
In the Rat Hippocampus, Pilocarpine-Induced Status Epilepticus Is Associated with Reactive Glia and Concomitant Increased Expression of CD31, PDGFRβ, and Collagen IV in Endothelial Cells and Pericytes of the Blood–Brain Barrier


Abstract
:1. Introduction
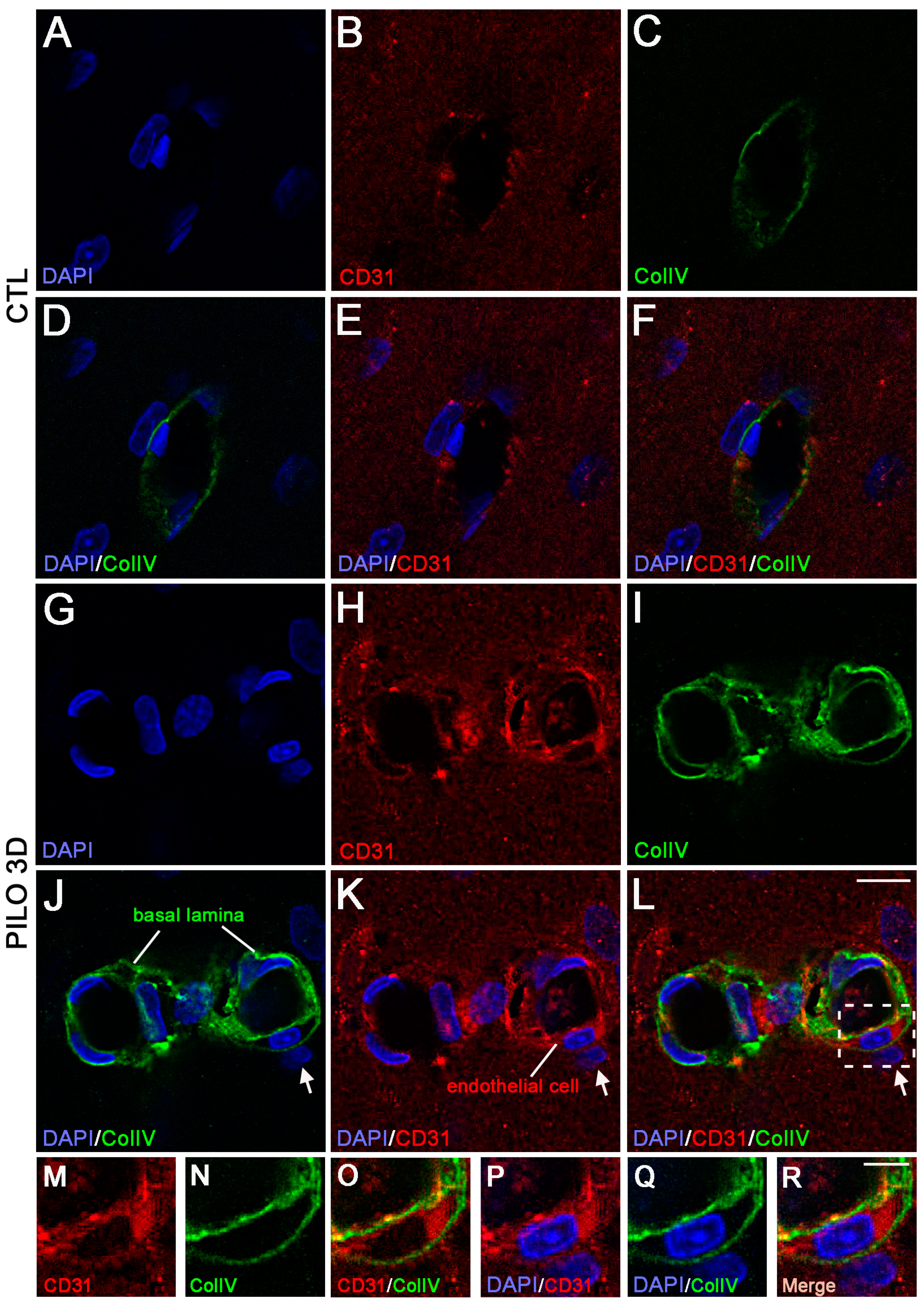
2. Results
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Rat Pilocarpine TLE Model
4.3. RNA Extraction and RT-qPCR
4.4. Tissue Preparation for Immunohistochemistry
4.5. Immunohistochemistry
4.6. Confocal Microscopy and Quantification
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baulac, M.; de Boer, H.; Elger, C.; Glynn, M.; Kälviäinen, R.; Little, A.; Mifsud, J.; Perucca, E.; Pitkänen, A.; Ryvlin, P. Epilepsy Priorities in Europe: A Report of the ILAE-IBE Epilepsy Advocacy Europe Task Force. Epilepsia 2015, 56, 1687–1695. [Google Scholar] [CrossRef] [PubMed]
- Ayalew, M.B.; Muche, E.A. Patient Reported Adverse Events among Epileptic Patients Taking Antiepileptic Drugs. SAGE Open Med. 2018, 6, 2050312118772471. [Google Scholar] [CrossRef]
- Ferhat, L. Potential Role of Drebrin a, an f-Actin Binding Protein, in Reactive Synaptic Plasticity after Pilocarpine-Induced Seizures: Functional Implications in Epilepsy. Int. J. Cell Biol. 2012, 2012, 474351. [Google Scholar] [CrossRef]
- Reddy, D.S.; Kuruba, R. Experimental Models of Status Epilepticus and Neuronal Injury for Evaluation of Therapeutic Interventions. Int. J. Mol. Sci. 2013, 14, 18284–18318. [Google Scholar] [CrossRef]
- Curia, G.; Lucchi, C.; Vinet, J.; Gualtieri, F.; Marinelli, C.; Torsello, A.; Costantino, L.; Biagini, G. Pathophysiogenesis of Mesial Temporal Lobe Epilepsy: Is Prevention of Damage Antiepileptogenic? Curr. Med. Chem. 2014, 21, 663–688. [Google Scholar] [CrossRef]
- Weissberg, I.; Reichert, A.; Heinemann, U.; Friedman, A. Blood-Brain Barrier Dysfunction in Epileptogenesis of the Temporal Lobe. Epilepsy Res Treat. 2011, 2011, 143908. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.J.; Patabendige, A.A.K.; Dolman, D.E.M.; Yusof, S.R.; Begley, D.J. Structure and Function of the Blood-Brain Barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Privratsky, J.R.; Newman, D.K.; Newman, P.J. PECAM-1: Conflicts of Interest in Inflammation. Life Sci. 2010, 87, 69–82. [Google Scholar] [CrossRef]
- Miettinen, M.; Lindenmayer, A.E.; Chaubal, A. Endothelial Cell Markers CD31, CD34, and BNH9 Antibody to H- and Y-Antigens--Evaluation of Their Specificity and Sensitivity in the Diagnosis of Vascular Tumors and Comparison with von Willebrand Factor. Mod. Pathol. 1994, 7, 82–90. [Google Scholar]
- Mbagwu, S.I.; Filgueira, L. Differential Expression of CD31 and Von Willebrand Factor on Endothelial Cells in Different Regions of the Human Brain: Potential Implications for Cerebral Malaria Pathogenesis. Brain Sci. 2020, 10, 31. [Google Scholar] [CrossRef]
- Yamanaka, G.; Takata, F.; Kataoka, Y.; Kanou, K.; Morichi, S.; Dohgu, S.; Kawashima, H. The Neuroinflammatory Role of Pericytes in Epilepsy. Biomedicines 2021, 9, 759. [Google Scholar] [CrossRef]
- Winkler, E.A.; Bell, R.D.; Zlokovic, B.V. Central Nervous System Pericytes in Health and Disease. Nat. Neurosci. 2011, 14, 1398–1405. [Google Scholar] [CrossRef]
- Armulik, A.; Genové, G.; Mäe, M.; Nisancioglu, M.H.; Wallgard, E.; Niaudet, C.; He, L.; Norlin, J.; Lindblom, P.; Strittmatter, K.; et al. Pericytes Regulate the Blood-Brain Barrier. Nature 2010, 468, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Dias, D.O.; Kim, H.; Holl, D.; Werne Solnestam, B.; Lundeberg, J.; Carlén, M.; Göritz, C.; Frisén, J. Reducing Pericyte-Derived Scarring Promotes Recovery after Spinal Cord Injury. Cell 2018, 173, 153–165.e22. [Google Scholar] [CrossRef]
- Klement, W.; Blaquiere, M.; Zub, E.; deBock, F.; Boux, F.; Barbier, E.; Audinat, E.; Lerner-Natoli, M.; Marchi, N. A Pericyte-Glia Scarring Develops at the Leaky Capillaries in the Hippocampus during Seizure Activity. Epilepsia 2019, 60, 1399–1411. [Google Scholar] [CrossRef]
- Xu, K.; Liu, F.; Xu, W.; Liu, J.; Chen, S.; Wu, G. Transplanting GABAergic Neurons Differentiated from Neural Stem Cells into Hippocampus Inhibits Seizures and Epileptiform Discharges in Pilocarpine-Induced Temporal Lobe Epilepsy Model. World Neurosurg. 2019, 128, e1–e11. [Google Scholar] [CrossRef]
- Klein, P.; Dingledine, R.; Aronica, E.; Bernard, C.; Blümcke, I.; Boison, D.; Brodie, M.J.; Brooks-Kayal, A.R.; Engel, J.; Forcelli, P.A.; et al. Commonalities in Epileptogenic Processes from Different Acute Brain Insults: Do They Translate? Epilepsia 2018, 59, 37–66. [Google Scholar] [CrossRef]
- van Vliet, E.A.; Otte, W.M.; Gorter, J.A.; Dijkhuizen, R.M.; Wadman, W.J. Longitudinal Assessment of Blood-Brain Barrier Leakage during Epileptogenesis in Rats. A Quantitative MRI Study. Neurobiol. Dis. 2014, 63, 74–84. [Google Scholar] [CrossRef]
- Gorter, J.A.; van Vliet, E.A.; Aronica, E. Status Epilepticus, Blood-Brain Barrier Disruption, Inflammation, and Epileptogenesis. Epilepsy Behav. 2015, 49, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Maroso, M.; Balosso, S.; Ravizza, T.; Liu, J.; Aronica, E.; Iyer, A.M.; Rossetti, C.; Molteni, M.; Casalgrandi, M.; Manfredi, A.A.; et al. Toll-like Receptor 4 and High-Mobility Group Box-1 Are Involved in Ictogenesis and Can Be Targeted to Reduce Seizures. Nat. Med. 2010, 16, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Liebner, S.; Dijkhuizen, R.M.; Reiss, Y.; Plate, K.H.; Agalliu, D.; Constantin, G. Functional Morphology of the Blood-Brain Barrier in Health and Disease. Acta Neuropathol. 2018, 135, 311–336. [Google Scholar] [CrossRef]
- van Vliet, E.A.; da Costa Araújo, S.; Redeker, S.; van Schaik, R.; Aronica, E.; Gorter, J.A. Blood-Brain Barrier Leakage May Lead to Progression of Temporal Lobe Epilepsy. Brain J. Neurol. 2007, 130, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Tomkins, O.; Shelef, I.; Kaizerman, I.; Eliushin, A.; Afawi, Z.; Misk, A.; Gidon, M.; Cohen, A.; Zumsteg, D.; Friedman, A. Blood-Brain Barrier Disruption in Post-Traumatic Epilepsy. J. Neurol. Neurosurg. Psychiatry 2008, 79, 774–777. [Google Scholar] [CrossRef] [PubMed]
- Raabe, A.; Schmitz, A.K.; Pernhorst, K.; Grote, A.; von der Brelie, C.; Urbach, H.; Friedman, A.; Becker, A.J.; Elger, C.E.; Niehusmann, P. Cliniconeuropathologic Correlations Show Astroglial Albumin Storage as a Common Factor in Epileptogenic Vascular Lesions. Epilepsia 2012, 53, 539–548. [Google Scholar] [CrossRef]
- Patel, D.C.; Tewari, B.P.; Chaunsali, L.; Sontheimer, H. Neuron-Glia Interactions in the Pathophysiology of Epilepsy. Nat. Rev. Neurosci. 2019, 20, 282–297. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Pristov, J.B.; Nobili, P.; Nikolić, L. Can Glial Cells Save Neurons in Epilepsy? Neural Regen. Res. 2023, 18, 1417–1422. [Google Scholar] [CrossRef] [PubMed]
- Kyriatzis, G.; Bernard, A.; Bôle, A.; Pflieger, G.; Chalas, P.; Masse, M.; Lécorché, P.; Jacquot, G.; Ferhat, L.; Khrestchatisky, M. Neurotensin Receptor 2 Is Induced in Astrocytes and Brain Endothelial Cells in Relation to Neuroinflammation Following Pilocarpine-Induced Seizures in Rats. Glia 2021, 69, 2618–2643. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Kastner, R.; Ingvar, M. Loss of Immunoreactivity for Glial Fibrillary Acidic Protein (GFAP) in Astrocytes as a Marker for Profound Tissue Damage in Substantia Nigra and Basal Cortical Areas after Status Epilepticus Induced by Pilocarpine in Rat. Glia 1994, 12, 165–172. [Google Scholar] [CrossRef]
- Sharma, A.K.; Reams, R.Y.; Jordan, W.H.; Miller, M.A.; Thacker, H.L.; Snyder, P.W. Mesial Temporal Lobe Epilepsy: Pathogenesis, Induced Rodent Models and Lesions. Toxicol. Pathol. 2007, 35, 984–999. [Google Scholar] [CrossRef]
- Knake, S.; Hamer, H.M.; Rosenow, F. Status Epilepticus: A Critical Review. Epilepsy Behav. 2009, 15, 10–14. [Google Scholar] [CrossRef]
- Papageorgiou, I.E.; Gabriel, S.; Fetani, A.F.; Kann, O.; Heinemann, U. Redistribution of Astrocytic Glutamine Synthetase in the Hippocampus of Chronic Epileptic Rats. Glia 2011, 59, 1706–1718. [Google Scholar] [CrossRef]
- Uzunalli, G.; Dieterly, A.M.; Kemet, C.M.; Weng, H.-Y.; Soepriatna, A.H.; Goergen, C.J.; Shinde, A.B.; Wendt, M.K.; Lyle, L.T. Dynamic Transition of the Blood-Brain Barrier in the Development of Non-Small Cell Lung Cancer Brain Metastases. Oncotarget 2019, 10, 6334–6348. [Google Scholar] [CrossRef] [PubMed]
- Mendes, N.F.; Pansani, A.P.; Carmanhães, E.R.F.; Tange, P.; Meireles, J.V.; Ochikubo, M.; Chagas, J.R.; da Silva, A.V.; Monteiro de Castro, G.; Le Sueur-Maluf, L. The Blood-Brain Barrier Breakdown During Acute Phase of the Pilocarpine Model of Epilepsy Is Dynamic and Time-Dependent. Front. Neurol. 2019, 10, 382. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Zhou, Y.; Chen, M.; Li, L.; Zhang, W. Pericytes for Therapeutic Approaches to Ischemic Stroke. Front. Neurosci. 2021, 15, 629297. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Kastner, R.; Ingvar, M. Laminar Damage of Neurons and Astrocytes in Neocortex and Hippocampus of Rat after Long-Lasting Status Epilepticus Induced by Pilocarpine. Epilepsy Res. Suppl. 1996, 12, 309–316. [Google Scholar] [PubMed]
- Massey, C.A.; Sowers, L.P.; Dlouhy, B.J.; Richerson, G.B. Mechanisms of Sudden Unexpected Death in Epilepsy: The Pathway to Prevention. Nat. Rev. Neurol. 2014, 10, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Teran, F.A.; Bravo, E.; Richerson, G.B. Chapter 8-Sudden Unexpected Death in Epilepsy: Respiratory Mechanisms. In Handbook of Clinical Neurology; Chen, R., Guyenet, P.G., Eds.; Respiratory Neurobiology: Physiology and Clinical Disorders, Part II.; Elsevier: Amsterdam, The Netherlands, 2022; Volume 189, pp. 153–176. [Google Scholar] [CrossRef]
- Olesen, J.B.; Abildstrøm, S.Z.; Erdal, J.; Gislason, G.H.; Weeke, P.; Andersson, C.; Torp-Pedersen, C.; Hansen, P.R. Effects of Epilepsy and Selected Antiepileptic Drugs on Risk of Myocardial Infarction, Stroke, and Death in Patients with or without Previous Stroke: A Nationwide Cohort Study. Pharmacoepidemiol. Drug Saf. 2011, 20, 964–971. [Google Scholar] [CrossRef] [PubMed]
- Boyd, E.M.; Fulford, R.A. Pilocarpine-Induced Convulsions and Delayed Psychotic-like Reaction. Can. J. Biochem. Physiol. 1961, 39, 1287–1294. [Google Scholar] [CrossRef]
- Kouchi, H.; Ogier, M.; Dieuset, G.; Morales, A.; Georges, B.; Rouanet, J.-L.; Martin, B.; Ryvlin, P.; Rheims, S.; Bezin, L. Respiratory Dysfunction in Two Rodent Models of Chronic Epilepsy and Acute Seizures and Its Link with the Brainstem Serotonin System. Sci. Rep. 2022, 12, 10248. [Google Scholar] [CrossRef]
- Sbai, O.; Soussi, R.; Bole, A.; Khrestchatisky, M.; Esclapez, M.; Ferhat, L. The Actin Binding Protein α-Actinin-2 Expression Is Associated with Dendritic Spine Plasticity and Migrating Granule Cells in the Rat Dentate Gyrus Following Pilocarpine-Induced Seizures. Exp. Neurol. 2021, 335, 113512. [Google Scholar] [CrossRef]
- Esclapez, M.; Hirsch, J.C.; Ben-Ari, Y.; Bernard, C. Newly Formed Excitatory Pathways Provide a Substrate for Hyperexcitability in Experimental Temporal Lobe Epilepsy. J. Comp. Neurol. 1999, 408, 449–460. [Google Scholar] [CrossRef]
- Goffin, K.; Nissinen, J.; Van Laere, K.; Pitkänen, A. Cyclicity of Spontaneous Recurrent Seizures in Pilocarpine Model of Temporal Lobe Epilepsy in Rat. Exp. Neurol. 2007, 205, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Garzillo, C.; Mello, L. Characterization of Reactive Astrocytes in the Chronic Phase of the Pilocarpine Model of Epilepsy. Epilepsia 2002, 43, 107–109. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Koh, S. Role of Brain Inflammation in Epileptogenesis. Yonsei Med. J. 2008, 49, 1–18. [Google Scholar] [CrossRef]
- Shapiro, L.A.; Wang, L.; Ribak, C.E. Rapid Astrocyte and Microglial Activation Following Pilocarpine-Induced Seizures in Rats. Epilepsia 2008, 49 (Suppl. S2), 33–41. [Google Scholar] [CrossRef] [PubMed]
- Lertkiatmongkol, P.; Liao, D.; Mei, H.; Hu, Y.; Newman, P.J. Endothelial Functions of Platelet/Endothelial Cell Adhesion Molecule-1 (CD31). Curr. Opin. Hematol. 2016, 23, 253–259. [Google Scholar] [CrossRef]
- Yan, B.C.; Xu, P.; Gao, M.; Wang, J.; Jiang, D.; Zhu, X.; Won, M.-H.; Su, P.Q. Changes in the Blood-Brain Barrier Function Are Associated With Hippocampal Neuron Death in a Kainic Acid Mouse Model of Epilepsy. Front. Neurol. 2018, 9, 775. [Google Scholar] [CrossRef]
- Rana, A.; Musto, A.E. The Role of Inflammation in the Development of Epilepsy. J. Neuroinflammation 2018, 15, 144. [Google Scholar] [CrossRef]
- Williams, K.C.; Zhao, R.W.; Ueno, K.; Hickey, W.F. PECAM-1 (CD31) Expression in the Central Nervous System and Its Role in Experimental Allergic Encephalomyelitis in the Rat. J. Neurosci. Res. 1996, 45, 747–757. [Google Scholar] [CrossRef]
- Cheung, K.; Ma, L.; Wang, G.; Coe, D.; Ferro, R.; Falasca, M.; Buckley, C.D.; Mauro, C.; Marelli-Berg, F.M. CD31 Signals Confer Immune Privilege to the Vascular Endothelium. Proc. Natl. Acad. Sci. USA 2015, 112, E5815–E5824. [Google Scholar] [CrossRef]
- Shen, J.; Ishii, Y.; Xu, G.; Dang, T.C.; Hamashima, T.; Matsushima, T.; Yamamoto, S.; Hattori, Y.; Takatsuru, Y.; Nabekura, J.; et al. PDGFR-β as a Positive Regulator of Tissue Repair in a Mouse Model of Focal Cerebral Ischemia. J. Cereb. Blood Flow Metab. 2012, 32, 353–367. [Google Scholar] [CrossRef]
- Iihara, K.; Sasahara, M.; Hashimoto, N.; Hazama, F. Induction of Platelet-Derived Growth Factor Beta-Receptor in Focal Ischemia of Rat Brain. J. Cereb. Blood Flow Metab. 1996, 16, 941–949. [Google Scholar] [CrossRef]
- Krupinski, J.; Issa, R.; Bujny, T.; Slevin, M.; Kumar, P.; Kumar, S.; Kaluza, J. A Putative Role for Platelet-Derived Growth Factor in Angiogenesis and Neuroprotection after Ischemic Stroke in Humans. Stroke 1997, 28, 564–573. [Google Scholar] [CrossRef]
- Arimura, K.; Ago, T.; Kamouchi, M.; Nakamura, K.; Ishitsuka, K.; Kuroda, J.; Sugimori, H.; Ooboshi, H.; Sasaki, T.; Kitazono, T. PDGF Receptor β Signaling in Pericytes Following Ischemic Brain Injury. Curr. Neurovasc. Res. 2012, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chintalgattu, V.; Ai, D.; Langley, R.R.; Zhang, J.; Bankson, J.A.; Shih, T.L.; Reddy, A.K.; Coombes, K.R.; Daher, I.N.; Pati, S.; et al. Cardiomyocyte PDGFR-Beta Signaling Is an Essential Component of the Mouse Cardiac Response to Load-Induced Stress. J. Clin. Investig. 2010, 120, 472–484. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Takata, F.; Yamanaka, G.; Yasunaga, M.; Hashiguchi, K.; Tominaga, K.; Itoh, K.; Kataoka, Y.; Yamauchi, A.; Dohgu, S. Reactive Pericytes in Early Phase Are Involved in Glial Activation and Late-Onset Hypersusceptibility to Pilocarpine-Induced Seizures in Traumatic Brain Injury Model Mice. J. Pharmacol. Sci. 2021, 145, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Arango-Lievano, M.; Boussadia, B.; De Terdonck, L.D.T.; Gault, C.; Fontanaud, P.; Lafont, C.; Mollard, P.; Marchi, N.; Jeanneteau, F. Topographic Reorganization of Cerebrovascular Mural Cells under Seizure Conditions. Cell Rep. 2018, 23, 1045–1059. [Google Scholar] [CrossRef] [PubMed]
- Garbelli, R.; de Bock, F.; Medici, V.; Rousset, M.C.; Villani, F.; Boussadia, B.; Arango-Lievano, M.; Jeanneteau, F.; Daneman, R.; Bartolomei, F.; et al. PDGFRβ(+) Cells in Human and Experimental Neuro-Vascular Dysplasia and Seizures. Neuroscience 2015, 306, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Milesi, S.; Boussadia, B.; Plaud, C.; Catteau, M.; Rousset, M.-C.; De Bock, F.; Scheffer, M.; Lerner-Natoli, M.; Rigau, V.; Marchi, N. Redistribution of PDGFRβ Cells and NG2DsRed Pericytes at the Cerebrovasculature after Status Epilepticus. Neurobiol. Dis. 2014, 71, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Klement, W.; Garbelli, R.; Zub, E.; Rossini, L.; Tassi, L.; Girard, B.; Blaquiere, M.; Bertaso, F.; Perroy, J.; de Bock, F.; et al. Seizure Progression and Inflammatory Mediators Promote Pericytosis and Pericyte-Microglia Clustering at the Cerebrovasculature. Neurobiol. Dis. 2018, 113, 70–81. [Google Scholar] [CrossRef]
- Michalski, D.; Spielvogel, E.; Puchta, J.; Reimann, W.; Barthel, H.; Nitzsche, B.; Mages, B.; Jäger, C.; Martens, H.; Horn, A.K.E.; et al. Increased Immunosignals of Collagen IV and Fibronectin Indicate Ischemic Consequences for the Neurovascular Matrix Adhesion Zone in Various Animal Models and Human Stroke Tissue. Front. Physiol. 2020, 11, 575598. [Google Scholar] [CrossRef]
- Christensen, N.M.; Trevisan, C.; Leifsson, P.S.; Johansen, M.V. The Association between Seizures and Deposition of Collagen in the Brain in Porcine Taenia Solium Neurocysticercosis. Vet. Parasitol. 2016, 228, 180–182. [Google Scholar] [CrossRef]
- Liesi, P.; Kauppila, T. Induction of Type IV Collagen and Other Basement-Membrane-Associated Proteins after Spinal Cord Injury of the Adult Rat May Participate in Formation of the Glial Scar. Exp. Neurol. 2002, 173, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Özkan, E.; Çetin-Taş, Y.; Şekerdağ, E.; Kızılırmak, A.B.; Taş, A.; Yıldız, E.; Yapıcı-Eser, H.; Karahüseyinoğlu, S.; Zeybel, M.; Gürsoy-Özdemir, Y. Blood-Brain Barrier Leakage and Perivascular Collagen Accumulation Precede Microvessel Rarefaction and Memory Impairment in a Chronic Hypertension Animal Model. Metab. Brain Dis. 2021, 36, 2553–2566. [Google Scholar] [CrossRef] [PubMed]
- Veznedaroglu, E.; Van Bockstaele, E.J.; O’Connor, M.J. Extravascular Collagen in the Human Epileptic Brain: A Potential Substrate for Aberrant Cell Migration in Cases of Temporal Lobe Epilepsy. J. Neurosurg. 2002, 97, 1125–1130. [Google Scholar] [CrossRef]
- Lowenstein, D.H.; Arsenault, L. Dentate Granule Cell Layer Collagen Explant Cultures: Spontaneous Axonal Growth and Induction by Brain-Derived Neurotrophic Factor or Basic Fibroblast Growth Factor. Neuroscience 1996, 74, 1197–1208. [Google Scholar] [CrossRef]
- Hoeppner, T.J.; Morrell, F. Control of Scar Formation in Experimentally Induced Epilepsy. Exp. Neurol. 1986, 94, 519–536. [Google Scholar] [CrossRef] [PubMed]
- Racine, R.J. Modification of Seizure Activity by Electrical Stimulation. II. Motor Seizure. Electroencephalogr. Clin. Neurophysiol. 1972, 32, 281–294. [Google Scholar] [CrossRef]
- Soussi, R.; Boulland, J.-L.; Bassot, E.; Bras, H.; Coulon, P.; Chaudhry, F.A.; Storm-Mathisen, J.; Ferhat, L.; Esclapez, M. Reorganization of Supramammillary-Hippocampal Pathways in the Rat Pilocarpine Model of Temporal Lobe Epilepsy: Evidence for Axon Terminal Sprouting. Brain Struct. Funct. 2015, 220, 2449–2468. [Google Scholar] [CrossRef] [PubMed]
- Bajorat, R.; Wilde, M.; Sellmann, T.; Kirschstein, T.; Köhling, R. Seizure Frequency in Pilocarpine-Treated Rats Is Independent of Circadian Rhythm. Epilepsia 2011, 52, e118–e122. [Google Scholar] [CrossRef]
- Meyerholz, D.K.; Beck, A.P. Principles and approaches for reproducible scoring of tissue stains in research. Lab. Invest. 2018, 98, 844–855. [Google Scholar] [CrossRef]
- Kim, S.W.; Roh, J.; Park, C.S. Immunohistochemistry for Pathologists: Protocols, Pitfalls, and Tips. J. Pathol. Transl. Med. 2016, 50, 411–418. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 4th ed.; Academic Press, Inc.: San Diego, CA, USA, 1998; ISBN 0125476183. [Google Scholar]
- Sbai, O.; Ould-Yahoui, A.; Ferhat, L.; Gueye, Y.; Bernard, A.; Charrat, E.; Mehanna, A.; Risso, J.-J.; Chauvin, J.-P.; Fenouillet, E.; et al. Differential Vesicular Distribution and Trafficking of MMP-2, MMP-9, and Their Inhibitors in Astrocytes. Glia 2010, 58, 344–366. [Google Scholar] [CrossRef]
- Pagano, A.; Breuzard, G.; Parat, F.; Tchoghandjian, A.; Figarella-Branger, D.; De Bessa, T.C.; Garrouste, F.; Douence, A.; Barbier, P.; Kovacic, H. Tau Regulates Glioblastoma Progression, 3D Cell Organization, Growth and Migration via the PI3K-AKT Axis. Cancers 2021, 13, 5818. [Google Scholar] [CrossRef]
- Silvani, G.; Romanov, V.; Cox, C.D.; Martinac, B. Biomechanical Characterization of Endothelial Cells Exposed to Shear Stress Using Acoustic Force Spectroscopy. Front. Bioeng. Biotechnol. 2021, 9, 612151. [Google Scholar] [CrossRef]
- Dell, R.B.; Holleran, S.; Ramakrishnan, R. Sample Size Determination. ILAR J. 2002, 43, 207–213. [Google Scholar] [CrossRef]
- Festing, M.F.W.; Altman, D.G. Guidelines for the Design and Statistical Analysis of Experiments Using Laboratory Animals. ILAR J. 2002, 43, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Haq, S.; Sarodaya, N.; Karapurkar, J.K.; Suresh, B.; Jo, J.K.; Singh, V.; Bae, Y.S.; Kim, K.-S.; Ramakrishna, S. CYLD Destabilizes NoxO1 Protein by Promoting Ubiquitination and Regulates Prostate Cancer Progression. Cancer Lett. 2022, 525, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Sharvit, E.; Abramovitch, S.; Reif, S.; Bruck, R. Amplified Inhibition of Stellate Cell Activation Pathways by PPAR-γ, RAR and RXR Agonists. PLoS ONE 2013, 8, e76541. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Description | Probe ID |
|---|---|---|
| Gfap | Glial fibrillary acid protein | Rn01253033 |
| Iba1 | Ionized calcium binding adaptor molecule1 | Rn00574125 |
| CD31 or PECAM-1 | EndoCAM or Platelet endothelial cell adhesion molecule-1 | Rn01467262 |
| PDGFRβ | Platelet-derived growth factor beta | Rn01502596 |
| ColIV a1 | Collagen, type IV, a1 | Rn01482927 |
| ColIV a3 | Collagen, type IV, a3 | Rn01400991 |
| RPL13 | Ribosomal Protein L13 | Rn00821258 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyriatzis, G.; Bernard, A.; Bôle, A.; Khrestchatisky, M.; Ferhat, L. In the Rat Hippocampus, Pilocarpine-Induced Status Epilepticus Is Associated with Reactive Glia and Concomitant Increased Expression of CD31, PDGFRβ, and Collagen IV in Endothelial Cells and Pericytes of the Blood–Brain Barrier. Int. J. Mol. Sci. 2024, 25, 1693. https://doi.org/10.3390/ijms25031693
Kyriatzis G, Bernard A, Bôle A, Khrestchatisky M, Ferhat L. In the Rat Hippocampus, Pilocarpine-Induced Status Epilepticus Is Associated with Reactive Glia and Concomitant Increased Expression of CD31, PDGFRβ, and Collagen IV in Endothelial Cells and Pericytes of the Blood–Brain Barrier. International Journal of Molecular Sciences. 2024; 25(3):1693. https://doi.org/10.3390/ijms25031693
Chicago/Turabian StyleKyriatzis, Grigorios, Anne Bernard, Angélique Bôle, Michel Khrestchatisky, and Lotfi Ferhat. 2024. "In the Rat Hippocampus, Pilocarpine-Induced Status Epilepticus Is Associated with Reactive Glia and Concomitant Increased Expression of CD31, PDGFRβ, and Collagen IV in Endothelial Cells and Pericytes of the Blood–Brain Barrier" International Journal of Molecular Sciences 25, no. 3: 1693. https://doi.org/10.3390/ijms25031693
APA StyleKyriatzis, G., Bernard, A., Bôle, A., Khrestchatisky, M., & Ferhat, L. (2024). In the Rat Hippocampus, Pilocarpine-Induced Status Epilepticus Is Associated with Reactive Glia and Concomitant Increased Expression of CD31, PDGFRβ, and Collagen IV in Endothelial Cells and Pericytes of the Blood–Brain Barrier. International Journal of Molecular Sciences, 25(3), 1693. https://doi.org/10.3390/ijms25031693