
A Versatile Aldehyde: Ferredoxin Oxidoreductase from the Organic Acid Reducing Thermoanaerobacter sp. Strain X514


Abstract
:1. Introduction
2. Results and Discussion
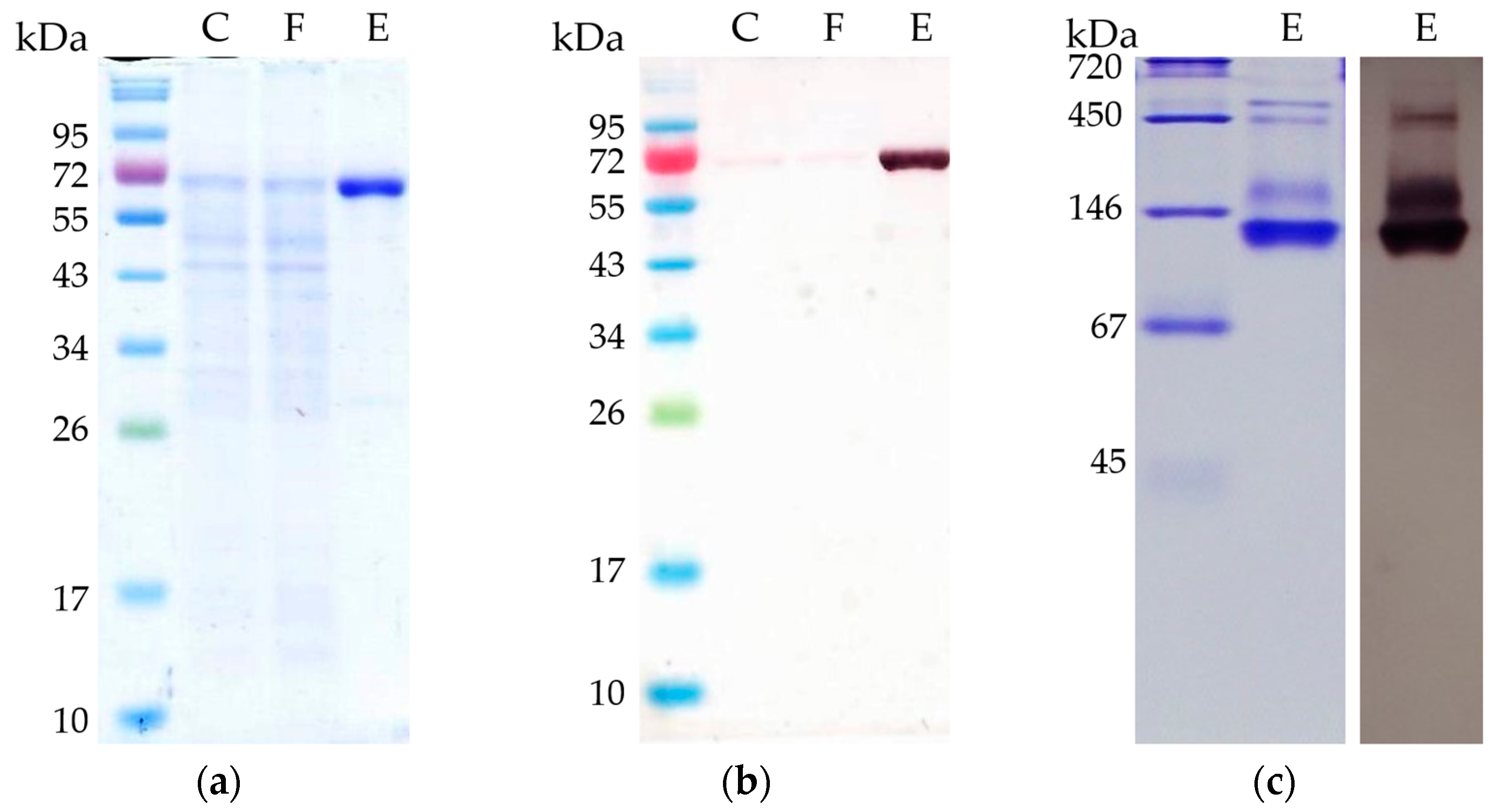
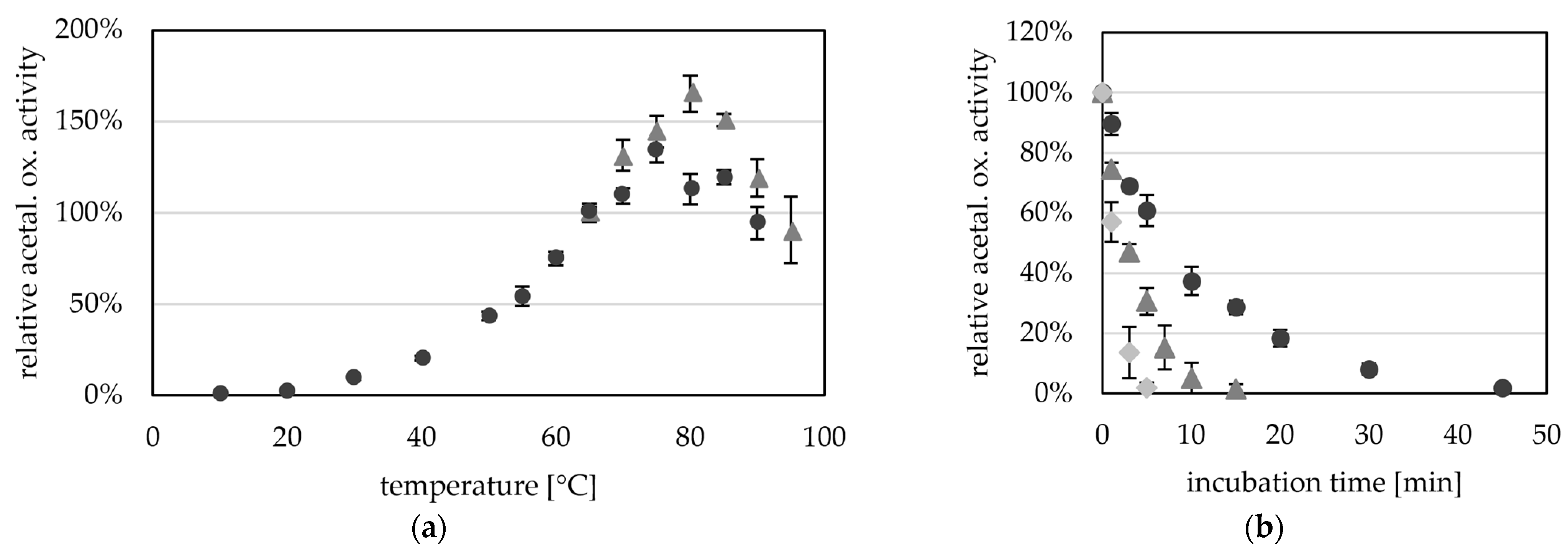
2.1. Recombinant AOR-His Obtained from a Related Strain Is Active
2.2. AOR-His Activity Can Be Improved by Media Optimization
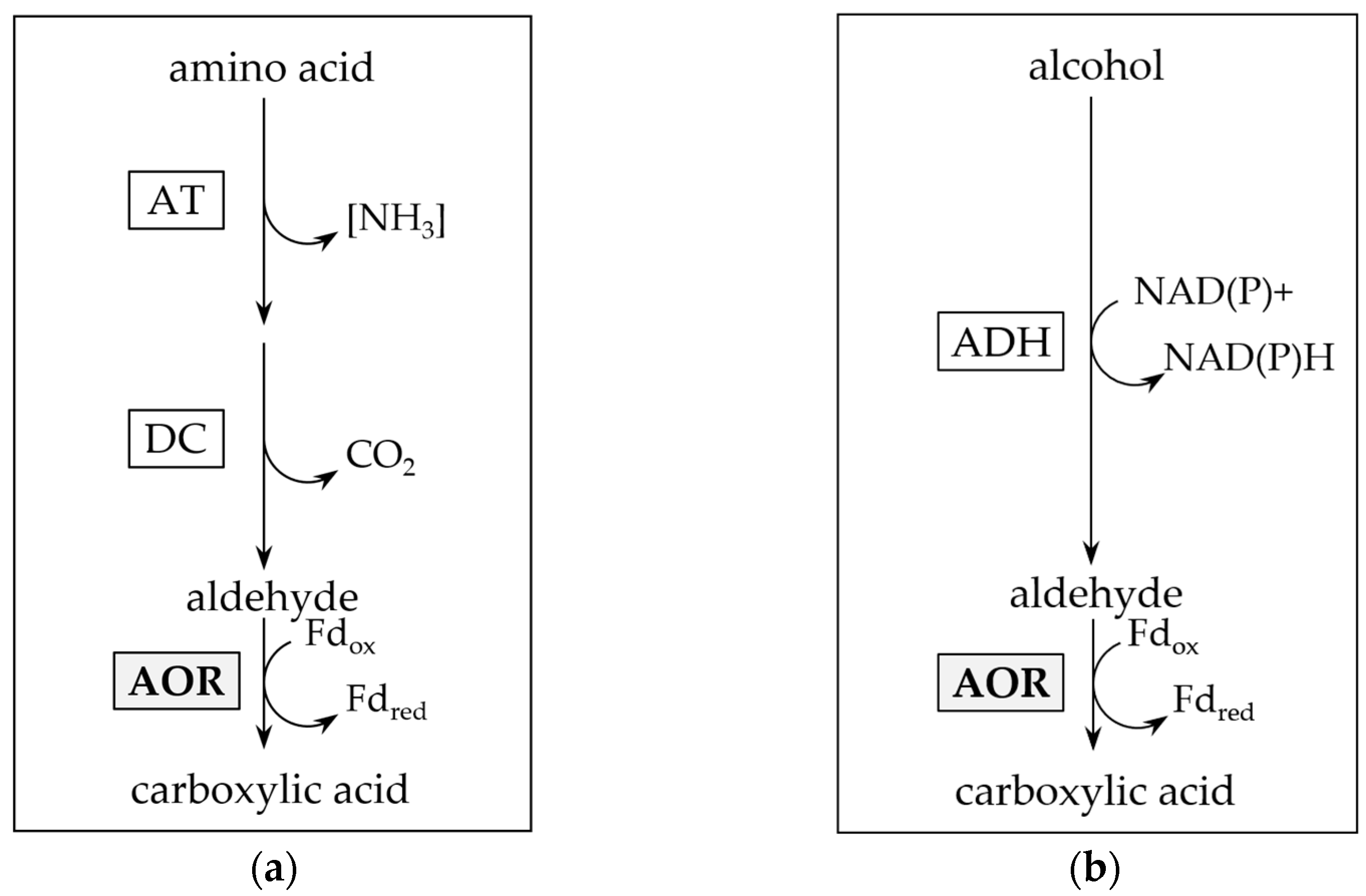
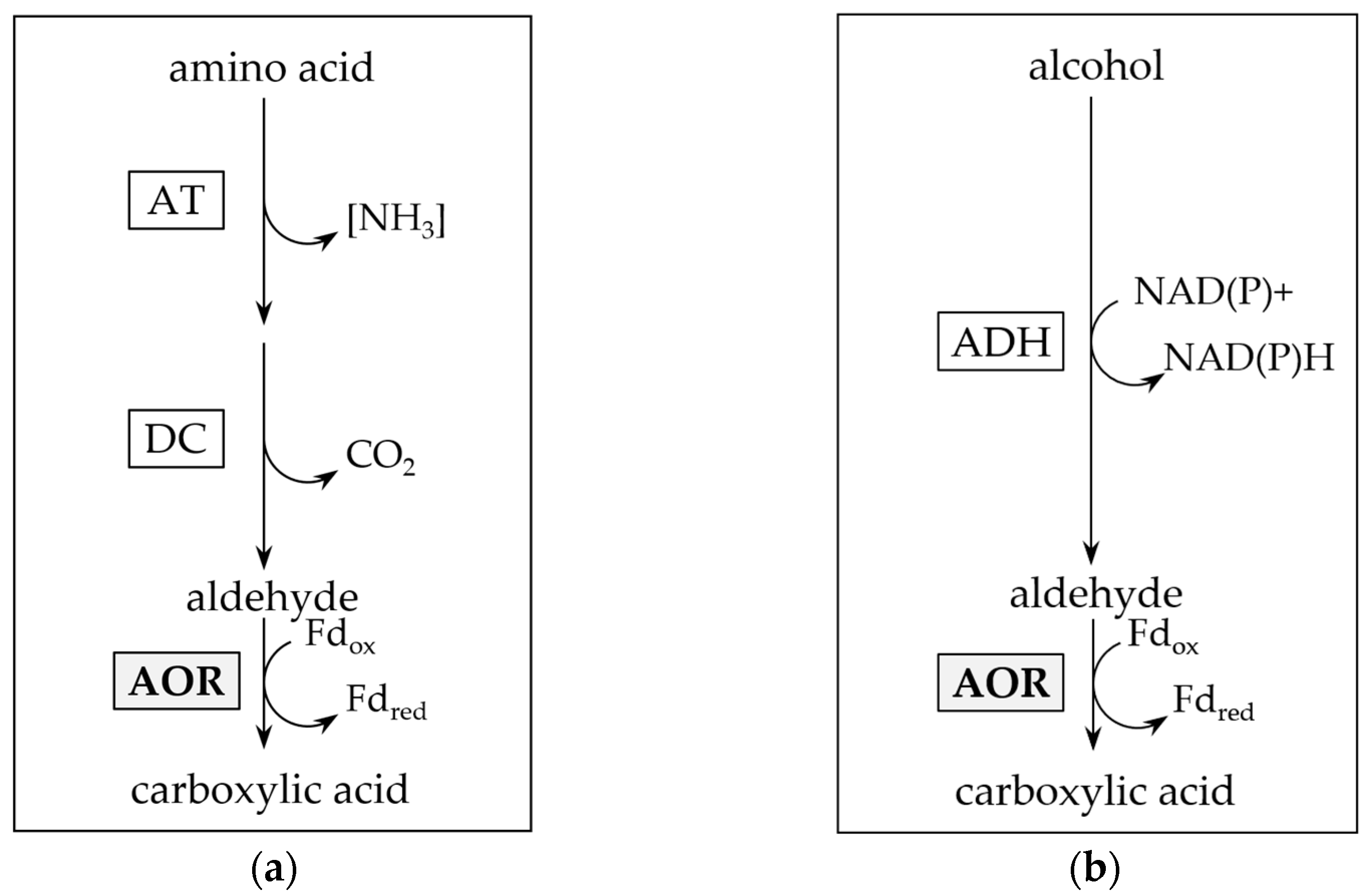
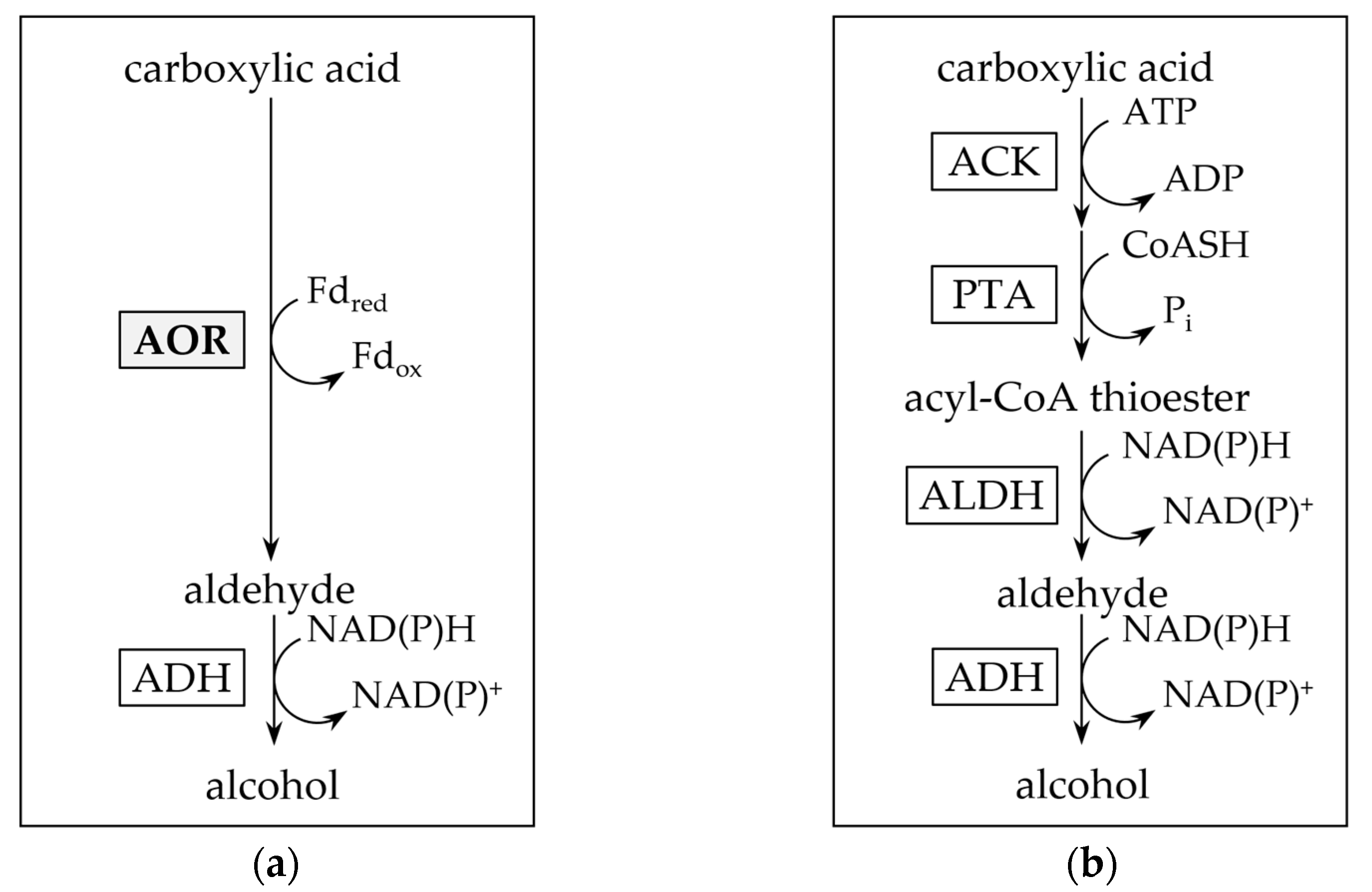
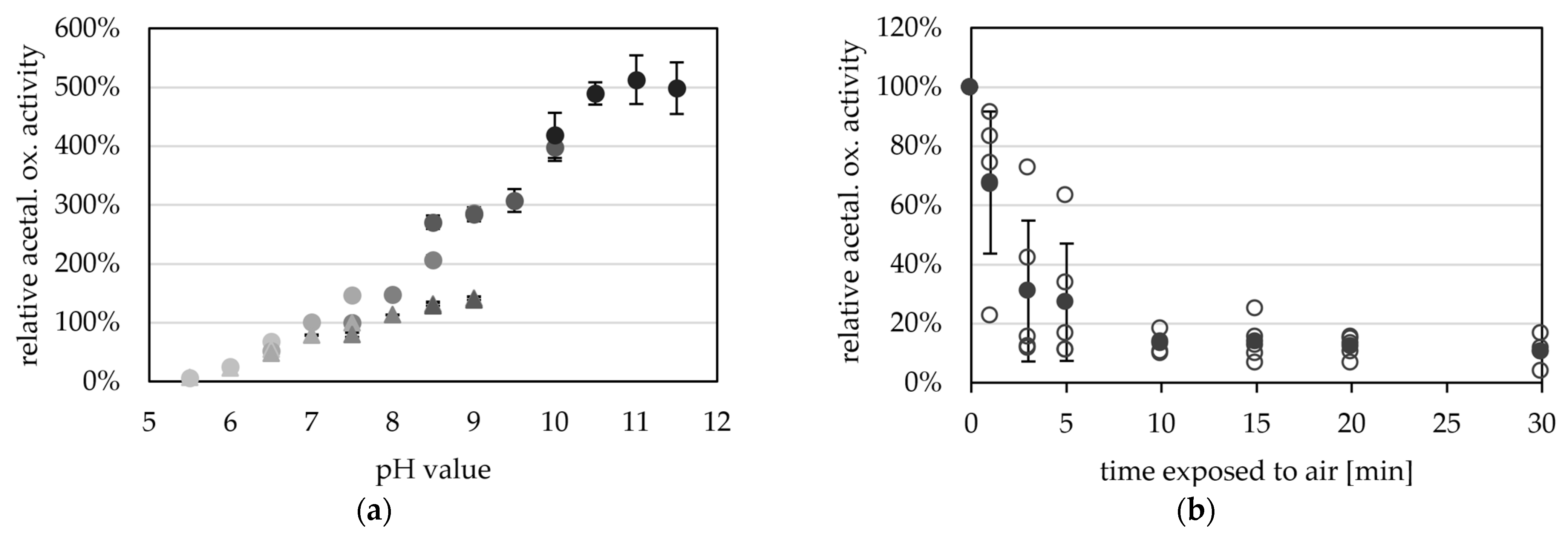
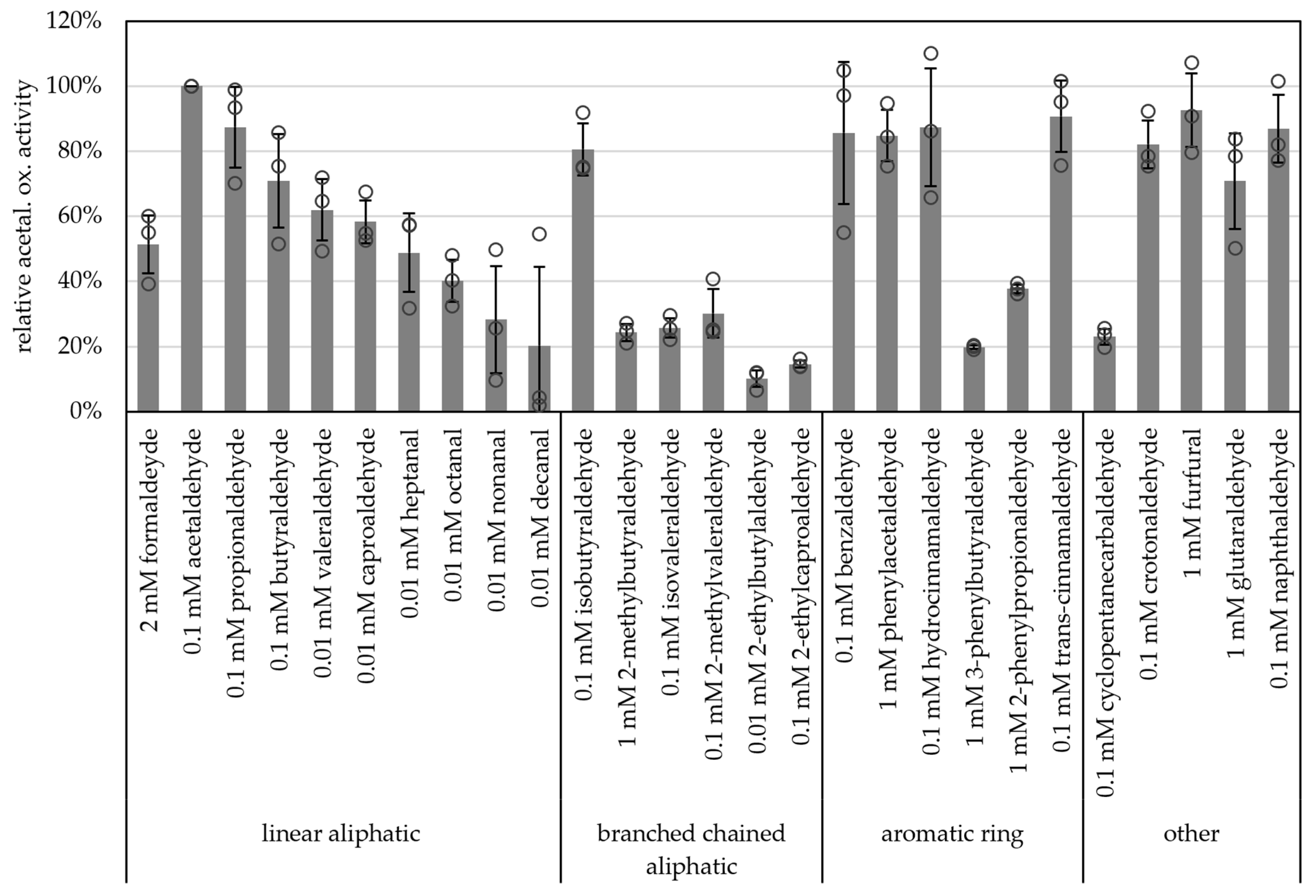
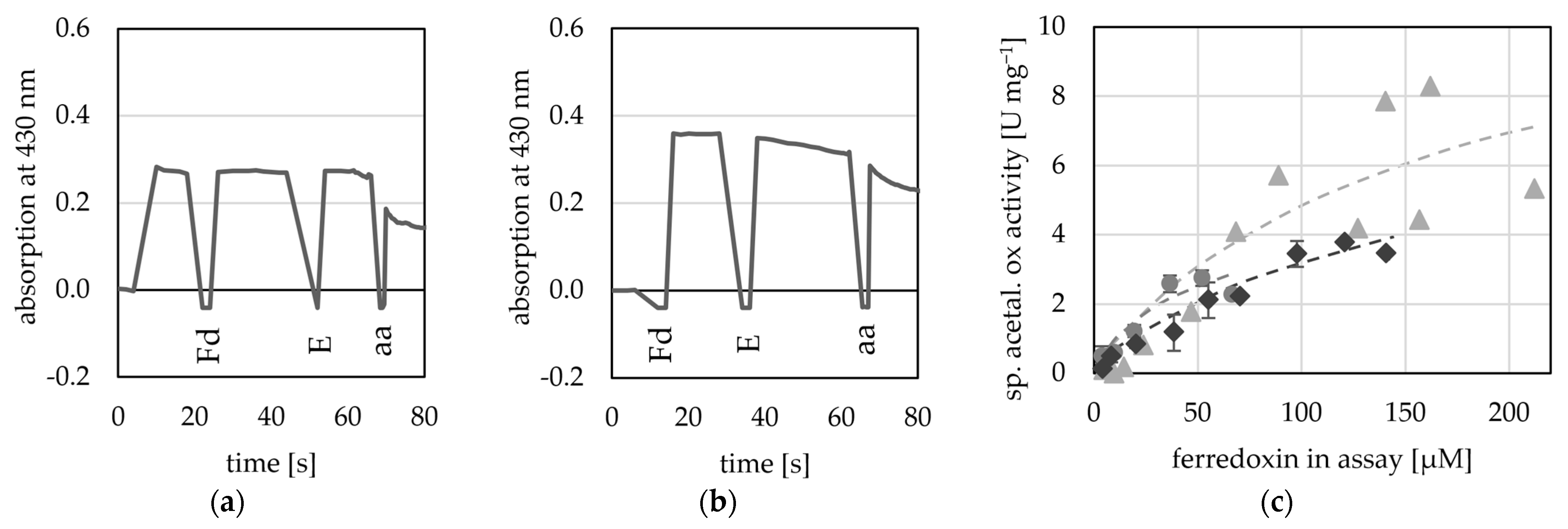
2.3. AORX514 Is a Versatile Ferredoxin-Dependent Aldehyde Oxidoreductase
3. Materials and Methods
3.1. Cultivation of Thermoanaerobacter kivui Strains
3.2. Transformation of Thermoanaerobacter kivui
3.3. Purification of His-Tagged Proteins from Thermoanaerobacter kivui
3.4. Enzyme Assays
3.5. Quantification of Elements
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, H.; Strobl, G.; Feicht, R.; Simon, H. Carboxylic acid reductase: A new tungsten enzyme catalyses the reduction of non-activated carboxylic acids to aldehydes. Eur. J. Biochem. 1989, 184, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Mukund, S.; Adams, M.W.W. The novel tungsten-iron-sulfur protein of the hyperthermophilic archaebacterium, Pyrococcus furiosus, is an aldehyde ferredoxin oxidoreductase: Evidence for its participation in a unique glycolytic pathway. J. Biol. Chem. 1991, 266, 14208–14216. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.K.; Mukund, S.; Kletzin, A.; Adams, M.W.W.; Rees, D.C. Structure of a hyperthermophilic tungstopterin enzyme, aldehyde ferredoxin oxidoreductase. Science 1995, 267, 1463–1469. [Google Scholar] [CrossRef] [PubMed]
- Huber, C.; Skopan, H.; Feicht, R.; White, H.; Simon, H. Pterin cofactor, substrate specificity, and observations on the kinetics of the reversible tungsten-containing aldehyde oxidoreductase from Clostridium thermoaceticum. Arch. Microbiol. 1995, 164, 110–118. [Google Scholar] [CrossRef]
- White, H.; Huber, C.; Feicht, R.; Simon, H. On a reversible molybdenum-containing aldehyde oxidoreductase from Clostridium formicoaceticum. Arch. Microbiol. 1993, 159, 244–249. [Google Scholar] [CrossRef]
- Trautwein, T.; Krauss, F.; Lottspeich, F.; Simon, H. The (2R)-hydroxycarboxylate-viologen-oxidoreductase from Proteus vulgaris is a molybdenum-containing iron-sulphur protein. Eur. J. Biochem. 1994, 222, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Strobl, G.; Feicht, R.; White, H.; Lottspeich, F.; Simon, H. The tungsten-containing aldehyde oxidoreductase from Clostridium thermoaceticum and its complex with a viologen-accepting NADPH oxidoreductase. Biol. Chem. Hoppe-Seyler 1992, 373, 123–132. [Google Scholar] [CrossRef]
- Winiarska, A.; Ramírez-Amador, F.; Hege, D.; Gemmecker, Y.; Prinz, S.; Hochberg, G.; Heider, J.; Szaleniec, M.; Schuller, J.M. A bacterial tungsten-containing aldehyde oxidoreductase forms an enzymatic decorated protein nanowire. bioRxiv 2023. [Google Scholar] [CrossRef]
- Rauh, D.; Graentzdoerffer, A.; Granderath, K.; Andreesen, J.R.; Pich, A. Tungsten-containing aldehyde oxidoreductase of Eubacterium acidaminophilum. Eur. J. Biochem. 2004, 271, 212–219. [Google Scholar] [CrossRef]
- Kim, G.; Azmi, L.; Jang, S.; Jung, T.; Hebert, H.; Roe, A.J.; Byron, O.; Song, J.-J. Aldehyde-alcohol dehydrogenase forms a high-order spirosome architecture critical for its activity. Nat. Commun. 2019, 10, 4527. [Google Scholar] [CrossRef]
- Dietrich, H.M.; Righetto, R.D.; Kumar, A.; Wietrzynski, W.; Trischler, R.; Schuller, S.K.; Wagner, J.; Schwarz, F.M.; Engel, B.D.; Müller, V.; et al. Membrane-anchored HDCR nanowires drive hydrogen-powered CO2 fixation. Nature 2022, 607, 823–830. [Google Scholar] [CrossRef] [PubMed]
- White, H.; Feicht, R.; Huber, C.; Lottspeich, F.; Simon, H. Purification and some properties of the tungsten-containing carboxylic acid reductase from Clostridium formicoaceticum. Biol. Chem. Hoppe-Seyler 1991, 372, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Heider, J.; Ma, K.; Adams, M.W.W. Purification, characterization, and metabolic function of tungsten-containing aldehyde ferredoxin oxidoreductase from the hyperthermophilic and proteolytic archaeon Thermococcus strain ES-1. J. Bacteriol. 1995, 177, 4757–4764. [Google Scholar] [CrossRef] [PubMed]
- Hensgens, C.M.; Hagen, W.R.; Hansen, T.A. Purification and characterization of a benzylviologen-linked, tungsten-containing aldehyde oxidoreductase from Desulfovibrio gigas. J. Bacteriol. 1995, 177, 6195–6200. [Google Scholar] [CrossRef] [PubMed]
- Hagedoorn, P.-L.; Chen, T.; Schröder, I.; Piersma, S.R.; de Vries, S.; Hagen, W.R. Purification and characterization of the tungsten enzyme aldehyde:ferredoxin oxidoreductase from the hyperthermophilic denitrifier Pyrobaculum aerophilum. J. Biol. Inorg. Chem. 2005, 10, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Arndt, F.; Schmitt, G.; Winiarska, A.; Saft, M.; Seubert, A.; Kahnt, J.; Heider, J. Characterization of an aldehyde oxidoreductase from the mesophilic bacterium Aromatoleum aromaticum EbN1, a member of a new subfamily of tungsten-containing enzymes. Front. Microbiol. 2019, 10, 71–83. [Google Scholar] [CrossRef]
- Debnar-Daumler, C.; Seubert, A.; Schmitt, G.; Heider, J. Simultaneous involvement of a tungsten-containing aldehyde:ferredoxin oxidoreductase and a phenylacetaldehyde dehydrogenase in anaerobic phenylalanine metabolism. J. Bacteriol. 2014, 196, 483–492. [Google Scholar] [CrossRef]
- Scully, S.M.; Iloranta, P.; Myllymaki, P.; Orlygsson, J. Branched-chain alcohol formation by thermophilic bacteria within the genera of Thermoanaerobacter and Caldanaerobacter. Extremophiles 2015, 19, 809–818. [Google Scholar] [CrossRef]
- Hensgens, C.M.H.; Nienhuis-Kuiper, M.E.; Hansen, T.A. Effects of tungstate on the growth of Desulfovibrio gigas NCIMB 9332 and other sulfate-reducing bacteria with ethanol as a substrate. Arch. Mikrobiol. 1994, 162, 143–147. [Google Scholar] [CrossRef]
- Hitschler, L.; Kuntz, M.; Langschied, F.; Basen, M. Thermoanaerobacter species differ in their potential to reduce organic acids to their corresponding alcohols. Appl. Microbiol. Biotechnol. 2018, 102, 8465–8476. [Google Scholar] [CrossRef]
- Nissen, L.S.; Basen, M. The emerging role of aldehyde:ferredoxin oxidoreductases in microbially-catalyzed alcohol production. J. Biotechnol. 2019, 306, 105–117. [Google Scholar] [CrossRef]
- Olson, D.G.; Sparling, R.; Lynd, L.R. Ethanol production by engineered thermophiles. Curr. Opin. Biotechnol. 2015, 33, 130–141. [Google Scholar] [CrossRef]
- Simon, H.; White, H.; Lebertz, H.; Thanos, I. Reduction of 2-enoates and alkanoates with carbon monoxide or formate, viologens, and Clostridium thermoaceticum to saturated acids and unsaturated and saturated alcohols. Angew. Chem. Int. Ed. Engl. 1987, 26, 785–787. [Google Scholar] [CrossRef]
- Basen, M.; Schut, G.J.; Nguyen, D.M.; Lipscomb, G.L.; Benn, R.A.; Prybol, C.J.; Vaccaro, B.J.; Poole, F.L.; Kelly, R.M.; Adams, M.W.W. Single gene insertion drives bioalcohol production by a thermophilic archaeon. Proc. Natl. Acad. Sci. USA 2014, 111, 17618–17623. [Google Scholar] [CrossRef] [PubMed]
- Mock, J.; Zheng, Y.; Mueller, A.P.; Ly, S.; Tran, L.; Segovia, S.; Nagaraju, S.; Köpke, M.; Dürre, P.; Thauer, R.K. Energy conservation associated with ethanol formation from H2 and CO2 in Clostridium autoethanogenum involving electron bifurcation. J. Bacteriol. 2015, 197, 2965–2980. [Google Scholar] [CrossRef] [PubMed]
- Liew, F.; Henstra, A.M.; Köpke, M.; Winzer, K.; Simpson, S.D.; Minton, N.P. Metabolic engineering of Clostridium autoethanogenum for selective alcohol production. Metab. Eng. 2017, 40, 104–114. [Google Scholar] [CrossRef]
- Perez, J.M.; Richter, H.; Loftus, S.E.; Angenent, L.T. Biocatalytic reduction of short-chain carboxylic acids into their corresponding alcohols with syngas fermentation. Biotechnol. Bioeng. 2013, 110, 1066–1077. [Google Scholar] [CrossRef]
- Richter, H.; Molitor, B.; Wei, H.; Chen, W.; Aristilde, L.; Angenent, L.T. Ethanol production in syngas-fermenting Clostridium ljungdahlii is controlled by thermodynamics rather than by enzyme expression. Energy Environ. Sci. 2016, 9, 2392–2399. [Google Scholar] [CrossRef]
- Scully, S.M.; Brown, A.E.; Ross, A.B.; Orlygsson, J. Biotransformation of organic acids to their corresponding alcohols by Thermoanaerobacter pseudoethanolicus. Anaerobe 2019, 57, 28–31. [Google Scholar] [CrossRef]
- Hitschler, L.; Nissen, L.S.; Kuntz, M.; Basen, M. Alcohol dehydrogenases AdhE and AdhB with broad substrate ranges are important enzymes for organic acid reduction in Thermoanaerobacter sp. strain X514. Biotechnol. Biofuels 2021, 14, 187. [Google Scholar] [CrossRef]
- Roy, R.; Menon, A.L.; Adams, M.W.W. Aldehyde oxidoreductases from Pyrococcus furiosus. Meth. Enzymol. 2001, 331, 132–144. [Google Scholar] [CrossRef]
- Kletzin, A.; Adams, M.W.W. Tungsten in biological systems. FEMS Microbiol. Rev. 1996, 18, 5–63. [Google Scholar] [CrossRef]
- Basen, M.; Geiger, I.; Henke, L.; Müller, V. A genetic system for the thermophilic acetogenic bacterium Thermoanaerobacter kivui. Appl. Environ. Microbiol. 2018, 84, e02210-17. [Google Scholar] [CrossRef]
- Leigh, J.A.; Mayer, F.; Wolfe, R.S. Acetogenium kivui, a new thermophilic hydrogen-oxidizing, acetogenic bacterium. Arch. Microbiol. 1981, 129, 275–280. [Google Scholar] [CrossRef]
- Weghoff, M.C.; Müller, V. CO metabolism in the thermophilic acetogen Thermoanaerobacter kivui. Appl. Environ. Microbiol. 2016, 82, 2312–2319. [Google Scholar] [CrossRef]
- Scott, I.M.; Rubinstein, G.M.; Lipscomb, G.L.; Basen, M.; Schut, G.J.; Rhaesa, A.M.; Lancaster, W.A.; Poole, F.L.; Kelly, R.M.; Adams, M.W.W. A new class of tungsten-containing oxidoreductase in Caldicellulosiruptor, a genus of plant biomass-degrading thermophilic bacteria. Appl. Environ. Microbiol. 2015, 81, 7339–7347. [Google Scholar] [CrossRef] [PubMed]
- Winiarska, A.; Hege, D.; Gemmecker, Y.; Kryściak-Czerwenka, J.; Seubert, A.; Heider, J.; Szaleniec, M. A tungsten enzyme using hydrogen as an electron donor to reduce carboxylic acids and NAD+. ACS Catal. 2022, 12, 8707–8717. [Google Scholar] [CrossRef] [PubMed]
- Schuchmann, K.; Müller, V. Direct and reversible hydrogenation of CO2 to formate by a bacterial carbon dioxide reductase. Biochemistry 2013, 342, 1382–1385. [Google Scholar] [CrossRef]
- Schuchmann, K.; Müller, V. Energetics and application of heterotrophy in acetogenic bacteria. Appl. Environ. Microbiol. 2016, 82, 4056–4069. [Google Scholar] [CrossRef]
- Jain, S.; Dietrich, H.M.; Müller, V.; Basen, M. Formate is required for growth of the thermophilic acetogenic bacterium Thermoanaerobacter kivui lacking hydrogen-dependent carbon dioxide reductase (HDCR). Front. Microbiol. 2020, 11, 59. [Google Scholar] [CrossRef]
- Katsyv, A.; Schoelmerich, M.C.; Basen, M.; Müller, V. The pyruvate:ferredoxin oxidoreductase of the thermophilic acetogen, Thermoanaerobacter kivui. FEBS Open Bio 2021, 11, 1332–1342. [Google Scholar] [CrossRef]
- Napora-Wijata, K.; Strohmeier, G.A.; Winkler, M. Biocatalytic reduction of carboxylic acids. Biotechnol. J. 2014, 9, 822–843. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, J.; Yuan, J.; Jiang, X.; Jiang, L.; Zhao, G.; Huang, D.; Liu, B. Omics-based analyses revealed metabolic responses of Clostridium acetobutylicum to lignocellulose-derived inhibitors furfural, formic acid and phenol stress for butanol fermentation. Biotechnol. Biofuels 2019, 12, 101. [Google Scholar] [CrossRef] [PubMed]
- Xia, A.; Feng, D.; Huang, Y.; Zhu, X.; Wang, Z.; Zhu, X.; Liao, Q. Activated carbon facilitates anaerobic digestion of furfural wastewater: Effect of direct interspecies electron transfer. ACS Sustain. Chem. Eng. 2022, 10, 8206–8215. [Google Scholar] [CrossRef]
- Roy, R.; Mukund, S.; Schut, G.J.; Dunn, D.M.; Weiss, R.; Adams, M.W.W. Purification and molecular characterization of the tungsten-containing formaldehyde ferredoxin oxidoreductase from the hyperthermophilic archaeon Pyrococcus furiosus: The third of a putative five-member tungstoenzyme family. J. Bacteriol. 1999, 181, 1171–1180. [Google Scholar] [CrossRef]
- Mukund, S.; Adams, M.W.W. Characteriztion of a novel tugsten containing formadehyde ferrodoxin oxidoreductase from the hyperthermophilic Archaeon. Thermococcus litoralis. J. Biol. Chem. 1993, 268, 13592–13600. [Google Scholar] [CrossRef] [PubMed]
- Wardman, P. The reduction potential of benzyl viologen: An important reference compound for oxidant/radical redox couples. Free Radic. Res. Commun. 1991, 14, 57–67. [Google Scholar] [CrossRef]
- Katsyv, A.; Essig, M.; Bedendi, G.; Sahin, S.; Milton, R.D.; Müller, V. Characterization of ferredoxins from the thermophilic, acetogenic bacterium Thermoanaerobacter kivui. FEBS J. 2023, 290, 4107–4125. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed]
- Beinert, H. Semi-micro methods for analysis of labile sulfide and of labile sulfide plus sulfane sulfur in unusually stable iron-sulfur proteins. Anal. Biochem. 1983, 131, 373–378. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tungsten | Molybdenum | Iron | Sulfur | Yeast Extract | Glucose | Relative Aldehyde Oxidation Activity |
|---|---|---|---|---|---|---|
| 12 µM | 490 nM | 11 µM | 2 mM SO42− | 0.2% | 25 mM | 100% |
| 1.2 nM | 490 nM | 11 µM | 2 mM SO42− | 0.2% | 25 mM | 0.2% |
| 12 nM | 490 nM | 11 µM | 2 mM SO42− | 0.2% | 25 mM | 0.5% |
| 120 nM | 490 nM | 11 µM | 2 mM SO42− | 0.2% | 25 mM | 65% |
| 1.2 µM | 490 nM | 11 µM | 2 mM SO42− | 0.2% | 25 mM | 75% |
| 120 µM | 490 nM | 11 µM | 2 mM SO42− | 0.2% | 25 mM | 98% |
| 1.2 mM | 490 nM | 11 µM | 2 mM SO42− | 0.2% | 25 mM | 113% |
| 120 nM | w/o | 11 µM | 2 mM SO42− | 0.2% | 25 mM | 79% |
| 120 nM | 4.9 µM | 11 µM | 2 mM SO42− | 0.2% | 25 mM | 63% |
| 120 nM | 49 µM | 11 µM | 2 mM SO42− | 0.2% | 25 mM | 4% |
| 12 µM | 490 nM | 83 µM | 2 mM SO42− | 0.2% | 25 mM | 160% |
| 12 µM | 490 nM | 11 µM | 11 mM SO4− | 0.2% | 25 mM | 132% |
| 12 µM | 490 nM | 11 µM | +1 mM SO32− * | 0.2% | 25 mM | 66% |
| 12 µM | 490 nM | 11 µM | +1 mM S2O42− * | 0.2% | 25 mM | 95% |
| 12 µM | 490 nM | 11 µM | +1 mM S2O32− * | 0.2% | 25 mM | 55% |
| 12 µM | 490 nM | 11 µM | 2 mM SO42− | 1% | 25 mM | 191% |
| 12 µM | 490 nM | 11 µM | 2 mM SO42− | 1% | 100 mM | 164% |
| 12 µM | 490 nM | 11 µM | 2 mM SO42− | 0.2% | 100 mM | 53% |
| kM [µM] | Vmax [µmol min−1 mg−1] | |
|---|---|---|
| BV | 2122 | 17.7 |
| MV | 19230 | 19.6 |
| His-TKV_c09620 (Fd) | ~50 to 200 | ~8 to 19 |
| Formaldehyde * | 93.2 | 25.3 |
| Acetaldehyde * | 9.7 | 38.0 |
| Propionaldehyde * | 18.8 | 38.5 |
| Butyraldehyde * | 42.3 | 38.0 |
| Valeraldehyde * | 6.8 | 23.4 |
| Capronaldehyde * | 2.8 | 18.5 |
| Heptanal * | 7.4 | 15.3 |
| Octanal * | 17.1 | 17.8 |
| Redox Partner | Relative Specific AOR Activity | |
|---|---|---|
| acetaldehyde oxidation | 2 mM BV | 100% |
| 19 mM MV | 56% | |
| 20 µM His-Fd09 | 4.4% | |
| 20 µM His-Fd16 | 3.5% | |
| acetate reduction | 10 µM MV (uncoupled) | 0.03% |
| 10 µM MV (coupled) | 0.04% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nissen, L.S.; Moon, J.; Hitschler, L.; Basen, M. A Versatile Aldehyde: Ferredoxin Oxidoreductase from the Organic Acid Reducing Thermoanaerobacter sp. Strain X514. Int. J. Mol. Sci. 2024, 25, 1077. https://doi.org/10.3390/ijms25021077
Nissen LS, Moon J, Hitschler L, Basen M. A Versatile Aldehyde: Ferredoxin Oxidoreductase from the Organic Acid Reducing Thermoanaerobacter sp. Strain X514. International Journal of Molecular Sciences. 2024; 25(2):1077. https://doi.org/10.3390/ijms25021077
Chicago/Turabian StyleNissen, Laura Sofie, Jimyung Moon, Lisa Hitschler, and Mirko Basen. 2024. "A Versatile Aldehyde: Ferredoxin Oxidoreductase from the Organic Acid Reducing Thermoanaerobacter sp. Strain X514" International Journal of Molecular Sciences 25, no. 2: 1077. https://doi.org/10.3390/ijms25021077
APA StyleNissen, L. S., Moon, J., Hitschler, L., & Basen, M. (2024). A Versatile Aldehyde: Ferredoxin Oxidoreductase from the Organic Acid Reducing Thermoanaerobacter sp. Strain X514. International Journal of Molecular Sciences, 25(2), 1077. https://doi.org/10.3390/ijms25021077