
Recombinase Polymerase Amplification Combined with Lateral Flow Dipstick Assay for the Rapid and Sensitive Detection of Pseudo-nitzschia multiseries

 and
and
Abstract
1. Introduction
2. Results
2.1. Recombinase Polymerase Amplification Primer and Probe Design, Screening, and System Optimization
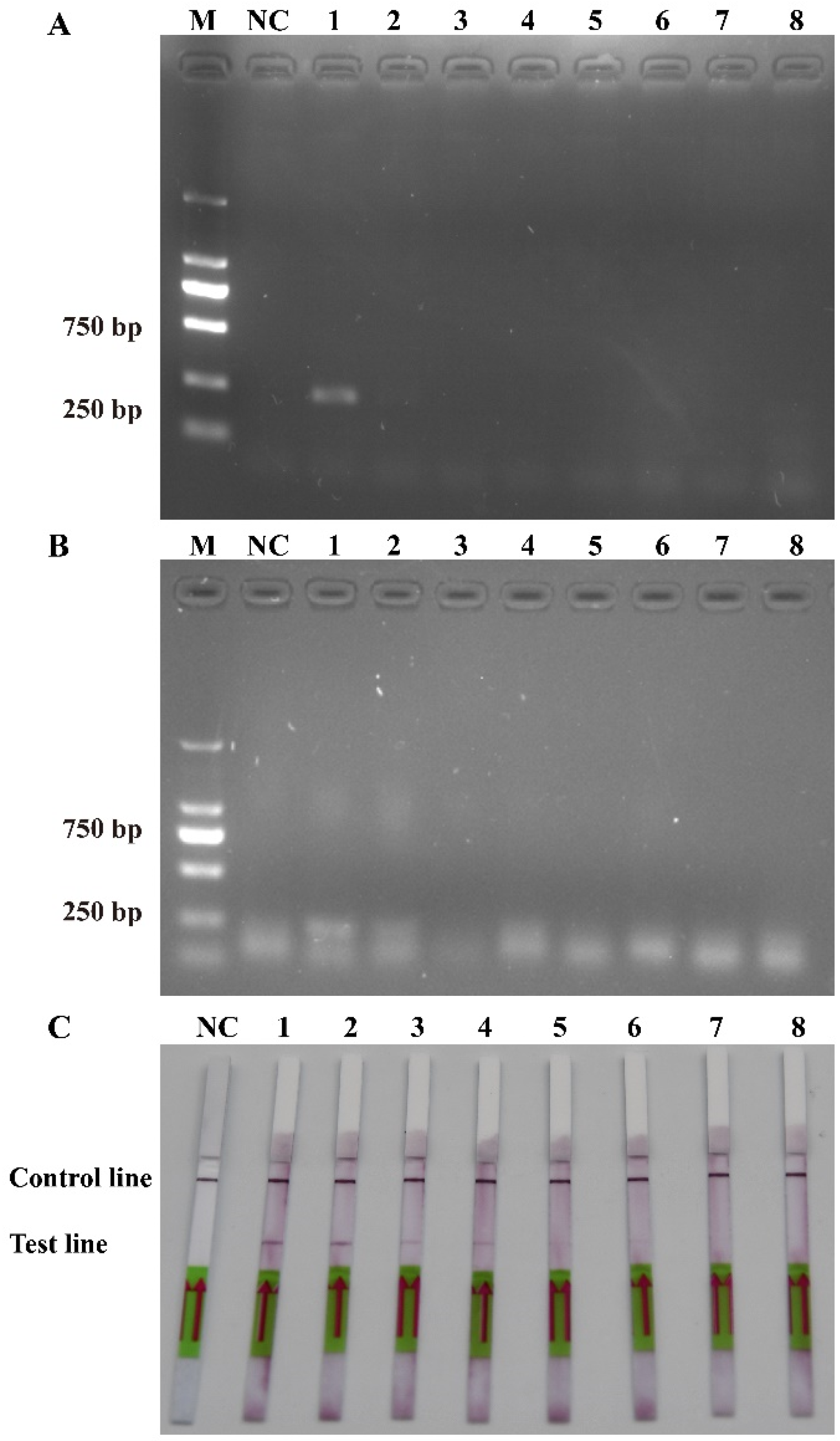
2.2. Recombinase Polymerase Amplification and Lateral Flow Dipstick Specificity
2.3. Recombinase Polymerase Amplification and Lateral Flow Dipstick Sensitivity
2.4. Evaluation of Recombinase Polymerase Amplification and Lateral Flow Dipstick
3. Materials and Methods
3.1. Algal Species and Cultures
3.2. DNA Extraction, Polymerase Chain Reaction, Cloning, and Sequencing
3.3. Recombinase Polymerase Amplification Primers and Lateral Flow Dipstick Probe Design
3.4. Recombinase Polymerase Amplification Conditions Optimization
3.5. Recombinase Polymerase Amplification and Lateral Flow Dipstick Assay
3.6. Specificity and Sensitivity Analysis
3.7. Recombinase Polymerase Amplification and Lateral Flow Dipstick Test of the Simulated and Natural Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gu, H.; Wu, Y.; Lü, S.; Lu, D.; Tang, Y.; Qi, Y. Emerging harmful algal bloom species over the last four decades in China. Harmful Algae 2022, 111, 102059. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.L.; Xu, S.S.; Li, S.Q.; Wang, X.W.; Guo, K.L.; Yan, R.M.; Xie, W.; Yin, K.D.; Hou, S.W.; Jiang, H.B. Diversity and distribution of harmful algal bloom species from seamount to coastal waters in the South China Sea. Microbiol. Spectr. 2023, 11, e0416922. [Google Scholar] [CrossRef] [PubMed]
- Kazmi, S.S.U.H.; Yapa, N.; Karunarathna, S.C.; Suwannarach, N. Perceived intensification in harmful algal blooms is a wave of cumulative threat to the aquatic ecosystems. Biology 2022, 11, 852. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.L.; Shao, Q.W.; Zhu, X.J.; Luo, J.; Meng, R.; Zhou, C.X.; Zhu, P.; Zhu, Y.F.; Yan, X.J. Distribution of Karlodinium veneficum in the coastal region of Xiangshan Bay in the East China Sea, as detected by a real-time quantitative PCR assay of ribosomal ITS sequence. Harmful Algae 2019, 81, 65–76. [Google Scholar] [CrossRef]
- Xu, D.; Zheng, G.; Brennan, G.; Wang, Z.; Jiang, T.; Sun, K.; Fan, X.; Bowler, C.; Zhang, X.; Zhang, Y.; et al. Plastic responses lead to increased neurotoxin production in the diatom Pseudo-nitzschia under ocean warming and acidification. ISME J. 2023, 17, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Ji, N.; Luo, H. Recent progress in marine harmful algal bloom research. Oceanol. Limnol. Sin. 2019, 50, 496–508. [Google Scholar]
- Chen, Y. Metabarcoding Dissection and Comparative Genomic Analysis of Harmful Algal Bloom Species in the East China Sea. Ph.D. Thesis, University of Chinese Academy of Sciences, Marine Biology, Qingdao, China, 2021. [Google Scholar] [CrossRef]
- Huang, C.X.; Dong, H.C.; Lundholm, N.; Teng, S.T.; Zheng, G.C.; Tan, Z.J.; Lim, P.T.; Li, Y. Species composition and toxicity of the genus Pseudo-nitzschia in Taiwan Strait, including P. chiniana sp. nov. and P. qiana sp. nov. Harmful Algae 2019, 84, 195–209. [Google Scholar] [CrossRef]
- Dong, H.C.; Lundholm, N.; Teng, S.T.; Li, A.F.; Wang, C.; Hu, Y.; Li, Y. Occurrence of Pseudo-nitzschia species and associated domoic acid production along the Guangdong coast, South China Sea. Harmful Algae 2020, 98, 101899. [Google Scholar] [CrossRef]
- Subba Rao, D.V.; Quilliam, M.A.; Pocklington, R. Domoic acid—A neurotoxic amino acid produced by the marine diatom Nitzschia pungens in culture. Can. J. Fish. Aquat. Sci. 1988, 45, 2076–2079. [Google Scholar] [CrossRef]
- Bates, S.S.; Bird, C.J.; de Freitas, A.S.W.; Foxall, R.; Gilgan, M.; Hanic, L.A.; Johnson, G.R.; McCulloch, A.W.; Odense, P.; Pocklington, R.; et al. Pennate diatom Nitzschia pungens as the primary source of domoic acid, a toxin in shellfish from eastern Prince Edward Island, Canada. Can. J. Fish. Aquat. Sci. 1989, 46, 1203–1215. [Google Scholar] [CrossRef]
- Quilliam, M.A.; Wright, J.L.C. The amnesic shellfish poisoning mystery. Anal. Chem. 1989, 61, 1053A–1060A. [Google Scholar] [CrossRef] [PubMed]
- Silver, M.W.; Bargu, S.; Coale, S.L. Toxic diatoms and domoic acid in natural and iron enriched waters of the oceanic pacific. Proc. Natl. Acad. Sci. USA 2010, 107, 20762–20767. [Google Scholar] [CrossRef] [PubMed]
- Trick, C.G.; Bill, B.D.; Cochlan, W.P.; Wells, M.L.; Trainer, V.L.; Pickell, L.D. Iron enrichment stimulates toxic diatom production in high-nitrate, low-chlorophyll areas. Proc. Natl. Acad. Sci. USA 2010, 107, 5887–5892. [Google Scholar] [CrossRef] [PubMed]
- Bates, S.S.; Hubbard, K.A.; Lundholm, N.; Montresor, M.; Leaw, C.P. Pseudo-nitzschia, Nitzschia, and domoic acid: New research since 2011. Harmful Algae 2018, 79, 3–43. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Wu, H.; Che, H.; Li, X.; Zhang, Z.; Peng, J.; Guo, M.; Tan, Z. Residue analysis and assessment of the risk of dietary exposure to domoic acid in shellfish from the coastal areas of China. Toxins 2022, 14, 862. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Murray, J.S.; Boundy, M.J.; Balci, M.; Bowers, H.A.; Smith, K.F.; Harwood, D.T.; Rhodes, L.L. Update of the planktonic diatom genus Pseudo-nitzschia in aotearoa New Zealand coastal waters: Genetic diversity and toxin production. Toxins 2021, 13, 637. [Google Scholar] [CrossRef] [PubMed]
- Antonella, P.; Elena, B.; Cecilia, B.; Luca, G.; Maria, G.G.; Magda, V.; Esther, G.; Antonella, L.; Mauro, M. Monitoring of HAB species in the Mediterranean Sea through molecular methods. J. Plankton Res. 2007, 29, 19–38. [Google Scholar] [CrossRef]
- Fitzpatrick, E.; Caron, D.A.; Schnetzer, A. Development and environmental application of a genus-specific quantitative PCR approach for Pseudo-nitzschia species. Mar. Biol. 2010, 157, 1161–1169. [Google Scholar] [CrossRef]
- Delaney, J.A.; Ulrich, R.M.; Paul, J.H. Detection of the toxic marine diatom Pseudo-nitzschia multiseries using the RuBisCO small subunit (rbcS) gene in two real-time RNA amplification formats. Harmful Algae 2011, 11, 54–64. [Google Scholar] [CrossRef]
- Andree, K.B.; Fernandez-Tejedor, M.; Elandaloussi, L.M.; Quijano-Scheggia, S.; Sampedro, N.; Garces, E.; Camp, J.; Dioge‘ne, J. Quantitative PCR coupled with melt curve analysis for detection of selected pseudo-nitzschia spp. (Bacillariophyceae) from the northwestern mediterranean sea. Appl. Environ. Microbiol. 2011, 77, 1651–1659. [Google Scholar] [CrossRef]
- Chin Chwan Chuong, J.J.; Rahman, M.; Ibrahim, N.; Heng, L.Y.; Tan, L.L.; Ahmad, A. Harmful microalgae detection: Biosensors versus some conventional methods. Sensors 2022, 22, 3144. [Google Scholar] [CrossRef]
- Zingg, J.M.; Daunert, S. Trinucleotide rolling circle amplification: A novel method for the detection of RNA and DNA. Methods Protoc. 2018, 1, 15. [Google Scholar] [CrossRef]
- Liu, M.; Li, C.C.; Luo, X.L.; Ma, F.; Zhang, C.Y. 5-hydroxymethylcytosine glucosylation-triggered helicase-dependent amplification based fluorescent biosensor for sensitive detection of b-glucosyltransferase with zero background signal. Anal. Chem. 2020, 92, 16307–16313. [Google Scholar] [CrossRef]
- Nai, Y.H.; Doeven, E.H.; Guijt, R.M. An improved nucleic acid sequence-based amplification method mediated by T4 gene 32 protein. PLoS ONE 2022, 17, e0265391. [Google Scholar] [CrossRef]
- Huang, H.L.; Zhu, P.; Zhou, C.X.; He, S.; Yan, X.J. The development of loop mediated isothermal amplification combined with lateral flow dipstick for detection of Karlodinium veneficum. Harmful Algae 2017, 62, 20–29. [Google Scholar] [CrossRef]
- Xu, M.T.; Zhang, C.Y.; Liu, F.G.; Wang, Y.Y.; Li, R.Q.; Chen, G.F. Recombinase-aided amplification combined with lateral flow dipstick for the rapid detection of Amphidinium carterae. J. Appl. Phycol. 2022, 34, 435–447. [Google Scholar] [CrossRef]
- Luo, N.J.; Huang, H.L.; Jiang, H.B. Establishment of methods for rapid detection of Prymnesium parvum by recombinase polymerase amplification combined with a lateral flow dipstick. Front. Mar. Sci. 2022, 9, 1032847. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, C.; Wang, Y.; Chen, G. A review of the current and emerging detection methods of marine harmful microalgae. Sci. Total Environ. 2022, 815, 152913. [Google Scholar] [CrossRef]
- Toldrà, A.; O’Sullivan, C.K.; Campàs, M. Detecting harmful algal blooms with isothermal molecular strategies. Trends Biotechnol. 2019, 37, 1278–1281. [Google Scholar] [CrossRef]
- Toldrà, A.; O’Sullivan, C.K.; Diogène, J.; Campàs, M. Detecting harmful algal blooms with nucleic acid amplification-based biotechnological tools. Sci. Total Environ. 2020, 749, 141605. [Google Scholar] [CrossRef]
- Hosam, Z.; Mahmoud, E.S. Recombinase polymerase amplification as a promising tool in Hepatitis C virus diagnosis. World J. Hepatol. 2014, 6, 916–922. [Google Scholar] [CrossRef]
- Boyle, D.; McNerney, R.; Teng Low, H.; Leader, B.; Pérez-Osorio, A. Rapid detection of mycobacterium tuberculosis by recombinase polymerase amplification. PLoS ONE 2014, 9, e1030919. [Google Scholar] [CrossRef]
- Abukhalid, N.; Pastey, M.K. Nucleic acid amplification using recombinase polymerase: Enzymatic approach. J. Med. Microbiol. Diagn. 2017, 6, 1–3. [Google Scholar] [CrossRef]
- Ghosh, D.K.; Kokane, S.B.; Kokane, A.D.; Warghane, A.J.; Motghare, M.R.; Bhose, S.; Sharma, A.K.; Reddy, M.K. Development of a recombinase polymerase based isothermal amplification combined with lateral flow assay (HLB-RPA-LFA) for rapid detection of “Candidatus Liberibacter asiaticus”. PLoS ONE 2018, 13, e0208530. [Google Scholar] [CrossRef]
- Ghosh, D.K.; Bhose, S.; Warghane, A.; Motghare, M.; Sharma, A.K.; Dhar, A.K.; Gowda, S. Loop-mediated isothermal amplification LAMP based method for rapid and sensitive detection of ‘Candidatus Liberibacter asiaticus’ in citrus and the psyllid vector, Diaphorinacitri Kuwayama. J. Plant Biochem. Biotechnol. 2016, 25, 219–223. [Google Scholar] [CrossRef]
- Piepenburg, O.; Williams, C.H.; Stemple, D.L.; Armes, N.A. DNA detection using recombination proteins. PLoS Biol. 2006, 4, e204. [Google Scholar] [CrossRef]
- Qiao, M.; Zhang, L.; Chang, J.; Li, H.; Li, J.; Wang, W.; Yuan, G.; Su, J. Rapid and sensitive detection of pathogenic Elizabethkingia miricola in black spotted frog by RPA-LFD and fluorescent probe-based RPA. Fish Shellfish Immunol. Rep. 2022, 3, 100059. [Google Scholar] [CrossRef]
- Pang, J.H.; Wang, Q.; Fei, Y.J.; Zhu, P.; Qiao, L.L.; Huang, H.L.; Dang, C.Y.; Gao, W.F. A real-time recombinase polymerase amplification assay for the rapid detection of Vibrio harveyi. Mol. Cell. Probes 2019, 44, 8–13. [Google Scholar] [CrossRef]
- Mabrok, M.; Elayaraja, S.; Chokmangmeepisarn, P.; Jaroenram, W.; Arunrut, N.; Kiatpathomchai, W.; Debnath, P.P.; Delamare-Deboutteville, J.; Mohan, C.V.; Fawzy, A.; et al. Rapid visualization in the specific detection of flavobacterium columnare, a causative agent of freshwater columnaris using a novel recombinase polymerase amplification (RPA) combined with lateral flow dipstick (LFD) assay. Aquaculture 2021, 531, 735780. [Google Scholar] [CrossRef]
- Onchan, W.; Ritbamrung, O.; Changtor, P.; Pradit, W.; Chomdej, S.; Nganvongpanit, K.; Siengdee, P.; Suyasunanont, U.; Buddhachat, K. Sensitive and rapid detection of Babesia species in dogs by recombinase polymerase amplification with lateral flow dipstick (RPA-LFD). Sci. Rep. 2022, 12, 20560. [Google Scholar] [CrossRef]
- Bian, Z.; Liu, W.; Jin, J.; Hao, Y.; Jiang, L.; Xie, Y.; Zhang, H. Development of a recombinase polymerase amplification assay with lateral flow dipstick (RPA-LFD) for rapid detection of Shigella spp. and enteroinvasive Escherichia coli. PLoS ONE 2022, 17, e0278869. [Google Scholar] [CrossRef]
- Chen, Z.; Jiao, B.; Zhou, J.; He, H.; Dai, T. Rapid detection of Phytophthora cinnamomic based on a new target gene Pcinn13739. Front. Cell. Infect. Microbiol. 2022, 12, 923700. [Google Scholar] [CrossRef]
- Fu, M.Q.; Chen, G.F.; Zhang, C.Y.; Wang, Y.Y.; Sun, R.; Zhou, J. Rapid and sensitive detection method for Karlodinium veneficum by recombinase polymerase amplification coupled with lateral flow dipstick. Harmful Algae 2019, 84, 1–9. [Google Scholar] [CrossRef]
- Zhou, S.; Zheng, X.; Yang, Z.; Huang, Q.; Yi, J.; Su, L.; Guo, B.; Xiu, Y. Development of two recombinase polymerase amplification EXO (RPA-EXO) and lateral flow dipstick (RPA-LFD) techniques for the rapid visual detection of Aeromonas salmonicida. Mar. Biotechnol. 2022, 24, 1094–1109. [Google Scholar] [CrossRef]
- Lillis, L.; Siverson, J.; Lee, A.; Cantera, J.; Parker, M.; Piepenburg, O.; Lehman, D.A.; Boyle, D.S. Factors influencing recombinase polymerase amplification (RPA) assay outcomes at point of care. Mol. Cell. Probes 2016, 30, 74–78. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, J., Sainsky, J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Diercks, S.; Metfies, K.; Medlin, L.K. Molecular probe sets for the detection of toxic algae for use in sandwich hybridization formats. J. Plankton Res. 2008, 30, 439–448. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Yang, Y.C.; Liu, F.G.; Wang, Y.Y.; Chen, G.F. Recombinase polymerase amplification combined with lateral flow dipstick for the rapid detection of Chattonella marina. J. Appl. Phycol. 2022, 34, 1607–1620. [Google Scholar] [CrossRef]
- Hubbard, K.A.; Rocap, G.; Armbrust, E.V. Inter- and intraspecific community structure within the diatom genus Pseudo-nitzschia (Bacillariophyceae). J. Phycol. 2008, 44, 637–649. [Google Scholar] [CrossRef]
- Kim, J.M.; Lim, S.-J.; Truong, T.A.; Wang, J.-H.; Lee, C.-W.; Yoon, B. Comparison Between Specific DNA-amplifications using Recombinase Polymerase Amplification (RPA) and using Polymerase Chain Reaction (PCR). J. Apic. 2017, 32, 41–50. [Google Scholar] [CrossRef]
- Zhai, Y.D.; Li, R.Q.; Liu, F.G.; Zhang, C.Y.; Wang, Y.Y.; Chen, G.F. Recombinase polymerase amplification combined with lateral flow dipstick for the rapid detection of Prorocentrum donghaiense. Mar. Biol. Res. 2021, 17, 646–657. [Google Scholar] [CrossRef]
- Tomar, S.; Lavickova, B.; Guiducci, C. Recombinase polymerase amplification in minimall buffered conditions. Biosens. Bioelectron. 2022, 198, 113802. [Google Scholar] [CrossRef]
- Luo, G.C.; Yi, T.T.; Jiang, B.; Guo, X.L.; Zhang, G.Y. Betaine-assisted recombinase polymerase assay with enhanced specificity. Anal. Biochem. 2019, 575, 36–39. [Google Scholar] [CrossRef]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef]
- Williams, M.A.; O’Grady, J.; Bernard, B.; Jens, C.; de Eyto, E.; McGinnity, P.; Jennings, E.; Regan, F.; McDermott, A.P. The application of CRISPR-Cas for single species identification from environmental DNA. Mol. Ecol. Resour. 2019, 19, 1106–1114. [Google Scholar] [CrossRef]
- Wang, Y.; Ke, Y.; Liu, W.; Sun, Y.Q.; Ding, X.T. A one-pot toolbox based on Cas12a/crRNA enables rapid foodborne pathogen detection at attomolar level. ACS Sens. 2020, 5, 1427–1435. [Google Scholar] [CrossRef]
- Zhang, W.S.; Pan, J.; Li, F.; Zhu, M.; Su, G. Reverse transcription recombinase polymerase amplification coupled with CRISPR-Cas12a for facile and highly sensitive colorimetric SARS-CoV-2 detection. Anal. Chem. 2021, 93, 4126–4133. [Google Scholar] [CrossRef]
- Chen, J.S.; Ma, E.; Harrington, L.B.; Da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef]
- Gootenberg, J.S.; Abudayyeh, O.O.; Kellner, M.J.; Joung, J.L.; Collins, J.J.; Zhang, F. Multiplexed and portable nucleic acid detection platform with Cas13, Cas12a, and Csm6. Science 2018, 360, 439–444. [Google Scholar] [CrossRef]
- Myhrvold, C.; Freije, C.A.; Gootenberg, J.S.; Abudayyeh, O.O.; Metsky, H.C.; Durbin, A.F.; Kellner, M.J.; Tan, A.L.; Paul, L.M.; Parham, L.A.; et al. Field-deployable viral diagnostics using CRISPR-Cas13. Science 2018, 360, 444–448. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | GC (%) | Amplification Length (bp) |
|---|---|---|---|
| PM-RPA-F-1 | GTTCCCACAACGATGAAGAACGCAGCGAAAT | 48.4 | 187 |
| PM-RPA-R-1 | AGTCAAAGCCAAAACAACCAGCAGCCAGCAC | 51.6 | |
| PM-RPA-F-2 | CCTCGTGCTGGCTGCTGGTTGTTTTGGCTTT | 54.8 | 235 |
| PM-RPA-R-2 | AGGCATAGAAGTGCTCGTTCCATCAGTTTCA | 45.2 | |
| PM-RPA-F-3 | AACGATGAAGAACGCAGCGAAATGCGATACGT | 46.9 | 190 |
| PM-RPA-R-3 | GCAATAGTGCCAGTCAAAGCCAAAACAACCAG | 46.9 | |
| PM-P-1 | [FAM] GTGCATAGACGTGGAAGGCTTGACCTGTCTAGTT [dSpacer]AAGACGGCGTTGACAC[C3-Spacer] | ||
| PM-P-2 | [FAM] GCCTGTCTCTGCTTAAGTTCTACTGTATAG [dSpacer]ACGTGCATAGACGTG[C3-Spacer] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, Y.; Luo, N.; Zong, Y.; Jia, M.; Rao, Y.; Huang, H.; Jiang, H. Recombinase Polymerase Amplification Combined with Lateral Flow Dipstick Assay for the Rapid and Sensitive Detection of Pseudo-nitzschia multiseries. Int. J. Mol. Sci. 2024, 25, 1350. https://doi.org/10.3390/ijms25021350
Yao Y, Luo N, Zong Y, Jia M, Rao Y, Huang H, Jiang H. Recombinase Polymerase Amplification Combined with Lateral Flow Dipstick Assay for the Rapid and Sensitive Detection of Pseudo-nitzschia multiseries. International Journal of Molecular Sciences. 2024; 25(2):1350. https://doi.org/10.3390/ijms25021350
Chicago/Turabian StyleYao, Yuqing, Ningjian Luo, Yujie Zong, Meng Jia, Yichen Rao, Hailong Huang, and Haibo Jiang. 2024. "Recombinase Polymerase Amplification Combined with Lateral Flow Dipstick Assay for the Rapid and Sensitive Detection of Pseudo-nitzschia multiseries" International Journal of Molecular Sciences 25, no. 2: 1350. https://doi.org/10.3390/ijms25021350
APA StyleYao, Y., Luo, N., Zong, Y., Jia, M., Rao, Y., Huang, H., & Jiang, H. (2024). Recombinase Polymerase Amplification Combined with Lateral Flow Dipstick Assay for the Rapid and Sensitive Detection of Pseudo-nitzschia multiseries. International Journal of Molecular Sciences, 25(2), 1350. https://doi.org/10.3390/ijms25021350