
Unveiling the Antiviral Properties of Panduratin A through SARS-CoV-2 Infection Modeling in Cardiomyocytes

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. iPSC-Derived CMs Express Cardiac Markers and Host Receptors for SARS-CoV-2
2.2. iPSC-Derived CMs Are Susceptible to SARS-CoV-2 Infection In Vitro
2.3. iPSC-Derived CMs-Based Platform Provides Accurate Screening for Cardiac Damage
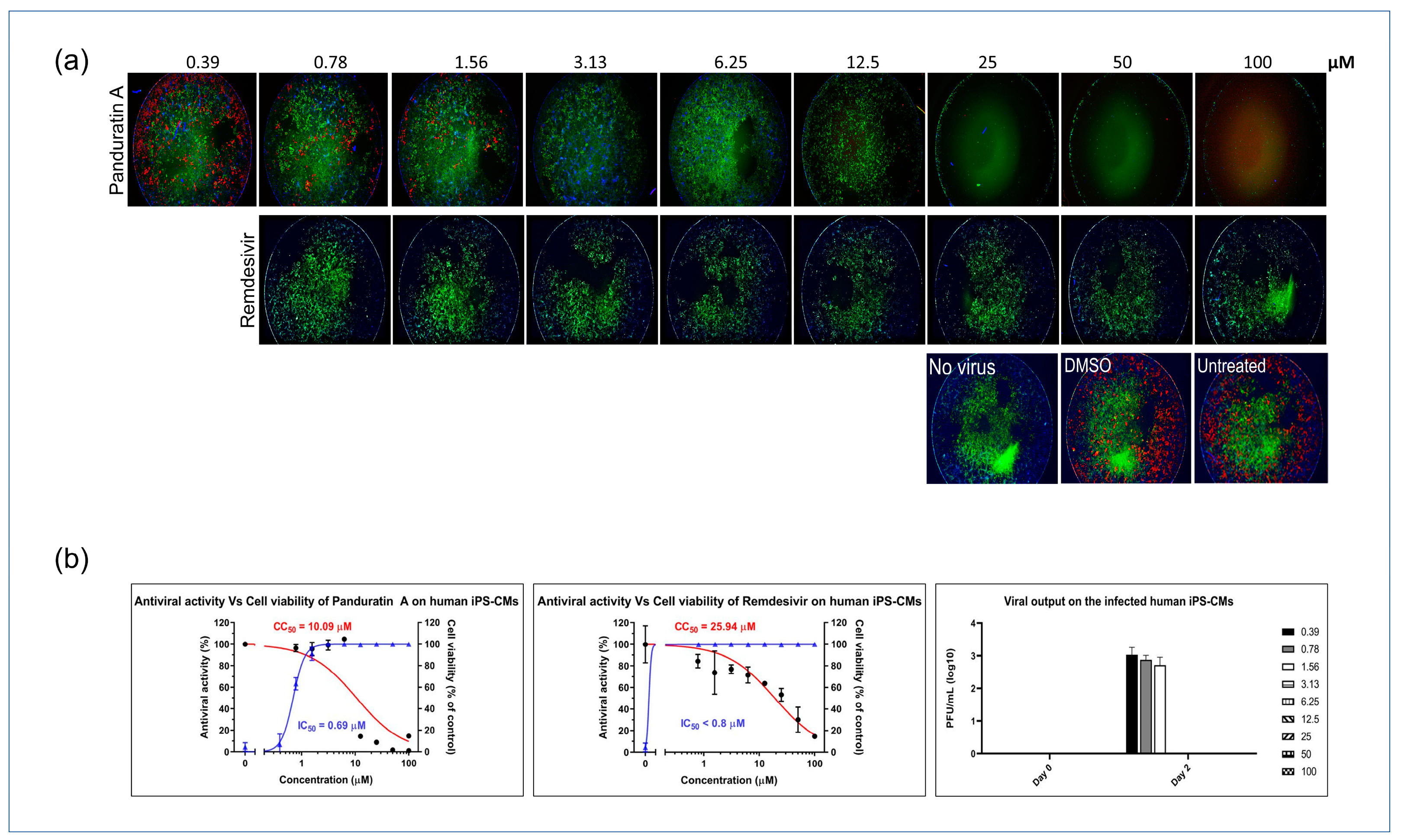
2.4. Panduratin A Potentially Inhibits SARS-CoV-2 Infection in iPSC-Derived CMs
3. Discussion
4. Materials and Methods
4.1. Compounds
4.2. iPSC Culture and CM Differentiation
4.3. SARS-CoV-2 Infection of Cultured Myocytes
4.4. Imaging and Immunofluorescence
4.5. Cytotoxicity Assays
4.6. Micro-Plaque Assay
4.7. Data and Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bekhite, M.M.; Schulze, P.C. Human Induced Pluripotent Stem Cell as a Disease Modeling and Drug Development Platform-A Cardiac Perspective. Cells 2021, 10, 3483. [Google Scholar] [CrossRef]
- Sharma, A.; McKeithan, W.L.; Serrano, R.; Kitani, T.; Burridge, P.W.; del Álamo, J.C.; Mercola, M.; Wu, J.C. Use of human induced pluripotent stem cell-derived cardiomyocytes to assess drug cardiotoxicity. Nat. Protoc. 2018, 13, 3018–3041. [Google Scholar] [CrossRef] [PubMed]
- McKeithan, W.L.; Savchenko, A.; Yu, M.S.; Cerignoli, F.; Bruyneel, A.A.N.; Price, J.H.; Mercola, M. An Automated Platform for Assessment of Congenital and Drug-Induced Arrhythmia with hiPSC-Derived Cardiomyocytes. Front. Physiol. 2017, 8, 766. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.P.; Rees, S.; Kalindjian, S.B.; Philpott, K.L. Principles of early drug discovery. Br. J. Pharmacol. 2011, 162, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Robinson, P.C.; Liew, D.F.L.; Tanner, H.L.; Grainger, J.R.; Dwek, R.A.; Reisler, R.B.; Steinman, L.; Feldmann, M.; Ho, L.-P.; Hussell, T.; et al. COVID-19 therapeutics: Challenges and directions for the future. Proc. Natl. Acad. Sci. USA 2022, 119, e2119893119. [Google Scholar] [CrossRef] [PubMed]
- Antonio, A.D.S.; Wiedemann, L.S.M.; Veiga-Junior, V.F. Natural products’ role against COVID-19. RSC Adv. 2020, 10, 23379–23393. [Google Scholar] [CrossRef]
- Shawky, E.; Nada, A.A.; Ibrahim, R.S. Potential role of medicinal plants and their constituents in the mitigation of SARS-CoV-2: Identifying related therapeutic targets using network pharmacology and molecular docking analyses. RSC Adv. 2020, 10, 27961–27983. [Google Scholar] [CrossRef]
- Kiat, T.S.; Pippen, R.; Yusof, R.; Ibrahim, H.; Khalid, N.; Rahman, N.A. Inhibitory activity of cyclohexenyl chalcone derivatives and flavonoids of fingerroot, Boesenbergia rotunda (L.), towards dengue-2 virus NS3 protease. Bioorg. Med. Chem. Lett. 2006, 16, 3337–3340. [Google Scholar] [CrossRef]
- Hanafy, N.A.N.; El-Kemary, M.A. Silymarin/curcumin loaded albumin nanoparticles coated by chitosan as muco-inhalable delivery system observing anti-inflammatory and anti COVID-19 characterizations in oleic acid triggered lung injury and in vitro COVID-19 experiment. Int. J. Biol. Macromol. 2022, 198, 101–110. [Google Scholar] [CrossRef]
- Su, H.; Yao, S.; Zhao, W.; Zhang, Y.; Liu, J.; Shao, Q.; Wang, Q.; Li, M.; Xie, H.; Shang, W.; et al. Identification of pyrogallol as a warhead in design of covalent inhibitors for the SARS-CoV-2 3CL protease. Nat. Commun. 2021, 12, 3623. [Google Scholar] [CrossRef] [PubMed]
- Trakoontivakorn, G.; Nakahara, K.; Shinmoto, H.; Takenaka, M.; Onishi-Kameyama, M.; Ono, H.; Yoshida, M.; Nagata, T.; Tsushida, T. Structural analysis of a novel antimutagenic compound, 4-Hydroxypanduratin A, and the antimutagenic activity of flavonoids in a Thai spice, fingerroot (Boesenbergia pandurata Schult.) against mutagenic heterocyclic amines. J. Agric. Food Chem. 2001, 49, 3046–3050. [Google Scholar] [CrossRef]
- Kanjanasirirat, P.; Suksatu, A.; Manopwisedjaroen, S.; Munyoo, B.; Tuchinda, P.; Jearawuttanakul, K.; Thitithanyanont, A. High-content screening of Thai medicinal plants reveals Boesenbergia rotunda extract and its component Panduratin A as anti-SARS-CoV-2 agents. Sci. Rep. 2020, 10, 19963. [Google Scholar] [CrossRef] [PubMed]
- Boonserm, P.; Khunrae, P.; Sutthibutpong, T. A computational study on the molecular mechanisms of panduratin A as a potential inhibitor on SARS-CoV-2 protein targets. Heliyon 2023, 9, e12780. [Google Scholar] [CrossRef]
- Vergoten, G.; Bailly, C. Interaction of panduratin A and derivatives with the SARS-CoV-2 main protease (m(pro)): A molecular docking study. J. Biomol. Struct. Dyn. 2023, 41, 6834–6844. [Google Scholar] [CrossRef] [PubMed]
- Kongratanapasert, T.; Kongsomros, S.; Arya, N.; Sutummaporn, K.; Wiriyarat, W.; Akkhawattanangkul, Y.; Boonyarattanasoonthorn, T.; Asavapanumas, N.; Kanjanasirirat, P.; Suksatu, A.; et al. Pharmacological Activities of Fingerroot Extract and Its Phytoconstituents against SARS-CoV-2 Infection in Golden Syrian Hamsters. J. Exp. Pharmacol. 2023, 15, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Tangprasittipap, A.; Jittorntrum, B.; Wongkummool, W.; Kitiyanant, N.; Tubsuwan, A. Generation of induced pluripotent stem cells from peripheral blood CD34+ hematopoietic progenitors of a 31year old healthy woman. Stem Cell Res. 2017, 20, 91–93. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef]
- Choi, S.W.; Shin, J.S.; Park, S.-J.; Jung, E.; Park, Y.-G.; Lee, J.; Kim, S.J.; Park, H.-J.; Lee, J.-H.; Park, S.-M.; et al. Antiviral activity and safety of remdesivir against SARS-CoV-2 infection in human pluripotent stem cell-derived cardiomyocytes. Antivir. Res. 2020, 184, 104955. [Google Scholar] [CrossRef]
- Li, S.-Y.; Chen, C.; Zhang, H.-Q.; Guo, H.-Y.; Wang, H.; Wang, L.; Zhang, X.; Hua, S.-N.; Yu, J.; Xiao, P.-G.; et al. Identification of natural compounds with antiviral activities against SARS-associated coronavirus. Antivir. Res. 2005, 67, 18–23. [Google Scholar] [CrossRef]
- Chen, L.; Li, J.; Luo, C.; Liu, H.; Xu, W.; Chen, G.; Liew, O.W.; Zhu, W.; Puah, C.M.; Shen, X.; et al. Binding interaction of quercetin-3-beta-galactoside and its synthetic derivatives with SARS-CoV 3CL(pro): Structure-activity relationship studies reveal salient pharmacophore features. Bioorg. Med. Chem. 2006, 14, 8295–8306. [Google Scholar] [CrossRef]
- Chen, G.; Gulbranson, D.R.; Hou, Z.; Bolin, J.M.; Ruotti, V.; Probasco, M.D.; Smuga-Otto, K.; Howden, S.E.; Diol, N.R.; Propson, N.E.; et al. Chemically defined conditions for human iPSC derivation and culture. Nat. Methods 2011, 8, 424–429. [Google Scholar] [CrossRef]
- Burridge, P.W.; Matsa, E.; Shukla, P.; Lin, Z.C.; Churko, J.M.; Ebert, A.D.; Lan, F.; Diecke, S.; Huber, B.; Mordwinkin, N.M.; et al. Chemically defined generation of human cardiomyocytes. Nat. Methods 2014, 11, 855–860. [Google Scholar] [CrossRef]
- Sharma, A.; Li, G.; Rajarajan, K.; Hamaguchi, R.; Burridge, P.W.; Wu, S.M. Derivation of highly purified cardiomyocytes from human induced pluripotent stem cells using small molecule-modulated differentiation and subsequent glucose starvation. J. Vis. Exp. 2015, 97, 52628. [Google Scholar]
- Mandal, S.; Gamit, N.; Biswas, S.; Rao, C.M.; Sethi, G.; Warrier, S. An efficient human stem cells derived cardiotoxicity testing platform for testing oncotherapeutic analogues of quercetin and cinnamic acid. Sci. Rep. 2022, 12, 21362. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linn, A.K.; Manopwisedjaroen, S.; Kanjanasirirat, P.; Borwornpinyo, S.; Hongeng, S.; Phanthong, P.; Thitithanyanont, A. Unveiling the Antiviral Properties of Panduratin A through SARS-CoV-2 Infection Modeling in Cardiomyocytes. Int. J. Mol. Sci. 2024, 25, 1427. https://doi.org/10.3390/ijms25031427
Linn AK, Manopwisedjaroen S, Kanjanasirirat P, Borwornpinyo S, Hongeng S, Phanthong P, Thitithanyanont A. Unveiling the Antiviral Properties of Panduratin A through SARS-CoV-2 Infection Modeling in Cardiomyocytes. International Journal of Molecular Sciences. 2024; 25(3):1427. https://doi.org/10.3390/ijms25031427
Chicago/Turabian StyleLinn, Aung Khine, Suwimon Manopwisedjaroen, Phongthon Kanjanasirirat, Suparerk Borwornpinyo, Suradej Hongeng, Phetcharat Phanthong, and Arunee Thitithanyanont. 2024. "Unveiling the Antiviral Properties of Panduratin A through SARS-CoV-2 Infection Modeling in Cardiomyocytes" International Journal of Molecular Sciences 25, no. 3: 1427. https://doi.org/10.3390/ijms25031427
APA StyleLinn, A. K., Manopwisedjaroen, S., Kanjanasirirat, P., Borwornpinyo, S., Hongeng, S., Phanthong, P., & Thitithanyanont, A. (2024). Unveiling the Antiviral Properties of Panduratin A through SARS-CoV-2 Infection Modeling in Cardiomyocytes. International Journal of Molecular Sciences, 25(3), 1427. https://doi.org/10.3390/ijms25031427