 2010, 25, 61–68. [Google Scholar] [CrossRef]
- Kiani, A.K.; Bonetti, G.; Donato, K.; Kaftalli, J.; Herbst, K.L.; Stuppia, L.; Fioretti, F.; Nodari, S.; Perrone, M.; Chiurazzi, P.; et al. Polymorphisms, diet and nutrigenomics. J. Prev. Med. Hyg. 2022, 63, E125–E141. [Google Scholar]
- Schwartzfarb, E.M.; Romanelli, P. Hyperhomocysteinemia and lower extremity wounds. Int. J. Low Extrem. Wounds 2008, 7, 126–136. [Google Scholar] [CrossRef]
- Moll, S.; Varga, E.A. Homocysteine and MTHFR Mutations. Circulation 2015, 132, e6–e9. [Google Scholar] [CrossRef] [PubMed]
- New, D.; Eaton, P.; Knable, A.; Callen, J.P. The use of B vitamins for cutaneous ulcerations mimicking pyoderma gangrenosum in patients with MTHFR polymorphism. Arch. Dermatol. 2011, 147, 450–453. [Google Scholar] [CrossRef] [PubMed]
- Dedoussis, G.V.; Panagiotakos, D.B.; Pitsavos, C.; Chrysohoou, C.; Skoumas, J.; Choumerianou, D.; Stefanadis, C.; Group, A.S. An association between the methylenetetrahydrofolate reductase (MTHFR) C677T mutation and inflammation markers related to cardiovascular disease. Int. J. Cardiol. 2005, 100, 409–414. [Google Scholar] [CrossRef]
- Boykin, J.; Baylis, C. Homocysteine—A stealth mediator of impaired wound healing: A preliminary study. Wounds 2006, 18, 101–114. [Google Scholar]
- Frosst, P.; Blom, H.J.; Milos, R.; Goyette, P.; Sheppard, C.A.; Matthews, R.G.; Boers, G.J.; den Heijer, M.; Kluijtmans, L.A.; van den Heuvel, L.P.; et al. A candidate genetic risk factor for vascular disease: A common mutation in methylenetetrahydrofolate reductase. Nat. Genet. 1995, 10, 111–113. [Google Scholar] [CrossRef]
- Quigley, D.A.; To, M.D.; Perez-Losada, J.; Pelorosso, F.G.; Mao, J.H.; Nagase, H.; Ginzinger, D.G.; Balmain, A. Genetic architecture of mouse skin inflammation and tumour susceptibility. Nature 2009, 458, 505–508. [Google Scholar] [CrossRef]
- Oda, Y.; Tu, C.L.; Menendez, A.; Nguyen, T.; Bikle, D.D. Vitamin D and calcium regulation of epidermal wound healing. J. Steroid Biochem. Mol. Biol. 2016, 164, 379–385. [Google Scholar] [CrossRef]
- Lu, X.; Vick, S.; Chen, Z.; Chen, J.; Watsky, M.A. Effects of Vitamin D Receptor Knockout and Vitamin D Deficiency on Corneal Epithelial Wound Healing and Nerve Density in Diabetic Mice. Diabetes 2020, 69, 1042–1051. [Google Scholar] [CrossRef]
- Burkiewicz, C.J.; Guadagnin, F.A.; Skare, T.L.; do Nascimento, M.M.; Servin, S.C.; de Souza, G.D. Vitamin D and skin repair: A prospective, double-blind and placebo controlled study in the healing of leg ulcers. Rev. Col. Bras. Cir. 2012, 39, 401–407. [Google Scholar] [CrossRef]
- Kalava, U.R.; Cha, S.S.; Takahashi, P.Y. Association between vitamin D and pressure ulcers in older ambulatory adults: Results of a matched case-control study. Clin. Interv. Aging 2011, 6, 213–219. [Google Scholar]
- Bashutski, J.D.; Eber, R.M.; Kinney, J.S.; Benavides, E.; Maitra, S.; Braun, T.M.; Giannobile, W.V.; McCauley, L.K. The impact of vitamin D status on periodontal surgery outcomes. J. Dent. Res. 2011, 90, 1007–1012. [Google Scholar] [CrossRef] [PubMed]
- Smart, H.; AlGhareeb, A.M.; Smart, S.A. 25-Hydroxyvitamin D Deficiency: Impacting Deep-Wound Infection and Poor Healing Outcomes in Patients With Diabetes. Adv. Skin Wound Care 2019, 32, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Razzaghi, R.; Pourbagheri, H.; Momen-Heravi, M.; Bahmani, F.; Shadi, J.; Soleimani, Z.; Asemi, Z. The effects of vitamin D supplementation on wound healing and metabolic status in patients with diabetic foot ulcer: A randomized, double-blind, placebo-controlled trial. J. Diabetes Complicat. 2017, 31, 766–772. [Google Scholar] [CrossRef]
- Shaat, N.; Katsarou, A.; Shahida, B.; Prasad, R.B.; Kristensen, K.; Planck, T. Association between the rs1544410 polymorphism in the vitamin D receptor (VDR) gene and insulin secretion after gestational diabetes mellitus. PLoS ONE 2020, 15, e0232297. [Google Scholar] [CrossRef] [PubMed]
- Sharif, E.; Swaidan, N.; Shurbaji, S.; Rizk, N. Associations of vitamin D receptor polymorphism rs1544410 with adiposity phenotypes. Endocrinol. Metab. Int. J. 2016, 3, 165–172. [Google Scholar]
- Salo, T.; Makela, M.; Kylmaniemi, M.; Autio-Harmainen, H.; Larjava, H. Expression of matrix metalloproteinase-2 and -9 during early human wound healing. Lab. Investig 1994, 70, 176–182. [Google Scholar]
- Cho, H.; Balaji, S.; Hone, N.L.; Moles, C.M.; Sheikh, A.Q.; Crombleholme, T.M.; Keswani, S.G.; Narmoneva, D.A. Diabetic wound healing in a MMP9-/- mouse model. Wound Repair Regen. 2016, 24, 829–840. [Google Scholar] [CrossRef]
- Serra, R.; Buffone, G.; Falcone, D.; Molinari, V.; Scaramuzzino, M.; Gallelli, L.; de Franciscis, S. Chronic venous leg ulcers are associated with high levels of metalloproteinases-9 and neutrophil gelatinase-associated lipocalin. Wound Repair Regen. 2013, 21, 395–401. [Google Scholar] [CrossRef]
- Muller, M.; Trocme, C.; Lardy, B.; Morel, F.; Halimi, S.; Benhamou, P.Y. Matrix metalloproteinases and diabetic foot ulcers: The ratio of MMP-1 to TIMP-1 is a predictor of wound healing. Diabet. Med. 2008, 25, 419–426. [Google Scholar] [CrossRef]
- Rayment, E.A.; Upton, Z.; Shooter, G.K. Increased matrix metalloproteinase-9 (MMP-9) activity observed in chronic wound fluid is related to the clinical severity of the ulcer. Br. J. Dermatol. 2008, 158, 951–961. [Google Scholar] [CrossRef]
- Hernandez-Anzaldo, S.; Brglez, V.; Hemmeryckx, B.; Leung, D.; Filep, J.G.; Vance, J.E.; Vance, D.E.; Kassiri, Z.; Lijnen, R.H.; Lambeau, G.; et al. Novel Role for Matrix Metalloproteinase 9 in Modulation of Cholesterol Metabolism. J. Am. Heart Assoc. 2016, 5, e004228. [Google Scholar] [CrossRef] [PubMed]
- Aljada, A.; Ghanim, H.; Mohanty, P.; Syed, T.; Bandyopadhyay, A.; Dandona, P. Glucose intake induces an increase in activator protein 1 and early growth response 1 binding activities, in the expression of tissue factor and matrix metalloproteinase in mononuclear cells, and in plasma tissue factor and matrix metalloproteinase concentrations. Am. J. Clin. Nutr. 2004, 80, 51–57. [Google Scholar] [PubMed]
- Watson, C.; Spiers, J.P.; Waterstone, M.; Russell-Hallinan, A.; Gallagher, J.; McDonald, K.; Ryan, C.; Gilmer, J.; Ledwidge, M. Investigation of association of genetic variant rs3918242 of matrix metalloproteinase-9 with hypertension, myocardial infarction and progression of ventricular dysfunction in Irish Caucasian patients with diabetes: A report from the STOP-HF follow-up programme. BMC Cardiovasc. Disord. 2021, 21, 87. [Google Scholar]
- Perunovic, N.; Rakic, M.; Jankovic, S.; Aleksic, Z.; Struillou, X.; Cakic, S.; Puletic, M.; Lekovic, V.; Milasin, J. MMP-9 -1562 C>T (rs3918242) promoter polymorphism as a susceptibility factor for multiple gingival recessions. Int. J. Periodontics Restor. Dent. 2015, 35, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, Y.; Zhao, L. MMP-9 gene rs3918242 polymorphism increases risk of stroke: A meta-analysis. J. Cell Biochem. 2018, 119, 9801–9808. [Google Scholar] [CrossRef]
- Singh, K.; Agrawal, N.K.; Gupta, S.K.; Singh, K. A functional single nucleotide polymorphism -1562C>T in the matrix metalloproteinase-9 promoter is associated with type 2 diabetes and diabetic foot ulcers. Int. J. Low Extrem. Wounds 2013, 12, 199–204. [Google Scholar] [CrossRef]
- Marginean, C.O.; Marginean, C.; Banescu, C.; Melit, L.E.; Tripon, F.; Iancu, M. The relationship between MMP9 and ADRA2A gene polymorphisms and mothers-newborns’ nutritional status: An exploratory path model (STROBE compliant article). Pediatr. Res. 2019, 85, 822–829. [Google Scholar] [CrossRef]
- Munoz, N.; Litchford, M.; Cereda, E. Nutrition and Wound Care. Phys. Med. Rehabil. Clin. N. Am. 2022, 33, 811–822. [Google Scholar] [CrossRef]
- Subramaniam, T.; Fauzi, M.B.; Lokanathan, Y.; Law, J.X. The Role of Calcium in Wound Healing. Int. J. Mol. Sci. 2021, 22, 6486. [Google Scholar] [CrossRef]
- Simes, D.C.; Viegas, C.S.B.; Araujo, N.; Marreiros, C. Vitamin K as a Powerful Micronutrient in Aging and Age-Related Diseases: Pros and Cons from Clinical Studies. Int. J. Mol. Sci. 2019, 20, 4150. [Google Scholar] [CrossRef]
- Pazyar, N.; Houshmand, G.; Yaghoobi, R.; Hemmati, A.A.; Zeineli, Z.; Ghorbanzadeh, B. Wound healing effects of topical Vitamin K: A randomized controlled trial. Indian J. Pharmacol. 2019, 51, 88–92. [Google Scholar]
- Barchitta, M.; Maugeri, A.; Favara, G.; Magnano San Lio, R.; Evola, G.; Agodi, A.; Basile, G. Nutrition and Wound Healing: An Overview Focusing on the Beneficial Effects of Curcumin. Int. J. Mol. Sci. 2019, 20, 1119. [Google Scholar] [CrossRef] [PubMed]
- Wild, T.; Rahbarnia, A.; Kellner, M.; Sobotka, L.; Eberlein, T. Basics in nutrition and wound healing. Nutrition 2010, 26, 862–866. [Google Scholar] [CrossRef]
- McCarty, M.F. Glucosamine for wound healing. Med. Hypotheses 1996, 47, 273–275. [Google Scholar] [CrossRef]
- Ju, M.; Kim, Y.; Seo, K.W. Role of nutrition in wound healing and nutritional recommendations for promotion of wound healing: A narrative review. Ann. Clin. Nutr. Metab. 2023, 15, 67–71. [Google Scholar] [CrossRef]
- Demling, R.H. Nutrition, anabolism, and the wound healing process: An overview. Eplasty 2009, 9, e9. [Google Scholar] [PubMed]
- Guardiola-Marquez, C.E.; Santos-Ramirez, M.T.; Segura-Jimenez, M.E.; Figueroa-Montes, M.L.; Jacobo-Velazquez, D.A. Fighting Obesity-Related Micronutrient Deficiencies through Biofortification of Agri-Food Crops with Sustainable Fertilization Practices. Plants 2022, 11, 3477. [Google Scholar] [CrossRef]
- MacKay, D.; Miller, A.L. Nutritional support for wound healing. Altern. Med. Rev. 2003, 8, 359–377. [Google Scholar]
- Molnar, J.A.; Underdown, M.J.; Clark, W.A. Nutrition and Chronic Wounds. Adv. Wound Care 2014, 3, 663–681. [Google Scholar] [CrossRef]
- Russell, L. The importance of patients’ nutritional status in wound healing. Br. J. Nurs. 2001, 10, S42–S49. [Google Scholar] [CrossRef]
- Alves, A.; Morio, B. Alterations in glycine metabolism in obesity and chronic metabolic diseases—An update on new advances. Curr. Opin. Clin. Nutr. Metab. Care 2023, 26, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Shu, X.L.; Zhong, J.X.; Yu, T.T. Effect of L-arginine on immune function: A meta-analysis. Asia Pac. J. Clin. Nutr. 2014, 23, 351–359. [Google Scholar] [PubMed]
- Stechmiller, J.K.; Childress, B.; Cowan, L. Arginine supplementation and wound healing. Nutr. Clin. Pract. 2005, 20, 52–61. [Google Scholar] [CrossRef]
- Ellinger, S. Micronutrients, Arginine, and Glutamine: Does Supplementation Provide an Efficient Tool for Prevention and Treatment of Different Kinds of Wounds? Adv. Wound Care 2014, 3, 691–707. [Google Scholar] [CrossRef] [PubMed]
- Blanc, M.C.; Moinard, C.; Beziel, A.; Darquy, S.; Cynober, L.; De Bandt, J.P. Arginine and glutamine availability and macrophage functions in the obese insulin-resistant Zucker rat. J. Cell Physiol. 2005, 202, 153–159. [Google Scholar] [CrossRef]
- Zinder, R.; Cooley, R.; Vlad, L.G.; Molnar, J.A. Vitamin A and Wound Healing. Nutr. Clin. Pract. 2019, 34, 839–849. [Google Scholar] [CrossRef]
- Hof, H. Vitamin A, the “anti-infective” vitamin? (author’s transl). MMW Munch. Med. Wochenschr. 1976, 118, 1485–1488. [Google Scholar]
- Trasino, S.E.; Tang, X.H.; Jessurun, J.; Gudas, L.J. Obesity Leads to Tissue, but not Serum Vitamin A Deficiency. Sci. Rep. 2015, 5, 15893. [Google Scholar] [CrossRef]
- Hanna, M.; Jaqua, E.; Nguyen, V.; Clay, J. B Vitamins: Functions and Uses in Medicine. Perm. J. 2022, 26, 89–97. [Google Scholar] [CrossRef]
- Mochizuki, S.; Takano, M.; Sugano, N.; Ohtsu, M.; Tsunoda, K.; Koshi, R.; Yoshinuma, N. The effect of B vitamin supplementation on wound healing in type 2 diabetic mice. J. Clin. Biochem. Nutr. 2016, 58, 64–68. [Google Scholar] [CrossRef]
- Bosse, M.D.; Axelrod, A.E. Wound healing in rats with biotin, pyridoxin, or riboflavin deficiencies. Proc. Soc. Exp. Biol. Med. 1948, 67, 418–421. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Sun, M.; Liu, B.; Du, Y.; Rong, S.; Xu, G.; Snetselaar, L.G.; Bao, W. Inverse Association Between Serum Vitamin B12 Concentration and Obesity Among Adults in the United States. Front. Endocrinol. 2019, 10, 414. [Google Scholar] [CrossRef]
- Moores, J. Vitamin C: A wound healing perspective. Br. J. Community Nurs. 2013, 18 (Suppl. S6), S8–S11. [Google Scholar] [CrossRef]
- Johnston, C.S.; Beezhold, B.L.; Mostow, B.; Swan, P.D. Plasma vitamin C is inversely related to body mass index and waist circumference but not to plasma adiponectin in nonsmoking adults. J. Nutr. 2007, 137, 1757–1762. [Google Scholar] [CrossRef]
- Arnold, M.; Barbul, A. Nutrition and wound healing. Plast Reconstr. Surg. 2006, 117, 42S–58S. [Google Scholar] [CrossRef] [PubMed]
- Grundmann, M.; Haidar, M.; Placzko, S.; Niendorf, R.; Darashchonak, N.; Hubel, C.A.; von Versen-Hoynck, F. Vitamin D improves the angiogenic properties of endothelial progenitor cells. Am. J. Physiol. Cell Physiol. 2012, 303, C954–C962. [Google Scholar] [CrossRef]
- Youssef, D.A.; Miller, C.W.; El-Abbassi, A.M.; Cutchins, D.C.; Cutchins, C.; Grant, W.B.; Peiris, A.N. Antimicrobial implications of vitamin D. Dermatoendocrinol 2011, 3, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, W.; Wang, Y.; Han, X.; Gao, L. Vitamin D Receptor Polymorphisms Associated with Susceptibility to Obesity: A Meta-Analysis. Med. Sci. Monit. 2019, 25, 8297–8305. [Google Scholar] [CrossRef]
- Vranic, L.; Mikolasevic, I.; Milic, S. Vitamin D Deficiency: Consequence or Cause of Obesity? Medicina 2019, 55, 541. [Google Scholar] [CrossRef]
- Al-Daghri, N.M.; Guerini, F.R.; Al-Attas, O.S.; Alokail, M.S.; Alkharfy, K.M.; Draz, H.M.; Agliardi, C.; Costa, A.S.; Saulle, I.; Mohammed, A.K.; et al. Vitamin D receptor gene polymorphisms are associated with obesity and inflammosome activity. PLoS ONE 2014, 9, e102141. [Google Scholar] [CrossRef]
- Guo, S.; Dipietro, L.A. Factors affecting wound healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Garcia, O.P.; Ronquillo, D.; del Carmen Caamano, M.; Martinez, G.; Camacho, M.; Lopez, V.; Rosado, J.L. Zinc, iron and vitamins A, C and e are associated with obesity, inflammation, lipid profile and insulin resistance in Mexican school-aged children. Nutrients 2013, 5, 5012–5030. [Google Scholar] [CrossRef] [PubMed]
- Tie, J.K.; Stafford, D.W. Structural and functional insights into enzymes of the vitamin K cycle. J. Thromb. Haemost. 2016, 14, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Ruan, Z.; Ma, A.; Wang, D.; Kou, J. Effect of vitamin K on wound healing: A systematic review and meta-analysis based on preclinical studies. Front. Pharmacol. 2022, 13, 1063349. [Google Scholar] [CrossRef] [PubMed]
- Shea, M.K.; Booth, S.L.; Gundberg, C.M.; Peterson, J.W.; Waddell, C.; Dawson-Hughes, B.; Saltzman, E. Adulthood obesity is positively associated with adipose tissue concentrations of vitamin K and inversely associated with circulating indicators of vitamin K status in men and women. J. Nutr. 2010, 140, 1029–1034. [Google Scholar] [CrossRef]
- Silva, J.R.; Burger, B.; Kuhl, C.M.C.; Candreva, T.; Dos Anjos, M.B.P.; Rodrigues, H.G. Wound Healing and Omega-6 Fatty Acids: From Inflammation to Repair. Mediat. Inflamm. 2018, 2018, 2503950. [Google Scholar] [CrossRef]
- Simopoulos, A.P. An Increase in the Omega-6/Omega-3 Fatty Acid Ratio Increases the Risk for Obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef]
- Wright, J.A.; Richards, T.; Srai, S.K. The role of iron in the skin and cutaneous wound healing. Front. Pharmacol. 2014, 5, 156. [Google Scholar] [CrossRef]
- Alshwaiyat, N.M.; Ahmad, A.; Wan Hassan, W.M.R.; Al-Jamal, H.A.N. Association between obesity and iron deficiency (Review). Exp. Ther. Med. 2021, 22, 1268. [Google Scholar] [CrossRef]
- Hamoui, N.; Anthone, G.; Crookes, P.F. Calcium metabolism in the morbidly obese. Obes. Surg. 2004, 14, 9–12. [Google Scholar] [CrossRef]
- Lansdown, A.B. Calcium: A potential central regulator in wound healing in the skin. Wound Repair Regen. 2002, 10, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Lansdown, A.B.; Mirastschijski, U.; Stubbs, N.; Scanlon, E.; Agren, M.S. Zinc in wound healing: Theoretical, experimental, and clinical aspects. Wound Repair Regen. 2007, 15, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.H.; Sermersheim, M.; Li, H.; Lee, P.H.U.; Steinberg, S.M.; Ma, J. Zinc in Wound Healing Modulation. Nutrients 2017, 10, 16. [Google Scholar] [CrossRef]
- Liu, M.J.; Bao, S.; Bolin, E.R.; Burris, D.L.; Xu, X.; Sun, Q.; Killilea, D.W.; Shen, Q.; Ziouzenkova, O.; Belury, M.A.; et al. Zinc deficiency augments leptin production and exacerbates macrophage infiltration into adipose tissue in mice fed a high-fat diet. J. Nutr. 2013, 143, 1036–1045. [Google Scholar] [CrossRef]
- Watford, M.; Wu, G. Protein. Adv. Nutr. 2018, 9, 651–653. [Google Scholar] [CrossRef]
- Breslow, R.A.; Hallfrisch, J.; Guy, D.G.; Crawley, B.; Goldberg, A.P. The importance of dietary protein in healing pressure ulcers. J. Am. Geriatr. Soc. 1993, 41, 357–362. [Google Scholar] [CrossRef]
- Arribas-Lopez, E.; Zand, N.; Ojo, O.; Snowden, M.J.; Kochhar, T. The Effect of Amino Acids on Wound Healing: A Systematic Review and Meta-Analysis on Arginine and Glutamine. Nutrients 2021, 13, 2498. [Google Scholar] [CrossRef]
- Albaugh, V.L.; Mukherjee, K.; Barbul, A. Proline Precursors and Collagen Synthesis: Biochemical Challenges of Nutrient Supplementation and Wound Healing. J. Nutr. 2017, 147, 2011–2017. [Google Scholar] [CrossRef] [PubMed]
- Jara, C.P.; Mendes, N.F.; Prado, T.P.D.; de Araujo, E.P. Bioactive Fatty Acids in the Resolution of Chronic Inflammation in Skin Wounds. Adv. Wound Care 2020, 9, 472–490. [Google Scholar] [CrossRef]
- Astrup, A.; Teicholz, N.; Magkos, F.; Bier, D.M.; Brenna, J.T.; King, J.C.; Mente, A.; Ordovas, J.M.; Volek, J.S.; Yusuf, S.; et al. Dietary Saturated Fats and Health: Are the U.S. Guidelines Evidence-Based? Nutrients 2021, 13, 3305. [Google Scholar] [CrossRef]
- Coniglio, S.; Shumskaya, M.; Vassiliou, E. Unsaturated Fatty Acids and Their Immunomodulatory Properties. Biology 2023, 12, 279. [Google Scholar] [CrossRef] [PubMed]
- Rosa, D.F.; Sarandy, M.M.; Novaes, R.D.; da Matta, S.L.P.; Goncalves, R.V. Effect of a high-fat diet and alcohol on cutaneous repair: A systematic review of murine experimental models. PLoS ONE 2017, 12, e0176240. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, K.L. The science of fatty acids and inflammation. Adv. Nutr. 2015, 6, 293S–301S. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.W.; Supp, D.M. Role of Arginine and Omega-3 Fatty Acids in Wound Healing and Infection. Adv. Wound Care 2014, 3, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Puddu, P.E.; Menotti, A. Simple versus complex carbohydrates and health: A frequently neglected problem. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 1949–1952. [Google Scholar] [CrossRef] [PubMed]
- Akbar, A.; Shreenath, A.P. High Fiber Diet; StatPearls: Treasure Island, FL, USA, 2024. [Google Scholar]
- Canesso, M.C.C.; Cassini-Vieira, P.; Moreira, C.F.; Luong, S.; Rachid, M.A.; Martins, F.S.; Teixeira, M.M.; Vieira, A.T.; Mackay, C.R.; Barcelos, L.S. Dietary Fiber Improves Skin Wound Healing and Scar Formation through the Metabolite-Sensing Receptor GPR43. J. Investig. Dermatol. 2023, 143, 1850–1854.e6. [Google Scholar] [CrossRef]
- Waghmare, V.S.; Wadke, P.R.; Dyawanapelly, S.; Deshpande, A.; Jain, R.; Dandekar, P. Starch based nanofibrous scaffolds for wound healing applications. Bioact. Mater. 2018, 3, 255–266. [Google Scholar] [CrossRef]
- Seth, I.; Lim, B.; Cevik, J.; Gracias, D.; Chua, M.; Kenney, P.S.; Rozen, W.M.; Cuomo, R. Impact of nutrition on skin wound healing and aesthetic outcomes: A comprehensive narrative review. JPRAS Open 2024, 39, 291–302. [Google Scholar] [CrossRef]
- Nelson, R.L. The utility of pharmacokinetics to the pharmaceutical industry. J. Clin. Pharmacol. 1976, 16, 565–569. [Google Scholar] [CrossRef]
- Polcz, M.E.; Barbul, A. The Role of Vitamin A in Wound Healing. Nutr. Clin. Pract. 2019, 34, 695–700. [Google Scholar] [CrossRef]
- Rembe, J.D.; Fromm-Dornieden, C.; Stuermer, E.K. Effects of Vitamin B Complex and Vitamin C on Human Skin Cells: Is the Perceived Effect Measurable? Adv. Skin Wound Care 2018, 31, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Pullar, J.M.; Carr, A.C.; Vissers, M.C.M. The Roles of Vitamin C in Skin Health. Nutrients. 2017, 9, 866. [Google Scholar] [CrossRef] [PubMed]
- Mustacich, D.J.; Bruno, R.S.; Traber, M.G. Vitamin E. Vitam. Horm. 2007, 76, 1–21. [Google Scholar]
- Tanaydin, V.; Conings, J.; Malyar, M.; van der Hulst, R.; van der Lei, B. The Role of Topical Vitamin E in Scar Management: A Systematic Review. Aesthet. Surg. J. 2016, 36, 959–965. [Google Scholar] [CrossRef]
- Kogan, S.; Sood, A.; Garnick, M.S. Zinc and Wound Healing: A Review of Zinc Physiology and Clinical Applications. Wounds 2017, 29, 102–106. [Google Scholar] [PubMed]
- Sakae, K.; Agata, T.; Kamide, R.; Yanagisawa, H. Effects of L-carnosine and its zinc complex (Polaprezinc) on pressure ulcer healing. Nutr. Clin. Pract. 2013, 28, 609–616. [Google Scholar] [CrossRef]
- Bauer, J.D.; Isenring, E.; Waterhouse, M. The effectiveness of a specialised oral nutrition supplement on outcomes in patients with chronic wounds: A pragmatic randomised study. J. Hum. Nutr. Diet. 2013, 26, 452–458. [Google Scholar] [CrossRef]
- Bechara, N.; Flood, V.M.; Gunton, J.E. A Systematic Review on the Role of Vitamin C in Tissue Healing. Antioxidants 2022, 11, 1605. [Google Scholar] [CrossRef] [PubMed]
- Gunton, J.E.; Girgis, C.M.; Lau, T.; Vicaretti, M.; Begg, L.; Flood, V. Vitamin C improves healing of foot ulcers: A randomised, double-blind, placebo-controlled trial. Br. J. Nutr. 2021, 126, 1451–1458. [Google Scholar] [CrossRef]
- Taylor, T.V.; Rimmer, S.; Day, B.; Butcher, J.; Dymock, I.W. Ascorbic acid supplementation in the treatment of pressure-sores. Lancet 1974, 2, 544–546. [Google Scholar] [CrossRef]
- Song, Y.P.; Wang, L.; Yu, H.R.; Yuan, B.F.; Shen, H.W.; Du, L.; Cai, J.Y.; Chen, H.L. Zinc Therapy Is a Reasonable Choice for Patients With Pressure Injuries: A Systematic Review and Meta-Analysis. Nutr. Clin. Pract. 2020, 35, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Desneves, K.J.; Todorovic, B.E.; Cassar, A.; Crowe, T.C. Treatment with supplementary arginine, vitamin C and zinc in patients with pressure ulcers: A randomised controlled trial. Clin. Nutr. 2005, 24, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Cereda, E.; Klersy, C.; Serioli, M.; Crespi, A.; D’Andrea, F.; OligoElement Sore Trial Study, G. A nutritional formula enriched with arginine, zinc, and antioxidants for the healing of pressure ulcers: A randomized trial. Ann. Intern. Med. 2015, 162, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Rindone, J.P.; Hiller, D.; Collacott, E.; Nordhaugen, N.; Arriola, G. Randomized, controlled trial of glucosamine for treating osteoarthritis of the knee. West. J. Med. 2000, 172, 91–94. [Google Scholar] [CrossRef]
- Iyengar, N.M.; Gucalp, A.; Dannenberg, A.J.; Hudis, C.A. Obesity and Cancer Mechanisms: Tumor Microenvironment and Inflammation. J. Clin. Oncol. 2016, 34, 4270–4276. [Google Scholar] [CrossRef]
- Wagner, I.J.; Szpalski, C.; Allen, R.J., Jr.; Davidson, E.H.; Canizares, O.; Saadeh, P.B.; Warren, S.M. Obesity impairs wound closure through a vasculogenic mechanism. Wound Repair Regen. 2012, 20, 512–522. [Google Scholar] [CrossRef]
- Rodriguez, P.G.; Felix, F.N.; Woodley, D.T.; Shim, E.K. The role of oxygen in wound healing: A review of the literature. Dermatol. Surg. 2008, 34, 1159–1169. [Google Scholar] [CrossRef]
- Hong, W.X.; Hu, M.S.; Esquivel, M.; Liang, G.Y.; Rennert, R.C.; McArdle, A.; Paik, K.J.; Duscher, D.; Gurtner, G.C.; Lorenz, H.P.; et al. The Role of Hypoxia-Inducible Factor in Wound Healing. Adv. Wound Care 2014, 3, 390–399. [Google Scholar] [CrossRef]
- Emery, C.F.; Kiecolt-Glaser, J.K.; Glaser, R.; Malarkey, W.B.; Frid, D.J. Exercise accelerates wound healing among healthy older adults: A preliminary investigation. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 1432–1436. [Google Scholar] [CrossRef]
- Yang, D.J.; Moh, S.H.; Choi, Y.H.; Kim, K.W. Beta-Neoendorphin Enhances Wound Healing by Promoting Cell Migration in Keratinocyte. Molecules 2020, 25, 4640. [Google Scholar] [CrossRef]
- Goldfarb, A.H.; Jamurtas, A.Z. Beta-endorphin response to exercise. An update. Sports Med. 1997, 24, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Pence, B.D.; Woods, J.A. Exercise, Obesity, and Cutaneous Wound Healing: Evidence from Rodent and Human Studies. Adv. Wound Care 2014, 3, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Bolton, L. Exercise and Chronic Wound Healing. Wounds 2019, 31, 65–67. [Google Scholar] [PubMed]
- Eraydin, S.; Avsar, G. The Effect of Foot Exercises on Wound Healing in Type 2 Diabetic Patients With a Foot Ulcer: A Randomized Control Study. J. Wound Ostomy Cont. Nurs. 2018, 45, 123–130. [Google Scholar] [CrossRef]
- Drake, D.J.; Swanson, M.; Baker, G.; Pokorny, M.; Rose, M.A.; Clark-Reed, L.; Waters, W.; Watkins, F.R., Jr.; Engelke, M.K. The association of BMI and Braden total score on the occurrence of pressure ulcers. J. Wound Ostomy Cont. Nurs. 2010, 37, 367–371. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Socioeconomic Factor | Low SES Predictor | Impact on Chronic Wounds |
|---|---|---|
| Social Status | Prolonged social isolation [74] | Upregulation of pro-inflammatory mechanisms; disruption of wound healing stages [64,72] |
| Childhood adversity [75] | Heightened pro-inflammatory processes, altered leukocyte gene expression; disruption of wound healing process [64,73] | |
| Financial Status | Lower income—diminished access to healthcare [76] | Chronic wound susceptibility and aggressiveness [13,77] |
| Nutritional status | Poor diet and malnutrition [78] | Delayed and improper wound healing at all stages [79] |
| Educational Status | Lower educational attainment—increased risk-taking behaviors like cigarette smoking [21] | Bodily health detriment, ischemic consequences in wound healing [20,22] |
| Macronutrients/Micronutrients | Major Wound Healing Functions | Obesity; Negative Consequence |
|---|---|---|
| Amino Acids | ||
| Proline and Glycine | Building blocks of collagen, wound proliferation, and remodeling stages [70,141] | Decreased serum levels; wound dehiscence and impaired regeneration [71,142] |
| Arginine and Glutamine | Cell proliferation, nitric oxide (NO) and collagen synthesis, angiogenesis, inflammation control [143,144] | Abnormal cellular metabolism and signaling pathways; impaired wound-breaking strength [145,146] |
| Vitamins | ||
| Vitamin A | Epithelization and fibroplasia, anti-infection properties [147,148] | Impaired signaling pathways in tissues and organs; associated with increased wound infection susceptibility [149] |
| Vitamin B | Energy production, red blood cell synthesis, co-factor in various metabolic pathways [150] | Decreased serum levels; delayed wound contraction, remodeling, and impaired regeneration [151,152,153] |
| Vitamin C | Key role in collagen synthesis, maturation, and secretion, antioxidant properties [154] | Serum levels are inversely related to BMI; impaired proliferation and remodeling stages; decreased angiogenesis [155,156] |
| Vitamin D | Anti-infection properties, epidermal growth factors, angiogenesis [157,158] | Strongly correlated deficiency, polymorphisms in vitamin D receptor (VDR) gene; systemic inflammation and impaired healing [159,160,161] |
| Vitamin E | Anti-inflammatory and antioxidant properties, cellular membrane integrity [162] | Decreased serum levels; can prolong inflammatory response and lead to excessive scarring [163] |
| Vitamin K | Significant role in the blood coagulation cascade [164] | Decreased serum circulatory levels; reduced clotting in inflammatory response, delayed healing [165,166] |
| Fatty Acids | ||
| Omega-3 and Omega-6 fatty acids (unsaturated fats) | Roles in all phases of wound healing, anti-inflammatory processes [167] | Increased levels; can induce chronic systemic inflammation [168] |
| Minerals | ||
| Iron | Oxygen delivery, collagen synthesis, epithelization processes [169] | Disruption of iron homeostasis; ischemic consequences in wounds [170] |
| Calcium | Fibroblast and keratinocyte migration and proliferation; important coagulation factor [130] | Abnormal calcium metabolism; delayed wound healing and chronicity formation [171,172] |
| Zinc | Angiogenesis, tissue re-epithelization, immune and inflammation response [173] | Serum deficiency; delayed wound healing, reduced mitotic activity and scar maturation [174,175] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajj, J.; Sizemore, B.; Singh, K. Impact of Epigenetics, Diet, and Nutrition-Related Pathologies on Wound Healing. Int. J. Mol. Sci. 2024, 25, 10474. https://doi.org/10.3390/ijms251910474
Hajj J, Sizemore B, Singh K. Impact of Epigenetics, Diet, and Nutrition-Related Pathologies on Wound Healing. International Journal of Molecular Sciences. 2024; 25(19):10474. https://doi.org/10.3390/ijms251910474
Chicago/Turabian StyleHajj, John, Brandon Sizemore, and Kanhaiya Singh. 2024. "Impact of Epigenetics, Diet, and Nutrition-Related Pathologies on Wound Healing" International Journal of Molecular Sciences 25, no. 19: 10474. https://doi.org/10.3390/ijms251910474
APA StyleHajj, J., Sizemore, B., & Singh, K. (2024). Impact of Epigenetics, Diet, and Nutrition-Related Pathologies on Wound Healing. International Journal of Molecular Sciences, 25(19), 10474. https://doi.org/10.3390/ijms251910474