Abstract
Maternal obesity has been shown to impair the oxidative status in the placenta and newborns, potentially leading to adverse pregnancy outcomes and long-term effects on the programming of offspring metabolic status. This study aimed to investigate the impact of maternal obesity on maternal and umbilical cord plasma oxidative status, as well as placental oxidative adaptation. Maternal obesity (n = 20), defined as a pre-pregnancy BMI ≥ 25 kg/m2, and maternal leanness (n = 20), defined as a pre-pregnancy BMI < 23 kg/m2, were the group categories used in this study. Both groups were matched according to gestational age at delivery. Maternal blood, umbilical cord blood, and placental tissue were collected to assess nutritional content (cholesterol, triglyceride, and protein), oxidative stress markers (MDA and protein carbonyl), and antioxidant activity (SOD and catalase). Placental protein expression (SOD2, catalase, UCP2, and Nrf2) was evaluated using Western blot analysis. Catalase activity in maternal plasma significantly increased in the maternal obesity group (p = 0.0200), with a trend toward increased MDA and protein carbonyl levels. In umbilical cord plasma, triglyceride, protein carbonyl, and catalase activity were significantly elevated in the maternal obesity group compared with the lean controls (p = 0.0482, 0.0291, and 0.0347, respectively). Placental protein expression analysis revealed significantly decreased SOD2 (p = 0.0011) and catalase (p < 0.0001), along with Nrf2 downregulation (p < 0.0001). An increase in mitochondrial antioxidant UCP2 expression was observed (p = 0.0117). The neonatal protein carbonyl levels positively correlated with placental protein carbonyl (r = 0.7405, p < 0.0001) and negatively correlated with maternal catalase activity (r = −0.4332, p = 0.0052). This study thus provides evidence that maternal obesity is associated with placental and fetal oxidative stress, alongside a concurrent increase in placental antioxidant UCP2 expression.
1. Introduction
Obesity is one of the most significant global public health challenges of the 21st century, with a rising prevalence among women of reproductive age [1]. The criteria for obesity in Asians define the condition as excessive fat accumulation, with a body mass index (BMI) ≥ 25 kg/m2 [2,3]. This condition is particularly concerning in women of reproductive age due to its impact on pregnancy, affecting the health of at least two generations [4]. Maternal obesity represents a stress condition that can lead to impaired feto-placental development and function, potentially exerting long-term negative effects on metabolic programming in the offspring [5,6,7].
During pregnancy, metabolic changes occur in women to ensure sufficient nutrient availability for the developing placenta and fetus [8]. Maternal obesity has been linked to metabolic dysregulation, which disrupts both maternal and fetal metabolic and oxidative status [9,10]. The placenta, as the key organ facilitating maternal–fetal interaction, regulates critical aspects of the intrauterine environment, including oxygen and nutrient delivery, waste removal, and neutralization of reactive oxygen species (ROS) [11]. Thus, oxidative adaptation may modulate the effects of maternal obesity on fetal oxidative stress [12,13].
Pregnancy is considered a state of maternal oxidative stress that can affect the placenta. However, this oxidative stress is exacerbated in obese pregnant women [11,12,13]. Our previous study on placental adaptation to maternal malnutrition in a rat model demonstrated an oxidative imbalance in the placenta [14]. Various enzymes contribute to placental antioxidant defenses, including superoxide dismutase (SOD), catalase, and glutathione peroxidase (GPx) [15]. Additionally, due to the mitochondria-rich nature of the placental tissue, mitochondrial antioxidant uncoupling protein 2 (UCP2) has been proposed as a regulator of oxidative stress defense in the placenta [15,16]. Notably, UCP2 expression has been found to be responsive to changes in maternal nutrition [17]. A recent study reported a negative correlation between maternal BMI and placental SOD expression [18]. Furthermore, maternal oxidative stress markers have been positively correlated with neonatal oxidative stress markers [19]. However, findings on the placental antioxidant response in obese pregnancies are inconsistent. Most studies have been conducted in non-Asian populations using different BMI cut-off points [12,20]. Some studies have reported increased placental antioxidant levels, such as SOD, catalase, and GSH, alongside reduced maternal plasma antioxidant activity [12,20]. In contrast, another study found reduced total antioxidant capacity in the placenta, along with decreased SOD and catalase activity and increased oxidative and nitrosative stress markers [21]. Taken together, these findings suggest that maternal obesity may contribute to a placental oxidative imbalance, potentially predisposing the fetus to oxidative stress.
The aim of this study was to investigate the effect of maternal obesity during pregnancy on maternal and umbilical cord plasma oxidative status, as well as placental oxidative adaptation. We hypothesized that compromised placental antioxidant defenses may contribute to neonatal oxidative stress under adverse maternal conditions.
2. Results
Demographic and pregnancy-outcome data are presented in Table 1. There were no significant differences in maternal age or gestational age at delivery between the maternal lean and the maternal obesity groups (p > 0.05). However, pre-pregnancy weight, pre-pregnancy BMI, and weight at delivery were significantly higher in the maternal obesity group compared to the normal-weight group (p < 0.0001). The percentage of male neonates did not differ significantly between obese and normal-weight women (p > 0.05). Neonatal birth weight and length also did not differ significantly between the two groups (p > 0.05). Interestingly, the placental weight of the maternal obesity group was significantly higher compared to that of the maternal lean group (p = 0.0224), and the feto-placental ratio was significantly lower in the maternal obesity group (p = 0.0374).

Table 1.
Demographics and pregnancy outcomes.
To assess the impact of maternal obesity on the nutritional content on both the maternal and the neonatal sides, we assessed total cholesterol, triglyceride, and protein content in maternal and umbilical cord plasma. There were no significant differences in the maternal plasma levels of cholesterol, triglyceride, or protein between the maternal obesity group and the maternal lean control group (cholesterol: p = 0.3081; triglyceride: p = 0.8163; protein: p = 0.4884; Figure 1a,c,e, respectively). However, a significantly higher level of umbilical cord plasma triglyceride was observed in the maternal obesity group (p = 0.0482; Figure 1d) compared with the lean group, while no significant differences were found in umbilical cord plasma cholesterol or protein content (p = 0.9776 and p = 0.3900, respectively; Figure 1b,f).
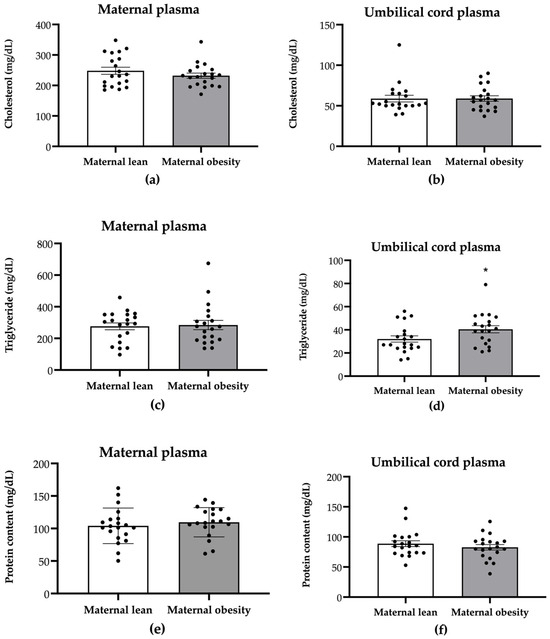
Figure 1.
Maternal plasma and umbilical cord plasma levels of total cholesterol (a,b), triglyceride (c,d), and protein (e,f) in maternal lean and maternal obesity groups. Results are expressed as mean ± SEM, n = 20 each. p-value was tested using Student’s t-test; * p < 0.05 relative to the maternal lean group.
To evaluate whether maternal plasma oxidative stress markers were affected by maternal obesity, we examined the levels of MDA and protein carbonyl. There were no significant differences in the maternal plasma MDA or protein carbonyl levels between the maternal obesity group and the maternal lean control group (p = 0.1452 and p = 0.1744, respectively; Figure 2a,b).
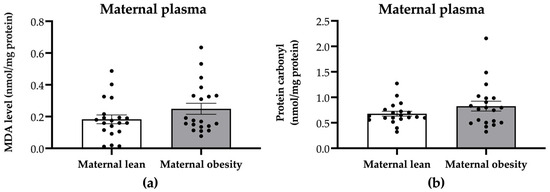
Figure 2.
Levels of (a) malondialdehyde (MDA) and (b) protein carbonyl in the maternal plasma of the maternal lean and maternal obesity groups. Results are expressed as mean ± SEM; n = 20 each. p-value was tested using Student’s t-test.
Maternal plasma catalase activity was significantly decreased in the maternal obesity group compared to the maternal lean group (p = 0.0201; Figure 3a), while SOD activity showed no significant difference between the groups (p = 0.9526; Figure 3b).
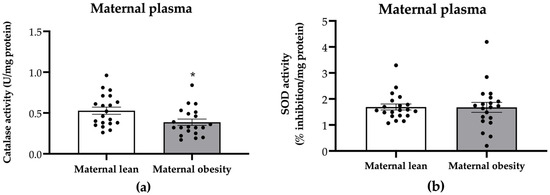
Figure 3.
Activity of (a) catalase and (b) superoxide dismutase (SOD) in the maternal plasma of the maternal lean and maternal obesity groups. Results are expressed as mean ± SEM; n = 20 each; p-value was tested using Student’s t-test; * p < 0.05 relative to the maternal lean group.
The oxidative stress markers in umbilical cord plasma are presented in Figure 4. The protein carbonyl levels were significantly higher in the umbilical cord plasma of the maternal obesity group compared to the maternal lean group (p = 0.0291; Figure 4b). However, there were no significant differences in umbilical cord plasma MDA levels between the maternal obesity group and the maternal lean control (p = 0.9162; Figure 4a).
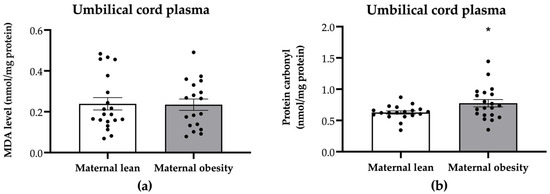
Figure 4.
Levels of (a) malondialdehyde (MDA) and (b) protein carbonyl in the umbilical cord plasma of the maternal lean and maternal obesity groups. Results are expressed as mean ± SEM; n = 20 each. p-value was tested using Student’s t-test. * p < 0.05 relative to the maternal lean group.
Regarding the antioxidant activity on the neonatal side, we assessed catalase and SOD activity in umbilical cord plasma. Notably, the umbilical cord plasma from the maternal obesity group exhibited significantly higher catalase activity (p = 0.0347; Figure 5a), while SOD activity showed no significant difference compared to that in the maternal lean group (p = 0.1739; Figure 5b).
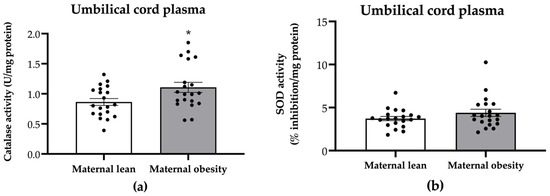
Figure 5.
Activity of (a) catalase and (b) superoxide dismutase (SOD) in the umbilical cord plasma of the maternal lean and maternal obesity groups. Results are expressed as mean ± SEM; n = 20 each. p-value was tested using Student’s t-test; * p < 0.05 relative to the maternal lean group.
Regarding oxidative markers in the placental tissue, the MDA level was not significantly different between the maternal obesity group and the maternal lean control group (p = 0.3415; Figure 6a). However, the placental protein carbonyl level was significantly higher in the maternal obesity group compared to the maternal lean group (p = 0.0300; Figure 6b).
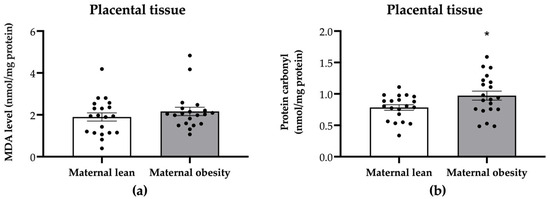
Figure 6.
Level of (a) malondialdehyde (MDA) and (b) protein carbonyl in the placental tissue of the maternal lean and maternal obesity groups. Results are expressed as mean ± SEM; n = 20 each; p-value was tested using Student’s t-test. * p < 0.05 relative to the maternal lean group.
The relative placental antioxidant protein expressions are presented in Figure 7a–d. Catalase and SOD2 expressions in the placental tissue were significantly lower in the maternal obesity group compared to the maternal lean control (p < 0.0001 and p = 0.0011, respectively; Figure 7a,b). Additionally, the expression of the signaling pathway Nrf2 protein was significantly reduced in the maternal obesity group (p < 0.0001; Figure 7c). In contrast, the level of the mitochondrial antioxidant UCP2 was significantly higher in the maternal obesity group compared to the maternal lean group (p = 0.0104; Figure 7d).
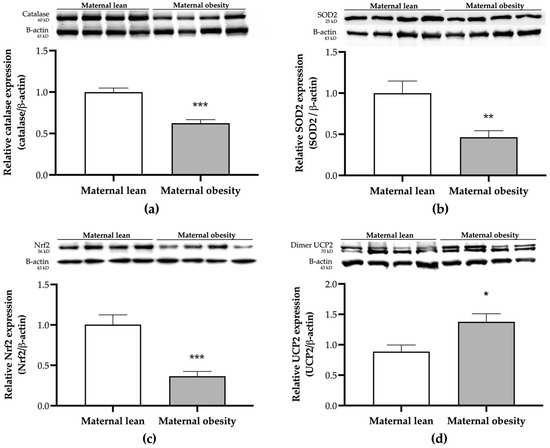
Figure 7.
Placental protein expression of (a) catalase, (b) SOD2, (c) Nrf2, and (d) UCP2, measured using Western blot analysis in the maternal lean and maternal obesity groups (n = 12/group). Results are expressed as mean ± SEM; n = 12 each; p-value was tested using the Man–Whitney test; * p < 0.05, ** p < 0.01, *** p < 0.001 relative to the maternal lean group.
Next, we explored the correlations between newborn oxidative levels and placental and maternal oxidative status. The level of umbilical cord protein carbonyl was positively correlated with that of placental protein carbonyl (r = 0.7405, p < 0.0001) and negatively correlated with maternal catalase activity (r = −0.4332, p = 0.0052). A trend of negative correlation was also observed between umbilical cord protein carbonyl and placental catalase expression (r = −0.3942; p = 0.0566). Additionally, umbilical cord protein carbonyl expression showed a positive correlation with maternal pre-pregnancy weight, pre-pregnancy BMI, and weight at delivery (r = 0.3621, 0.3832, and 0.3168, respectively; p = 0.0217, 0.0147, and 0.0464, respectively) (Table 2).
Table 2.
Correlation between umbilical cord protein carbonyl level and placental oxidative status, as well as with maternal oxidative status and anthropometric parameters.
3. Discussion
In this study, maternal obesity maintained maternal plasma oxidative balance, as suggested by the trend of increased MDA and protein carbonyl levels alongside decreased plasma catalase activity. Interestingly, placental oxidative adaptation was observed, characterized by reduced placental catalase and SOD2 expression, coupled with enhanced UCP2 expression. This placental response was linked to the downregulation of the Nrf2 signaling pathway. Additionally, umbilical cord protein carbonyl and antioxidant catalase activity were significantly increased, indicating oxidative imbalance in the newborns. Neonatal oxidative stress positively correlated with placental oxidative stress and negatively correlated with maternal catalase activity.
Obesity during pregnancy has been associated with dysregulated lipid metabolism, which may expose the fetus to an adverse intrauterine environment [22,23,24]. This condition increases the risk of obstetric complications and adverse neonatal health outcomes at birth, while also affecting fetal metabolic programming and potentially influencing long-term health [25]. Maternal obesity has been linked to alterations in plasma lipid, glucose, and protein levels [26,27]. In our study, maternal and umbilical cord cholesterol and triglyceride levels in both groups fell within the normal ranges for the third trimester of pregnancy, indicating adequate nutritional health in both mothers and their newborns [28,29]. We did not observe any significant differences in total cholesterol, triglyceride, or protein levels in the maternal plasma of the obesity group. This contrasts with previous studies reporting elevated cholesterol and triglyceride levels in obese pregnancies. A possible explanation for this discrepancy could be differences in the study populations, which may result in varying genetic predispositions to dyslipidemia, as well as differences in BMI that might influence the severity of obesity [24,27,30]. However, the umbilical cord plasma from the maternal obesity group had higher triglyceride levels compared to the that from the lean control. Similar findings have been reported, showing elevated triglyceride levels in fetuses and placentas of mothers with metabolic disorders [11,20,31]. Additionally, there was evidence of dysregulation in placental fatty acid transporters and lipase enzymes in mothers with complications such as metabolic disorders, which directly affects lipid transfer from the placenta to the fetus [31,32,33]. We hypothesize that maternal obesity may lead to an increased transport of free fatty acids to the fetus, resulting in greater fat accumulation in the form of triglycerides. Elevated triglyceride levels can contribute to inflammation and potentially impact organ development and function [34,35]. Consequently, fetuses with elevated triglyceride levels are more likely to become obese in childhood and adulthood [33,36]. Therefore, our findings suggest that, even when the maternal nutrient levels are within normal ranges, maternal obesity can impair the fetal lipid metabolism, potentially increasing the risk of metabolic disorders later in life.
Additionally, maternal overnutrition has been correlated with an altered oxidative status in both the placenta and the newborn [7,35,37]. In normal pregnancy, increased ROS production is typically accompanied by an upregulation of antioxidant enzymes [38]. However, the oxidative status in maternal obesity remains controversial [19,21,39]. To investigate this, we assessed the oxidative status in maternal plasma by measuring lipoperoxidation products (MDA and protein carbonyl) alongside catalase and SOD activity. Our results showed no significant changes in the plasma MDA and protein carbonyl levels in the maternal obesity group, but there was a reduction in catalase activity. Catalase is a crucial antioxidant responsible for reducing free radicals and converting the final products of lipid peroxidation into inactive forms [40]. The reduced catalase activity observed in maternal obesity could stem from an impaired antioxidant response or from the depletion of antioxidants due to prolonged ROS exposure [20]. This impairment may be linked to diminished Nrf2 signaling or to the overactivation of xanthine oxidase, both of which have been associated with maternal metabolic disorders [41,42]. Our findings align with previous studies, which have shown that pregnant women with obesity exhibit elevated levels of plasma MDA and other oxidative stress markers, along with alterations in plasma antioxidants [19,20]. Hence, our data suggest that maternal obesity, even in the absence of obstetric complications, is associated with oxidative imbalance.
There is evidence that maternal nutrition influences the oxidative status in the placenta [18]. Maintaining a proper balance between pro-oxidants and antioxidants is essential for preserving placental homeostasis as part of its adaptive response [12]. Under physiological conditions, reactive molecules play a role in cell signaling, regulating normal placental development and functions such as angiogenesis, nutrient transport, hormone production, and oxidative control. However, excessive radical production can disrupt placental function [43]. To assess the impact of maternal obesity on placental oxidative alterations, we evaluated the oxidative status in the placenta. As expected, maternal obesity was associated with an altered placental oxidative status, demonstrated by a significant increase in placental protein carbonyl levels and a compromised antioxidant enzyme expression. Specifically, we observed decreased SOD2 and catalase levels, as well as reduced expression of the downstream signaling pathway Nrf2. In contrast, mitochondrial antioxidant UCP2 expression was elevated. SOD2 and catalase are general antioxidants located in mitochondria and peroxisomes, respectively [20], and their expression is regulated by Nrf2, a key transcriptional regulator of cellular responses to oxidative stress [44]. The reduction in placental SOD2 and catalase suggests the inactivation of the antioxidant system in the placenta of obese mothers. Furthermore, the significant reduction in Nrf2 protein expression observed in this study provides a clear mechanistic explanation for the impaired placental antioxidant defense. Several studies support our findings, showing elevated levels of oxidative stress markers along with downregulated SOD, catalase, and GPx in the placentas of mothers with metabolic disorders [12,18,20]. UCP2 is also recognized as a key mitochondrial antioxidant in the placenta [14]. Given the high mitochondrial activity in the placenta and the observed reductions in SOD and catalase expression, the upregulation of UCP2 may represent a compensatory response to the pro-oxidant state induced by maternal metabolic abnormalities. Our findings highlight the role of UCP2 in maintaining oxidative balance in the placenta, as previously reported [13,14]. However, despite the upregulation of UCP2, which may reflect placental adaptation to increased ROS production, there was a rise in placental protein carbonyl levels, indicating oxidative stress. While the mode of delivery has been linked to changes in placental oxidative stress [45], there was no significant difference in delivery mode between the two groups, ruling out its influence on the elevated placental protein carbonyl levels observed in maternal obesity. This suggests that the antioxidant response in the placentas of obese mothers may be insufficient to restore oxidative balance, potentially leading to placental dysfunction. Further studies on placental enzymatic activity could provide additional insights into this mechanism.
In addition to maternal and placental changes, we also observed alterations in the oxidative status of the newborn. We hypothesized that placental dysfunction induced by maternal obesity might impair the placenta’s ability to protect the fetus from adverse oxidative effects. Our results demonstrated significantly increased protein carbonyl levels and a non-significant trend toward higher MDA levels in the umbilical cord plasma from infants born to obese mothers compared to that from infants born to normal-weight mothers. Protein carbonylation, an irreversible oxidative modification, is a general marker of oxidative stress [46]. Elevated lipid peroxidation in the umbilical cord plasma further supports the presence of oxidative stress, as lipids are primary targets of radical reactions. The increased fatty acid transfer to the fetus through the placenta in maternal obesity may contribute to this oxidative stress [35]. Our findings are consistent with other studies reporting higher levels of oxidative stress markers in newborns of obese mothers [37,47]. Regarding the neonatal antioxidant response, our study found significantly increased umbilical cord plasma catalase activity without changes in SOD activity. Previous research showed that catalase plays a critical role in responding to oxidative stress under high H₂O₂ conditions, whereas SOD remains unaffected [48,49], which may explain the lack of alteration in SOD activity. The observed increase in catalase activity in newborns exposed to maternal obesity suggests an active antioxidant defense system, likely stimulated to neutralize the harmful effects of ROS. The simultaneous increase in protein oxidative products and antioxidant enzyme activity indicates that maternal obesity compromises neonatal oxidative status. To determine the connection between oxidative stress in newborns and placental oxidative status, we assessed the correlation between umbilical cord protein carbonyl and placental oxidative markers. The umbilical cord protein carbonyl levels were positively correlated with the placental protein carbonyl levels, while negatively correlated with placental catalase expression and maternal catalase activity. These findings suggest that neonatal oxidative stress is linked to an impaired placental oxidative function. Additionally, our analysis showed a positive correlation between umbilical cord protein carbonyl levels and maternal pre-pregnancy weight, BMI, and weight at delivery, indicating that maternal obesity likely contributes to oxidative stress in both the placenta and the newborn. This may play a role in fetal programming, increasing susceptibility to oxidative stress-related diseases later in life [35,50].
Regarding neonatal outcomes, no significant differences were observed in neonatal birth weight or length. However, the growth of the infant was disproportionate to the growth of the placenta, as indicated by a low feto-placental ratio and a high placental weight. Increased placental weight has been associated with greater capillary surface density in cotyledons, enhancing the available surface area for placental exchange and transport functions [51,52]. This enlargement of the placenta demands more nutrients, potentially resulting in inadequate nutrient supply for the developing fetus [53,54]. These findings suggest impaired placental efficiency in the context of metabolic oxidative imbalance, as previously reported [18,54].
However, this study has several limitations. First, it focused exclusively on lean and obese groups; future research should include individuals with overweight pregnancies for a more comprehensive analysis. Second, the study did not perform immunohistochemical assessments of placental vascular density or examine the localization of oxidative stress-related substances within the placenta. Further research in these areas is needed to validate our findings and clarify the underlying mechanisms. Despite these limitations, our study provides evidence that maternal obesity is associated with placental and fetal oxidative stress, along with a concurrent increase in placental antioxidant expression. The neonatal oxidative stress observed in obese pregnancies may contribute to fetal programming, increasing susceptibility to oxidative stress-related diseases later in life. A follow-up study examining the metabolic and oxidative status of children born to mothers with obesity is warranted.
4. Materials and Methods
4.1. Study Subjects
This study was approved by the Khon Kaen University Ethics Committee for Human Research in adherence with the Declaration of Helsinki and the ICH Good Clinical Practice Guidelines. The study identification code is HE651023. All participants were informed and provided written consent to participate as study volunteers.
A total of 40 pregnant women attending the labor department for delivery at Srinagarind Hospital, Faculty of Medicine, Khon Kaen University, Khon Kaen, Thailand, were recruited for this cross-sectional study. The study groups were classified based on the recommended BMI cut-off points for determining overweight and obesity in Asian populations [2,3] into maternal lean (pre-pregnancy BMI < 23 kg/m2) (n = 20) and maternal obesity (pre-pregnancy BMI ≥ 25 kg/m2) (n = 20) groups. Both groups were matched for gestational age at delivery. All volunteers had no history of chronic diseases, including chronic hypertension, diabetes mellitus, and kidney, liver, immune, or respiratory diseases.
4.2. Characteristics of the Mothers and Their Offspring
Anthropometric data and pregnancy outcomes, including maternal age, gestational age at delivery, pre-pregnancy weight, pre-pregnancy BMI, weight at delivery, neonatal gender, neonatal birth weight and length, and placental weight, were collected through personal interviews and medical records.
4.3. Maternal Plasma Collection
A maternal venous blood sample (10 mL) was collected from the arm veins of the mothers before delivery using ethylenediaminetetraacetic acid (EDTA) tubes. The blood was centrifuged at 3500 rpm for 15 min, and the plasma was separated and stored at −80 °C until analysis. The plasma was used to measure nutritional content (total cholesterol, triglyceride, and protein), oxidative stress markers (MDA and protein carbonyl), and antioxidant activity (SOD and catalase).
4.4. Umbilical Cord Plasma Collection
Immediately after the birth of the newborn and the separation of the umbilical cord, the cord was clamped, with another clamp placed 8–10 inches away from the first. A blood sample (10 mL) was promptly collected from the section between the clamps into a tube containing EDTA. The blood was centrifuged at 3500 rpm for 15 min, and the plasma was separated and stored at −80 °C until analysis. The umbilical cord plasma was used to measure nutritional content (total cholesterol, triglyceride, and protein), oxidative stress markers (MDA and protein carbonyl), and antioxidant activity (SOD and catalase).
4.5. Placental Sample Collection
Placental samples were randomly harvested within 10 min of delivery. Small fragments of villous tissue (~100 mg) were isolated from the central area of the placenta, placed on ice, and washed with phosphate-buffered solution (PBS) to remove any remaining blood. The samples were then stored at −80 °C until further analysis.
4.6. Assessment of Plasma and Placental MDA
The levels of MDA in maternal plasma, umbilical cord plasma, and placental tissue were assessed following a previously described protocol [55]. In brief, 150 μL of plasma was mixed with a stock reagent containing 10% TCA, 5 mmol/L EDTA, 8% SDS, and 0.5 μg/mL BHT. The mixture was incubated at room temperature for 10 min, followed by the addition of 0.6% TBA. The mixture was then boiled in a water bath for 30 min. After cooling, it was centrifuged at 1000× g for 10 min. The absorbance of the supernatant was measured at 532 nm using a spectrophotometer. A standard curve was generated using 1,1,3,3-tetraethoxypropane, and the data are expressed as nmol/mg protein.
4.7. Assessment of Plasma and Placental Protein Carbonyl
The plasma protein carbonyl content was determined using 2,4-dinitrophenylhydrazine (DNPH) derivatization, followed by spectrophotometric analysis at 370 nm, as previously described [49]. The extinction coefficient of DNPH was used to convert the absorbance into protein carbonyl concentration, expressed as nmol/mg protein.
4.8. Assessment of Plasma Catalase Activity
Catalase activity in maternal plasma, umbilical cord plasma, and placental tissue was assessed following a previously described protocol [55]. In brief, 20 µL of plasma or standard solution was added to a microplate, followed by a substrate mixture containing 65 μmol/mL of H₂O₂ in 60 mmol/L PBS (pH 7.4). After incubating at 37 °C for 1 min, 100 µL of ammonium molybdate ((NH4)6Mo7O24·4H2O) was added to stop the reaction. The yellow complex formed was measured at a wavelength of 405 nm. Catalase from bovine liver (Sigma-Aldrich, Ann Arbor, MI, USA) was used to generate a standard curve, and the data are expressed as U/mg protein.
4.9. Assessment of Plasma SOD Activity
Plasma SOD activity was assessed using the SOD Assay Kit-WST (Dojindo Laboratories, Japan) based on colorimetric analysis. The procedure involved adding 20 µL of sample solution to each well, with 20 µL of ultrapure H2O added to each blank well. The working solution, dilution buffer, and enzyme solution were then added, followed by incubation at 37 °C for 20 min. The absorbance was measured at 450 nm using a microplate reader. SOD activity was calculated as the % inhibition rate per mg of protein.
4.10. Assessment of Plasma Protein Content and Lipids
Maternal blood and umbilical cord blood were used to measure the lipid profiles, including total cholesterol and triglycerides. These measurements were performed at the Community Medical Laboratory, Faculty of Associated Medical Sciences, Khon Kaen University.
Protein concentration in the plasma samples was quantified using the Bradford assay. Briefly, diluted samples were incubated with the Bradford reagent, and the absorbance was measured at 595 nm using a microplate reader. A standard curve, generated using a bovine serum albumin (BSA) protein standard, was used to convert the absorbance values into protein concentrations.
4.11. Placental Protein Extraction and Expression Analysis
The placental samples were homogenized in PBS containing proteolytic enzyme inhibitors (Complete-Mini; Roche, Basel, Switzerland) using an Ultra-Turrax homogenizer (Bioblock Scientific, Illkirch, France). After homogenization, the samples were centrifuged at 4000 rpm for 30 min, and the supernatant (protein extracts) was collected for Western blot analysis.
Placental antioxidant protein expression was measured using 30–50 µg of protein per sample. The proteins were separated on 10–12% SDS-PAGE gels. Primary antibodies against SOD2 (Santa Cruz Biotechnology, Heidelberg, Germany; 1:1000), catalase (Sigma-Aldrich, St. Louis, MO, USA; 1:2000), UCP2 (Santa Cruz Biotechnology, Heidelberg, Germany; 1:1000), and Nrf2 (Santa Cruz Biotechnology, Heidelberg, Germany; 1:500) were applied overnight at 4 °C. After washing, a secondary antibody (anti-mouse IgG peroxidase-conjugated) was applied for 1 h at room temperature. The blots were washed again and incubated with enhanced chemiluminescence reagents (ECL Prime, Amersham Bioscience, Little Chalfont, Buckinghamshire, UK), and the bands were detected using the ChemiDoc XRS+ imaging system (Bio-Rad, Ann Arbor, MI, USA). The blots were re-incubated with a β-actin antibody (Santa Cruz Biotechnology, Heidelberg, Germany; 1:4000) as an internal control. The expression levels of SOD2, catalase, UCP2, and Nrf2 were normalized with β-actin.
4.12. Statistical Analysis
The data were statistically analyzed using GraphPad Prism software version 9.3.1 (GraphPad, East Lansing, MI, USA). The Kolmogorov–Smirnov test was used to assess data normality. Normally distributed data are expressed as mean ± standard error of the mean (SEM). Comparisons between the two groups were performed using Student’s t-test and Pearson’s Chi-square test. Correlations between parameters were evaluated using Pearson’s correlation test. Differences were considered statistically significant when p < 0.05.
5. Conclusions
This study provides evidence that maternal obesity disrupts the maternal–fetal–placental oxidative balance, with neonatal oxidative stress linked to impaired placental oxidative function. The oxidative stress observed in neonates from obese pregnancies may be a key factor in fetal programming, increasing the susceptibility to oxidative stress-related diseases later in life. Understanding the relationship between neonatal and placental oxidative stress could offer valuable insights into oxidative damage caused by maternal obesity and help inform strategies to mitigate this damage.
Author Contributions
Conceptualization, T.J., S.P., K.K., S.M.A. and W.S.-I.; methodology, S.P., T.J., K.K., S.M.A. and W.S.-I.; validation, S.P., S.M.A., P.R.-R. and W.S.-I.; formal analysis, T.J., S.P., S.M.A. and W.S.-I.; investigation, S.P., K.K., T.J., M.I., P.R.-R. and W.S.-I.; writing—original draft preparation, S.P., T.J., K.K., S.M.A., P.R.-R. and W.S.-I.; writing—review and editing, S.P., T.J., S.M.A., P.R.-R. and W.S.-I. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Young Researcher Development Project of Khon Kaen University Year 2022, Thailand, and by the Research Institute for Human High Performance and Health Promotion (HHP&HP), Khon Kaen University, Thailand, and Invitation Research Grant (No. IN67029), Faculty of Medicine, Khon Kaen University, Thailand.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and the ICH Good Clinical Practice Guidelines and approved by the Khon Kaen University Ethics Committee for Human Research, Khon Kaen University, Khon Kaen, Thailand; study identification code: HE651023 (date of approval: 22 March 2022).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data are contained within the article. Additional raw data can be obtained from the corresponding author on request.
Acknowledgments
We thank the women who participated in the study; the staff of the labor department at Srinagarind Hospital, Faculty of Medicine, Khon Kaen University, Khon Kaen, Thailand, for their support in collecting samples for this study; and the Department of Physiology, Faculty of Medicine, Universidad Autónoma de Madrid, Madrid, Spain, for providing the reagents for Western blot analysis. We would also like to acknowledge Dylan Southard, for editing the MS via the KKU Publication Clinic (Thailand).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tauqeer, Z.; Gomez, G.; Stanford, F.C. Obesity in Women: Insights for the Clinician. J. Women’s Health 2018, 27, 444–457. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Appropriate body-mass index for Asian populations and its implications for policy and intervention strategies. Lancet 2004, 363, 157–163. [Google Scholar] [CrossRef]
- World Health Organization. The Asia-Pacific perspective: Redefining obesity and its treatment. Health Commun. Aust. 2000. Available online: https://apps.who.int/iris/handle/10665/206936 (accessed on 6 October 2024).
- Lim, S.; Harrison, C.; Callander, E.; Walker, R.; Teede, H.; Moran, L. Addressing Obesity in Preconception, Pregnancy, and Postpartum: A Review of the Literature. Curr. Obes. Rep. 2022, 11, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.C.; Powell, T.L.; Jansson, T. Placental function in maternal obesity. Clin. Sci. 2020, 134, 961–984. [Google Scholar] [CrossRef]
- Lamothe, J.; Khurana, S.; Tharmalingam, S.; Williamson, C.; Byrne, C.J.; Lees, S.J.; Khaper, N.; Kumar, A.; Tai, T.C. Oxidative Stress Mediates the Fetal Programming of Hypertension by Glucocorticoids. Antioxidants 2021, 10, 531. [Google Scholar] [CrossRef]
- Palaska, E.; Antoniou, E.; Tzitiridou-Chatzopoulou, M.; Eskitzis, P.; Orovou, E. Correlation between Breastfeeding, Maternal Body Mass Index, and Childhood Obesity. Epidemiologia 2024, 5, 411–420. [Google Scholar] [CrossRef]
- Jovandaric, M.Z.; Babic, S.; Raus, M.; Medjo, B. The Importance of Metabolic and Environmental Factors in the Occurrence of Oxidative Stress during Pregnancy. Int. J. Mol. Sci. 2023, 24, 11964. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Y.; Mao, X.; Du, M. Maternal obesity impairs fetal mitochondriogenesis and brown adipose tissue development partially via upregulation of miR-204-5p. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 2706–2715. [Google Scholar] [CrossRef]
- Anam, A.K.; Cooke, K.M.; Dratver, M.B.; O’Bryan, J.V.; Perley, L.E.; Guller, S.M.; Hwang, J.J.; Taylor, H.S.; Goedeke, L.; Kliman, H.J.; et al. Insulin increases placental triglyceride as a potential mechanism for fetal adiposity in maternal obesity. Mol. Metab. 2022, 64, 101574. [Google Scholar] [CrossRef]
- Johns, E.C.; Denison, F.C.; Reynolds, R.M. The impact of maternal obesity in pregnancy on placental glucocorticoid and macronutrient transport and metabolism. Biochim. Biophys. Acta Mol. Basis. Dis. 2020, 1866, 165374. [Google Scholar] [CrossRef] [PubMed]
- Santos-Rosendo, C.; Bugatto, F.; Gonzalez-Dominguez, A.; Lechuga-Sancho, A.M.; Mateos, M.R.; Visiedo, F. Placental Adaptive Changes to Protect Function and Decrease Oxidative Damage in Metabolically Healthy Maternal Obesity. Antioxidants 2020, 9, 794. [Google Scholar] [CrossRef] [PubMed]
- Martino, J.; Sebert, S.; Segura, M.T.; Garcia-Valdes, L.; Florido, J.; Padilla, M.C.; Marcos, A.; Rueda, R.; McArdle, H.J.; Budge, H.; et al. Maternal Body Weight and Gestational Diabetes Differentially Influence Placental and Pregnancy Outcomes. J. Clin. Endocrinol. Metab. 2016, 101, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Phuthong, S.; Reyes-Hernandez, C.G.; Rodriguez-Rodriguez, P.; Ramiro-Cortijo, D.; Gil-Ortega, M.; Gonzalez-Blazquez, R.; Gonzalez, M.C.; Lopez de Pablo, A.L.; Arribas, S.M. Sex Differences in Placental Protein Expression and Efficiency in a Rat Model of Fetal Programming Induced by Maternal Undernutrition. Int. J. Mol. Sci. 2020, 22, 237. [Google Scholar] [CrossRef]
- Jones, M.L.; Mark, P.J.; Lewis, J.L.; Mori, T.A.; Keelan, J.A.; Waddell, B.J. Antioxidant defenses in the rat placenta in late gestation: Increased labyrinthine expression of superoxide dismutases, glutathione peroxidase 3, and uncoupling protein 2. Biol. Reprod. 2010, 83, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Stark, M.J.; Hodyl, N.A.; Butler, M.; Clifton, V.L. Localisation and characterisation of uncoupling protein-2 (UCP2) in the human preterm placenta. Placenta 2012, 33, 1020–1025. [Google Scholar] [CrossRef]
- Gnanalingham, M.G.; Williams, P.; Wilson, V.; Bispham, J.; Hyatt, M.A.; Pellicano, A.; Budge, H.; Stephenson, T.; Symonds, M.E. Nutritional manipulation between early to mid-gestation: Effects on uncoupling protein-2, glucocorticoid sensitivity, IGF-I receptor and cell proliferation but not apoptosis in the ovine placenta. Reproduction 2007, 134, 615–623. [Google Scholar] [CrossRef][Green Version]
- Mando, C.; Castiglioni, S.; Novielli, C.; Anelli, G.M.; Serati, A.; Parisi, F.; Lubrano, C.; Zocchi, M.; Ottria, R.; Giovarelli, M. Placental Bioenergetics and Antioxidant Homeostasis in Maternal Obesity and Gestational Diabetes. Antioxidants 2024, 13, 858. [Google Scholar] [CrossRef]
- Lopez-Yanez Blanco, A.; Diaz-Lopez, K.M.; Vilchis-Gil, J.; Diaz-Garcia, H.; Gomez-Lopez, J.; Medina-Bravo, P.; Granados-Riveron, J.T.; Gallardo, J.M.; Klunder-Klunder, M.; Sanchez-Urbina, R. Diet and Maternal Obesity Are Associated with Increased Oxidative Stress in Newborns: A Cross-Sectional Study. Nutrients 2022, 14, 746. [Google Scholar] [CrossRef]
- Malti, N.; Merzouk, H.; Merzouk, S.A.; Loukidi, B.; Karaouzene, N.; Malti, A.; Narce, M. Oxidative stress and maternal obesity: Feto-placental unit interaction. Placenta 2014, 35, 411–416. [Google Scholar] [CrossRef]
- Evans, L.; Myatt, L. Sexual dimorphism in the effect of maternal obesity on antioxidant defense mechanisms in the human placenta. Placenta 2017, 51, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Zafaranieh, S.; Stadler, J.T.; Pammer, A.; Marsche, G.; van Poppel, M.N.M.; Desoye, G.; Dali Core Investigator, G. The Association of Physical Activity and Sedentary Behavior with Maternal and Cord Blood Anti-Oxidative Capacity and HDL Functionality: Findings of DALI Study. Antioxidants 2023, 12, 827. [Google Scholar] [CrossRef] [PubMed]
- Mannino, A.; Sarapis, K.; Moschonis, G. The Effect of Maternal Overweight and Obesity Pre-Pregnancy and During Childhood in the Development of Obesity in Children and Adolescents: A Systematic Literature Review. Nutrients 2022, 14, 5125. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Chen, Y.; Wu, X.; Yin, A.; Tian, F.; Zhang, H.; Huang, X.; Wu, L.; Niu, J. Mediation effect of maternal triglyceride and fasting glucose level on the relationship between maternal overweight/ obesity and fetal growth: A prospective cohort study. BMC Pregnancy Childbirth 2023, 23, 449. [Google Scholar] [CrossRef] [PubMed]
- Francis, E.C.; Kechris, K.; Johnson, R.K.; Rawal, S.; Pathmasiri, W.; Rushing, B.R.; Du, X.; Jansson, T.; Dabelea, D.; Sumner, S.J.; et al. Maternal Serum Metabolomics in Mid-Pregnancy Identifies Lipid Pathways as a Key Link to Offspring Obesity in Early Childhood. Int. J. Mol. Sci. 2024, 25, 7620. [Google Scholar] [CrossRef]
- Corrales, P.; Vidal-Puig, A.; Medina-Gomez, G. Obesity and pregnancy, the perfect metabolic storm. Eur. J. Clin. Nutr. 2021, 75, 1723–1734. [Google Scholar] [CrossRef]
- Song, X.; Chen, L.; Zhang, S.; Liu, Y.; Wei, J.; Sun, M.; Shu, J.; Wang, T.; Qin, J. High Maternal Triglyceride Levels Mediate the Association between Pre-Pregnancy Overweight/Obesity and Macrosomia among Singleton Term Non-Diabetic Pregnancies: A Prospective Cohort Study in Central China. Nutrients 2022, 14, 2075. [Google Scholar] [CrossRef]
- Abbassi-Ghanavati, M.; Greer, L.G.; Cunningham, F.G. Pregnancy and laboratory studies: A reference table for clinicians. Obstet. Gynecol. 2009, 114, 1326–1331. [Google Scholar] [CrossRef]
- Go, H.; Hashimoto, K.; Maeda, H.; Ogasawara, K.; Kume, Y.; Murata, T.; Sato, A.; Ogata, Y.; Shinoki, K.; Nishigori, H.; et al. Cord blood triglyceride and total cholesterol in preterm and term neonates: Reference values and associated factors from the Japan Environment and Children’s Study. Eur. J. Pediatr. 2023, 182, 4547–4556. [Google Scholar] [CrossRef]
- Stein, R.; Ferrari, F.; Scolari, F. Genetics, Dyslipidemia, and Cardiovascular Disease: New Insights. Curr. Cardiol. Rep. 2019, 21, 68. [Google Scholar] [CrossRef]
- Zhou, J.; Bai, J.; Guo, Y.; Fu, L.; Xing, J. Higher Levels of Triglyceride, Fatty Acid Translocase, and Toll-Like Receptor 4 and Lower Level of HDL-C in Pregnant Women with GDM and Their Close Correlation with Neonatal Weight. Gynecol. Obstet. Investig. 2021, 86, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Keleher, M.R.; Erickson, K.; Kechris, K.; Yang, I.V.; Dabelea, D.; Friedman, J.E.; Boyle, K.E.; Jansson, T. Associations between the activity of placental nutrient-sensing pathways and neonatal and postnatal metabolic health: The ECHO Healthy Start cohort. Int. J. Obes. 2020, 44, 2203–2212. [Google Scholar] [CrossRef] [PubMed]
- Leonard, S.A.; Rasmussen, K.M.; King, J.C.; Abrams, B. Trajectories of maternal weight from before pregnancy through postpartum and associations with childhood obesity. Am. J. Clin. Nutr. 2017, 106, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Balachandiran, M.; Bobby, Z.; Dorairajan, G.; Jacob, S.E.; Gladwin, V.; Vinayagam, V.; Packirisamy, R.M. Placental Accumulation of Triacylglycerols in Gestational Diabetes Mellitus and Its Association with Altered Fetal Growth are Related to the Differential Expressions of Proteins of Lipid Metabolism. Exp. Clin. Endocrinol. Diabetes 2021, 129, 803–812. [Google Scholar] [CrossRef]
- Phengpol, N.; Thongnak, L.; Lungkaphin, A. The programming of kidney injury in offspring affected by maternal overweight and obesity: Role of lipid accumulation, inflammation, oxidative stress, and fibrosis in the kidneys of offspring. J. Physiol. Biochem. 2023, 79, 1–17. [Google Scholar] [CrossRef]
- Wang, L.; O’Kane, A.M.; Zhang, Y.; Ren, J. Maternal obesity and offspring health: Adapting metabolic changes through autophagy and mitophagy. Obes. Rev. 2023, 24, e13567. [Google Scholar] [CrossRef]
- Hernandez-Trejo, M.; Montoya-Estrada, A.; Torres-Ramos, Y.; Espejel-Nunez, A.; Guzman-Grenfell, A.; Morales-Hernandez, R.; Tolentino-Dolores, M.; Laresgoiti-Servitje, E. Oxidative stress biomarkers and their relationship with cytokine concentrations in overweight/obese pregnant women and their neonates. BMC Immunol. 2017, 18, 3. [Google Scholar] [CrossRef]
- Hussain, T.; Murtaza, G.; Metwally, E.; Kalhoro, D.H.; Kalhoro, M.S.; Rahu, B.A.; Sahito, R.G.A.; Yin, Y.; Yang, H.; Chughtai, M.I.; et al. The Role of Oxidative Stress and Antioxidant Balance in Pregnancy. Mediat. Inflamm. 2021, 2021, 9962860. [Google Scholar] [CrossRef]
- Ballesteros-Guzman, A.K.; Carrasco-Legleu, C.E.; Levario-Carrillo, M.; Chavez-Corral, D.V.; Sanchez-Ramirez, B.; Marinelarena-Carrillo, E.O.; Guerrero-Salgado, F.; Reza-Lopez, S.A. Prepregnancy Obesity, Maternal Dietary Intake, and Oxidative Stress Biomarkers in the Fetomaternal Unit. Biomed. Res. Int. 2019, 2019, 5070453. [Google Scholar] [CrossRef]
- Alfonso-Prieto, M.; Biarnes, X.; Vidossich, P.; Rovira, C. The molecular mechanism of the catalase reaction. J. Am. Chem. Soc. 2009, 131, 11751–11761. [Google Scholar] [CrossRef]
- Biri, A.; Onan, A.; Devrim, E.; Babacan, F.; Kavutcu, M.; Durak, I. Oxidant status in maternal and cord plasma and placental tissue in gestational diabetes. Placenta 2006, 27, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Psefteli, P.M.; Morris, J.K.; Ehler, E.; Smith, L.; Bowe, J.; Mann, G.E.; Taylor, P.D.; Chapple, S.J. Sulforaphane induced NRF2 activation in obese pregnancy attenuates developmental redox imbalance and improves early-life cardiovascular function in offspring. Redox Biol. 2023, 67, 102883. [Google Scholar] [CrossRef] [PubMed]
- Ruano, C.S.M.; Miralles, F.; Mehats, C.; Vaiman, D. The Impact of Oxidative Stress of Environmental Origin on the Onset of Placental Diseases. Antioxidants 2022, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Zhai, X.; Qiu, Y.; Lu, X.; Jiao, Y. The Nrf2 in Obesity: A Friend or Foe? Antioxidants 2022, 11, 2067. [Google Scholar] [CrossRef]
- Cindrova-Davies, T.; Yung, H.W.; Johns, J.; Spasic-Boskovic, O.; Korolchuk, S.; Jauniaux, E.; Burton, G.J.; Charnock-Jones, D.S. Oxidative stress, gene expression, and protein changes induced in the human placenta during labor. Am. J. Pathol. 2007, 171, 1168–1179. [Google Scholar] [CrossRef]
- Song, Y.R.; Kim, J.K.; Lee, H.S.; Kim, S.G.; Choi, E.K. Serum levels of protein carbonyl, a marker of oxidative stress, are associated with overhydration, sarcopenia and mortality in hemodialysis patients. BMC Nephrol. 2020, 21, 281. [Google Scholar] [CrossRef]
- Negro, S.; Boutsikou, T.; Briana, D.D.; Tataranno, M.L.; Longini, M.; Proietti, F.; Bazzini, F.; Dani, C.; Malamitsi-Puchner, A.; Buonocore, G.; et al. Maternal obesity and perinatal oxidative stress: The strength of the association. J. Biol. Regul. Homeost. Agents 2017, 31, 221–227. [Google Scholar]
- Rocha, S.; Gomes, D.; Lima, M.; Bronze-da-Rocha, E.; Santos-Silva, A. Peroxiredoxin 2, glutathione peroxidase, and catalase in the cytosol and membrane of erythrocytes under H2O2-induced oxidative stress. Free Radic. Res. 2015, 49, 990–1003. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef]
- Arogbokun, O.; Rosen, E.; Keil, A.P.; Milne, G.L.; Barrett, E.; Nguyen, R.; Bush, N.R.; Swan, S.H.; Sathyanarayana, S.; Ferguson, K.K. Maternal Oxidative Stress Biomarkers in Pregnancy and Child Growth from Birth to Age 6. J. Clin. Endocrinol. Metab. 2021, 106, 1427–1436. [Google Scholar] [CrossRef]
- Redmer, D.A.; Milne, J.S.; Aitken, R.P.; Johnson, M.L.; Borowicz, P.P.; Reynolds, L.P.; Caton, J.S.; Wallace, J.M. Decreasing maternal nutrient intake during the final third of pregnancy in previously overnourished adolescent sheep: Effects on maternal nutrient partitioning and feto-placental development. Placenta 2012, 33, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Louwen, F.; Kreis, N.N.; Ritter, A.; Yuan, J. Maternal obesity and placental function: Impaired maternal-fetal axis. Arch. Gynecol. Obstet. 2024, 309, 2279–2288. [Google Scholar] [CrossRef] [PubMed]
- Correia-Branco, A.; Keating, E.; Martel, F. Maternal undernutrition and fetal developmental programming of obesity: The glucocorticoid connection. Reprod. Sci. 2015, 22, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.D.; Hernandez, M.H.; Serazin, V.; Vialard, F.; Dieudonne, M.N. Human Placental Adaptive Changes in Response to Maternal Obesity: Sex Specificities. Int. J. Mol. Sci. 2023, 24, 9770. [Google Scholar] [CrossRef]
- Poasakate, A.; Maneesai, P.; Rattanakanokchai, S.; Bunbupha, S.; Tong-Un, T.; Pakdeechote, P. Genistein Prevents Nitric Oxide Deficiency-Induced Cardiac Dysfunction and Remodeling in Rats. Antioxidants 2021, 10, 237. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).