
The Role of PD-1/PD-L1 and IL-7 in Lymphocyte Dynamics and Sepsis Progression: A Biomarker Study in Critically Ill Patients
 ,
,  ,
,
Abstract
1. Introduction
2. Results
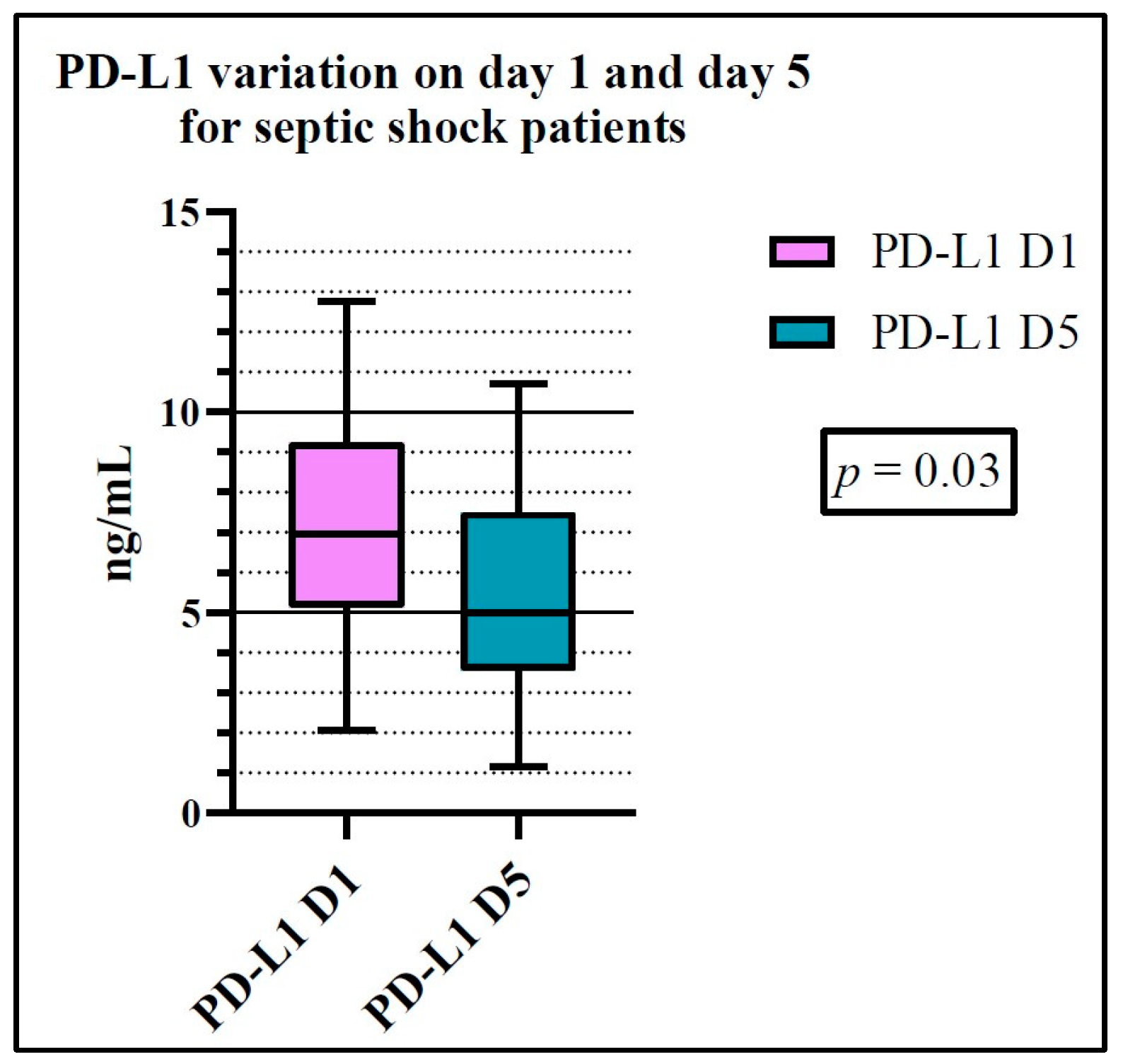
2.1. Population Analysis
2.2. Lymphopenia
2.3. Cutoff Value for IL-7
2.4. Comparison of Variables for the Sepsis and Septic Shock Groups
2.5. Comparison of Variables for the Survivor and Non-Survivor Groups
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Study Cohort
4.3. Evaluated Parameters
4.4. CBC and Lymphocyte Subsets
- CD4/CD8/CD3 (BDTritest, Cat. No. 342414) to identify and enumerate the following T-lymphocyte subset populations: CD3+ T lymphocytes, CD3+CD4+ helper/inducer T lymphocytes, and CD3+CD8+ suppressor/cytotoxic T lymphocytes.
- CD3/CD16+CD56/CD45/CD19 (BD Multitest, Cat. No. 342416) to identify and enumerate the following T-, B-, and NK-lymphocyte subset populations: CD3+ T lymphocytes, CD19+ B lymphocytes, and CD3-CD16+CD56+ NK lymphocytes.
4.5. PD-1/PD-L1
4.6. IL-7 Quantification
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The third international consensus definitions for sepsis and septic shock (SEpsis-3). JAMA 2016, 315, 801. [Google Scholar] [CrossRef] [PubMed]
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, regional, and national sepsis incidence and mortality, 1990–2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Pei, F.; Yao, R.-Q.; Ren, C.; Bahrami, S.; Billiar, T.R.; Chaudry, I.H.; Chen, D.-C.; Chen, X.-L.; Cui, N.; Fang, X.-M.; et al. Expert consensus on the monitoring and treatment of sepsis-induced immunosuppression. Mil. Med. Res. 2022, 9, 74. [Google Scholar] [CrossRef] [PubMed]
- Global Report on the Epidemiology and Burden of Sepsis: Current Evidence, Identifying Gaps and Future Directions; Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2020; ISBN 978-92-4-001078-9.
- Chen, J.; Wang, H.; Guo, R.; Li, H.; Cui, N. Early expression of functional markers on CD4+ T cells predicts outcomes in ICU patients with sepsis. Front. Immunol. 2022, 13, 938538. [Google Scholar] [CrossRef] [PubMed]
- Janssen, M.; Endeman, H.; Bos, L.D.J. Targeted immunomodulation: A primer for intensivists. Intensive Care Med. 2023, 49, 462–464. [Google Scholar] [CrossRef]
- Chen, R.; Qin, S.; Zhu, H.; Chang, G.; Li, M.; Lu, H.; Shen, M.; Gao, Q.; Lin, X. Dynamic monitoring of circulating CD8+ T and NK cell function in patients with septic shock. Immunol. Lett. 2022, 243, 61–68. [Google Scholar] [CrossRef]
- Doganyigit, Z.; Eroglu, E.; Akyuz, E. Inflammatory mediators of cytokines and chemokines in sepsis: From bench to bedside. Hum. Exp. Toxicol. 2022, 09603271221078871. [Google Scholar] [CrossRef]
- Leśnik, P.; Janc, J.; Mierzchala-Pasierb, M.; Tański, W.; Wierciński, J.; Łysenko, L. Interleukin-7 and interleukin-15 as prognostic biomarkers in sepsis and septic shock: Correlation with inflammatory markers and mortality. Cytokine 2023, 169, 156277. [Google Scholar] [CrossRef]
- Venet, F.; Demaret, J.; Blaise, B.J.; Rouget, C.; Girardot, T.; Idealisoa, E.; Rimmelé, T.; Mallet, F.; Lepape, A.; Textoris, J.; et al. IL-7 Restores T Lymphocyte Immunometabolic Failure in Septic Shock Patients through mTOR Activation. J. Immunol. 2017, 199, 1606–1615. [Google Scholar] [CrossRef]
- Ghosh, C.; Luong, G.; Sun, Y. A snapshot of the PD-1/PD-L1 pathway. J. Cancer 2021, 12, 2735–2746. [Google Scholar] [CrossRef]
- Qin, W.; Hu, L.; Zhang, X.; Jiang, S.; Li, J.; Zhang, Z.; Wang, X. The diverse function of PD-1/PD-L pathway beyond cancer. Front. Immunol. 2019, 10, 2298. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Yu-Jing, L.; Ma, T. Role of regulation of PD-1 and PD-L1 expression in sepsis. Front. Immunol. 2023, 14, 1029438. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Yin, Y. Regulatory role of the programmed cell death 1 signaling pathway in sepsis induced immunosuppression. Front. Immunol. 2023, 14, 1183542. [Google Scholar] [CrossRef] [PubMed]
- Rubio, I.; Osuchowski, M.F.; Shankar-Hari, M.; Skirecki, T.; Winkler, M.S.; Lachmann, G.; La Rosée, P.; Monneret, G.; Venet, F.; Bauer, M.; et al. Current gaps in sepsis immunology: New opportunities for translational research. Lancet Infect. Dis. 2019, 19, e422–e436. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.; Gerlach, H.; Vogelmann, T.; Preissing, F.; Stiefel, J.; Adam, D. Mortality in sepsis and septic shock in Europe, North America and Australia between 2009 and 2019—Results from a systematic review and meta-analysis. Crit. Care 2020, 24, 239. [Google Scholar] [CrossRef]
- Arefian, H.; Heublein, S.; Scherag, A.; Brunkhorst, F.M.; Younis, M.Z.; Moerer, O.; Fischer, D.; Hartmann, M. Hospital-related cost of sepsis: A systematic review. J. Infect. 2016, 74, 107–117. [Google Scholar] [CrossRef]
- Fleischmann-Struzek, C.; Mellhammar, L.; Rose, N.; Cassini, A.; Rudd, K.E.; Schlattmann, P.; Allegranzi, B.; Reinhart, K. Incidence and mortality of hospital- and ICU-treated sepsis: Results from an updated and expanded systematic review and meta-analysis. Intensive Care Med. 2020, 46, 1552–1562. [Google Scholar] [CrossRef]
- Pool, R.; Gomez, H.; Kellum, J.A. Mechanisms of organ dysfunction in sepsis. Crit. Care Clin. 2017, 34, 63–80. [Google Scholar] [CrossRef]
- Prest, J.; Nguyen, T.; Rajah, T.; Prest, A.B.; Sathananthan, M.; Jeganathan, N. Sepsis-Related mortality rates and trends based on site of infection. Crit. Care Explor. 2022, 4, e0775. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, J.; Xu, J.; Qiu, R.; Lin, T. Association between site of infection and mortality in patients with cancer with sepsis or septic shock: A retrospective cohort study. Exp. Ther. Med. 2022, 25, 33. [Google Scholar] [CrossRef]
- Van Vught, L.A.; Klouwenberg, P.M.C.K.; Spitoni, C.; Scicluna, B.P.; Wiewel, M.A.; Horn, J.; Schultz, M.J.; Nürnberg, P.; Bonten, M.J.M.; Cremer, O.L.; et al. Incidence, risk factors, and attributable mortality of secondary infections in the intensive care unit after admission for sepsis. JAMA 2016, 315, 1469. [Google Scholar] [CrossRef] [PubMed]
- Misra, A.K.; Levy, M.M.; Ward, N.S. Biomarkers of immunosuppression. Crit. Care Clin. 2019, 36, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Drewry, A.M.; Samra, N.; Skrupky, L.P.; Fuller, B.M.; Compton, S.M.; Hotchkiss, R.S. Persistent lymphopenia after diagnosis of sepsis predicts mortality. Shock 2014, 42, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Hohlstein, P.; Gussen, H.; Bartneck, M.; Warzecha, K.; Roderburg, C.; Buendgens, L.; Trautwein, C.; Koch, A.; Tacke, F. Prognostic Relevance of Altered Lymphocyte Subpopulations in Critical Illness and Sepsis. J. Clin. Med. 2019, 8, 353. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Du, H.; Su, Y.; Li, X.; Zhang, J.; Chen, M.; Ren, G.; He, F.; Niu, B. Nonviral infection-related lymphocytopenia for the prediction of adult sepsis and its persistence indicates a higher mortality. Medicine 2019, 98, e16535. [Google Scholar] [CrossRef]
- Huang, H.; Li, L.; Luo, W.; Yang, Y.; Ni, Y.; Song, T.; Zhu, Y.; Yang, Y.; Zhang, L. Lymphocyte percentage as a valuable predictor of prognosis in lung cancer. J. Cell. Mol. Med. 2022, 26, 1918–1931. [Google Scholar] [CrossRef]
- Daix, T.; Mathonnet, A.; Brakenridge, S.; Dequin, P.-F.; Mira, J.-P.; Berbille, F.; Morre, M.; Jeannet, R.; Blood, T.; Unsinger, J.; et al. Intravenously administered interleukin-7 to reverse lymphopenia in patients with septic shock: A double-blind, randomized, placebo-controlled trial. Ann. Intensive Care 2023, 13, 17. [Google Scholar] [CrossRef]
- Chen, D.; Tang, T.-X.; Deng, H.; Yang, X.-P.; Tang, Z.-H. Interleukin-7 Biology and its effects on immune cells: Mediator of generation, differentiation, survival, and homeostasis. Front. Immunol. 2021, 12, 747324. [Google Scholar] [CrossRef]
- De Roquetaillade, C.; Monneret, G.; Gossez, M.; Venet, F. IL-7 and its beneficial role in Sepsis-Induced T lymphocyte dysfunction. Crit. Rev. Immunol. 2018, 38, 433–451. [Google Scholar] [CrossRef]
- Bacârea, A.; Coman, O.; Bacârea, V.; Văsieșiu, A.; Săplăcan, I.; Fodor, R.; Grigorescu, B. Immune profile of patients-a new approach in management of sepsis and septic shock? Exp. Ther. Med. 2024, 27, 203. [Google Scholar] [CrossRef]
- Byun, H.K.; Chung, S.Y.; Kim, K.-J.; Seong, J. Role of Interleukin-7 in the Development of and Recovery from Radiation-Induced Lymphopenia: A Post-hoc Analysis of a Prospective Cohort. Cancer Res. Treat. 2021, 53, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Francois, B.; Jeannet, R.; Daix, T.; Walton, A.H.; Shotwell, M.S.; Unsinger, J.; Monneret, G.; Rimmelé, T.; Blood, T.; Morre, M.; et al. Interleukin-7 restores lymphocytes in septic shock: The IRIS-7 randomized clinical trial. JCI Insight 2018, 3, e98960. [Google Scholar] [CrossRef] [PubMed]
- Sermaxhaj, F.; Plavetić, N.D.; Gozalan, U.; Kulić, A.; Varga, L.R.; Popović, M.; Sović, S.; Mijatović, D.; Sermaxhaj, B.; Sopjani, M. The role of interleukin-7 serum level as biological marker in breast cancer: A cross-sectional, observational, and analytical study. World J. Surg. Oncol. 2022, 20, 225. [Google Scholar] [CrossRef] [PubMed]
- Ampuero, S.; Bahamonde, G.; Tempio, F.; Garmendia, M.L.; Ruiz, M.; Pizarro, R.; Rossi, P.; Huenchur, L.; Lizama, L.; López, M.; et al. IL-7/IL7R axis dysfunction in adults with severe community-acquired pneumonia (CAP): A cross-sectional study. Sci. Rep. 2022, 12, 13145. [Google Scholar] [CrossRef] [PubMed]
- Gunst, J.D.; Goonetilleke, N.; Rasmussen, T.A.; Søgaard, O.S. Immunomodulation with IL-7 and IL-15 in HIV-1 infection. J. Virus Erad. 2023, 9, 100347. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, Y.; Wang, J.-Y.; Wei, B.; Liu, Y.-G. Expression pattern of serum interleukin-7 in elderly septic patients and its prognostic value for predicting short-term mortality. Cytokine 2024, 180, 156664. [Google Scholar] [CrossRef]
- Martin, M.D.; Badovinac, V.P.; Griffith, T.S. CD4 T cell responses and the Sepsis-Induced Immunoparalysis state. Front. Immunol. 2020, 11, 1364. [Google Scholar] [CrossRef]
- Nedeva, C. Inflammation and Cell Death of the Innate and Adaptive Immune System during Sepsis. Biomolecules 2021, 11, 1011. [Google Scholar] [CrossRef]
- Wang, C.; Xu, H.; Gao, R.; Leng, F.; Huo, F.; Li, Y.; Liu, S.; Xu, M.; Bai, J. CD19+CD24hiCD38hi regulatory B cells deficiency revealed severity and poor prognosis in patients with sepsis. BMC Immunol. 2022, 23, 54. [Google Scholar] [CrossRef]
- Wu, H.; Tang, T.; Deng, H.; Chen, D.; Zhang, C.; Luo, J.; Chen, S.; Zhang, P.; Yang, J.; Dong, L. Immune checkpoint molecule Tim-3 promotes NKT cell apoptosis and predicts poorer prognosis in Sepsis. Clin. Immunol. 2023, 254, 109249. [Google Scholar] [CrossRef]
- Kim, E.Y.; Oldham, W.M. Innate T cells in the intensive care unit. Mol. Immunol. 2018, 105, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Young, J.S.; Monaghan, S.F.; Chung, C.S.; Cioffi, W.G.; Ayala, A.; Heffernan, D.S. Divergent Invariant Natural Killer T-Cell Response to Sepsis of Abdominal vs. Non-Abdominal Origin in Human Beings. Surg. Infect. 2015, 16, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Sari, M.I.; Ilyas, S. Correction: Sari, M.I.; Ilyas, S. The Expression Levels and Concentrations of PD-1 and PD-L1 proteins in septic patients: A Systematic review. Diagnostics 2022, 12, 2004, Erratum in Diagnostics 2023, 13, 1555. [Google Scholar] [CrossRef]
- Ruan, W.-S.; Feng, M.-X.; Xu, J.; Xu, Y.-G.; Song, C.-Y.; Lin, L.-Y.; Li, L.; Lu, Y.-Q. Early activation of Myeloid-Derived suppressor cells participate in Sepsis-Induced immune suppression via PD-L1/PD-1 axis. Front. Immunol. 2020, 11, 1299. [Google Scholar] [CrossRef]
- Sharpe, A.H.; Pauken, K.E. The diverse functions of the PD1 inhibitory pathway. Nat. Reviews. Immunol. 2017, 18, 153–167. [Google Scholar] [CrossRef]
- Konstantinidou, M.; Zarganes-Tzitzikas, T.; Magiera-Mularz, K.; Holak, T.A.; Dömling, A. Immune Checkpoint PD-1/PD-L1: Is there life beyond antibodies? Angew. Chem. Int. Ed. 2017, 57, 4840–4848. [Google Scholar] [CrossRef]
- Bakhshiani, Z.; Fouladi, S.; Mohammadzadeh, S.; Eskandari, N. Correlation of sPD1 with Procalcitonin and C-Reactive Protein Levels in Patients with Sepsis. Cell J. 2021, 23, 14–20. [Google Scholar] [CrossRef]
- Sun, S.; Chen, Y.; Liu, Z.; Tian, R.; Liu, J.; Chen, E.; Mao, E.; Pan, T.; Qu, H. Serum-soluble PD-L1 may be a potential diagnostic biomarker in sepsis. Scand. J. Immunol. 2021, 94, e13049. [Google Scholar] [CrossRef]
- Zhao, Y.; Jia, Y.; Li, C.; Shao, R.; Fang, Y. Predictive value of soluble programmed Death-1 for severe sepsis and septic shock during the first week in an intensive care unit. Shock 2018, 51, 289–297. [Google Scholar] [CrossRef]
- Shao, R.; Fang, Y.; Yu, H.; Zhao, L.; Jiang, Z.; Li, C.-S. Monocyte programmed death ligand-1 expression after 3–4 days of sepsis is associated with risk stratification and mortality in septic patients: A prospective cohort study. Crit. Care 2016, 20, 124. [Google Scholar] [CrossRef]
- Coman, O.; Grigorescu, B.-L.; Huțanu, A.; Bacârea, A.; Văsieșiu, A.M.; Fodor, R.Ș.; Stoica, F.; Azamfirei, L. The role of Programmed cell Death 1/Programmed Death ligand 1 (PD-1/PD-L1) axis in Sepsis-Induced Apoptosis. Medicina 2024, 60, 1174. [Google Scholar] [CrossRef] [PubMed]
- Torres, L.K.; Pickkers, P.; Van Der Poll, T. Sepsis-Induced immunosuppression. Annu. Rev. Physiol. 2021, 84, 157–181. [Google Scholar] [CrossRef] [PubMed]
- Nakamori, Y.; Park, E.J.; Shimaoka, M. Immune deregulation in sepsis and septic shock: Reversing immune paralysis by targeting PD-1/PD-L1 pathway. Front. Immunol. 2021, 11, 624279. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guo, D.-Z.; Zhu, C.-L.; Ren, S.-C.; Sun, C.-Y.; Wang, Y.; Wang, J.-F. The implication of targeting PD-1:PD-L1 pathway in treating sepsis through immunostimulatory and anti-inflammatory pathways. Front. Immunol. 2023, 14, 1323797. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Median (IQR) | p a Value | Normal Range | ||||
|---|---|---|---|---|---|---|
| Day 1 | Day 5 | % | ×103/μL | |||
| Sepsis | % | 0.067 (0.0589) | 0.085 (0.088) | 0.0032 | 0.205–0.45 | 1.2–3.4 |
| ×103/μL | 0.910 (0.405) | 1.02 (0.77) | 0.9323 | |||
| Septic shock | % | 0.0502 (0.0565) | 0.064 (0.0774) | 0.2387 | ||
| ×103/μL | 0.86 (0.6605) | 1.020 (0.8) | 0.1089 | |||
| Survivors | % | 0.0652 (0.058) | 0.116 (0.113) | 0.0014 | ||
| ×103/μL | 0.8985 (0.942) | 1.035 (1.718) | 0.6322 | |||
| Non-survivors | % | 0.057 (0.06) | 0.0635 (0.0537) | 0.1316 | ||
| ×103/μL | 0.91 (0.67) | 1.015 (0.7605) | 0.5014 | |||
| Median (IQR) (pg/mL) | IL-7 Cutoff Value (pg/mL) | ||
|---|---|---|---|
| Day 1 | Day 5 | ||
| Sepsis | 3.459 (4.204) | 2.382 (2.189) | 1.94 |
| Septic shock | 5.415 (5.35) | 3.459 (4.052) | |
| Survivors | 3.921 (4.346) | 3.227 (3.037) | |
| Non-survivors | 4.381 (5.834) | 2.542 (2.686) | |
| Tc Cells (CD8+), % | NKT (CD3+), % | B Cells (CD19+), % | ||
|---|---|---|---|---|
| Th cells (CD4+), % | Day 1 | r = −0.9368 (−0.9726 to −0.8575) p b < 0.0001 | r = −0.4108 (−0.7095 to 0.01305) p b = 0.0575 | r = 0.3740 (−0.04725 to 0.6823) p a = 0.0718 |
| Day 5 | r = −0.9788 (−0.9922 to −0.9426) p b < 0.0001 | r = −0.6998 (−0.8833 to −0.3302) p b = 0.0018 | r = 0.6929 (0.3004 to 0.8847) p b = 0.0029 | |
| Tc cells (CD8+), % | Day 1 | r = 0.5205 (0.1267 to 0.7726) p b = 0.0130 | r = −0.4517 (−0.7293 to −0.04645) p a = 0.0267 | |
| Day 5 | r = 0.6379 (0.2269 to 0.8561) p b = 0.0059 | r = −0.6794 (−0.8791 to −0.2770) p b = 0.0038 | ||
| NKT (CD3+), % | Day 1 | r = −0.4864 (−0.7592 to −0.06832) p a = 0.0217 | ||
| Day 5 | r = −0.4824 (−0.7976 to 0.03964) p b = 0.0686 |
| Tc Cells (CD8+), % | NKT (CD3+), % | B Cells (CD19+), % | ||
|---|---|---|---|---|
| Th cells (CD4+), % | Day 1 | r = −0.7114 (−0.8155 to −0.5629) p b < 0.0001 | r = −0.6415 (−0.7722 to −0.4588) p a < 0.0001 | r = 0.1206 (−0.1451 to 0.3700) p a = 0.3588 |
| Day 5 | r = −0.9461 (−0.9730 to −0.8939) p b < 0.0001 | r = −0.4962 (−0.7145 to −0.1899) p b = 0.0028 | r = 0.1412 (−0.2168 to 0.4657) p a = 0.4257 | |
| Tc cells (CD8+), % | Day 1 | r = 0.5488 (0.3379 to 0.7072) p a < 0.0001 | r = −0.1183 (−0.3680 to 0.1474) p a = 0.3682 | |
| Day 5 | r = 0.4641 (0.1493 to 0.6934) p b = 0.0057 | r = −0.08319 (−0.4184 to 0.2720) p a = 0.6400 | ||
| NKT (CD3+), % | Day 1 | r = −0.2894 (−0.5153 to −0.02578) p a = 0.0276 | ||
| Day 5 | r = −0.2814 (−0.5728 to 0.07306) p a = 0.1069 |
| Parameters (on Day 1) | Th Cells (CD4+), % | Tc Cells (CD8+), % | NKT (CD3+), % | B Cells (CD19+), % |
|---|---|---|---|---|
| PD-1, ng/mL | r = 0.3463 (0.08886 to 0.5603) p a = 0.0078 | r = −0.3873 (−0.5920 to −0.1357) p a = 0.0027 | r = −0.3663 (−0.5775 to −0.1091) p a = 0.0051 | r = 0.1884 (−0.08889 to 0.4386) p a = 0.1683 |
| PD-L1, ng/mL | r = 0.1843 (−0.08264 to 0.4265) p b = 0.1740 | r = −0.2790 (−0.5049 to −0.01736) p b = 0.0373 | r = 0.003428 (−0.2696 to 0.2759) p a = 0.9802 | r = −0.1443 (−0.4035 to 0.1364) p a = 0.2979 |
| IL-7, pg/mL | r = 0.1172 (−0.1484 to 0.3671) p a = 0.3723 | r = −0.1029 (−0.3545 to 0.1625) p a = 0.4339 | r = −0.05624 (−0.3171 to 0.2125) p a = 0.6750 | r = 0.1398 (−0.1307 to 0.3908) p a = 0.2954 |
| Parameter (on day 5) | Th cells (CD4+), % | Tc cells (CD8+), % | NKT (CD3+), % | B cells (CD19+), % |
| PD-1, ng/mL | r = −0.4123 (−0.6749 to −0.05702) p a = 0.0212 | r = 0.3414 (−0.02566 to 0.6273) p a = 0.0602 | r = 0.05467 (−0.3155 to 0.4104) p a = 0.7702 | r = −0.1472 (−0.4851 to 0.2289) p a = 0.4294 |
| PD-L1, ng/mL | r = −0.2116 (−0.5536 to 0.1915) p b = 0.2995 | r = 0.2440 (−0.1584 to 0.5768) p b = 0.2297 | r = −0.1061 (−0.4739 to 0.2934) p b = 0.6061 | r = −0.02838 (−0.4212 to 0.3734) p a = 0.8906 |
| IL-7, pg/mL | r = −0.1133 (−0.4757 to 0.2819) p a = 0.5659 | r = 0.1375 (−0.2592 to 0.4944) p a = 0.4855 | r = 0.1150 (−0.2803 to 0.4770) p a = 0.5600 | r = −0.1754 (−0.5233 to 0.2225) p a = 0.3719 |
| PD-1, ng/mL on day 1 | PD-1, ng/mL on day 5 | |||
| IL-7, pg/mL | r = 0.2317 (−0.04387 to 0.4744) p a = 0.0888 | r = 0.3774 (0.001264 to 0.6600) p a = 0.0436 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coman, O.; Grigorescu, B.-L.; Huțanu, A.; Bacârea, A.; Văsieșiu, A.M.; Fodor, R.Ș.; Petrișor, M.; Azamfirei, L. The Role of PD-1/PD-L1 and IL-7 in Lymphocyte Dynamics and Sepsis Progression: A Biomarker Study in Critically Ill Patients. Int. J. Mol. Sci. 2024, 25, 12612. https://doi.org/10.3390/ijms252312612
Coman O, Grigorescu B-L, Huțanu A, Bacârea A, Văsieșiu AM, Fodor RȘ, Petrișor M, Azamfirei L. The Role of PD-1/PD-L1 and IL-7 in Lymphocyte Dynamics and Sepsis Progression: A Biomarker Study in Critically Ill Patients. International Journal of Molecular Sciences. 2024; 25(23):12612. https://doi.org/10.3390/ijms252312612
Chicago/Turabian StyleComan, Oana, Bianca-Liana Grigorescu, Adina Huțanu, Anca Bacârea, Anca Meda Văsieșiu, Raluca Ștefania Fodor, Marius Petrișor, and Leonard Azamfirei. 2024. "The Role of PD-1/PD-L1 and IL-7 in Lymphocyte Dynamics and Sepsis Progression: A Biomarker Study in Critically Ill Patients" International Journal of Molecular Sciences 25, no. 23: 12612. https://doi.org/10.3390/ijms252312612
APA StyleComan, O., Grigorescu, B.-L., Huțanu, A., Bacârea, A., Văsieșiu, A. M., Fodor, R. Ș., Petrișor, M., & Azamfirei, L. (2024). The Role of PD-1/PD-L1 and IL-7 in Lymphocyte Dynamics and Sepsis Progression: A Biomarker Study in Critically Ill Patients. International Journal of Molecular Sciences, 25(23), 12612. https://doi.org/10.3390/ijms252312612