
Plant miR8126-3p and miR8126-5p Decrease Lipid Accumulation through Modulation of Metabolic Genes in a Human Hepatocyte Model That Mimics Steatosis
 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Identification of Human Putative Target Genes of Plant miR8126-3p and miR8126-5p
2.2. Plant miR8126-3p and miR8126-5p Were Detected in HepG2 Cells after Transfection
2.3. Plant miR8126-3p and miR8126-5p Transfections Did Not Induce Cytotoxicity in HepG2 Cells
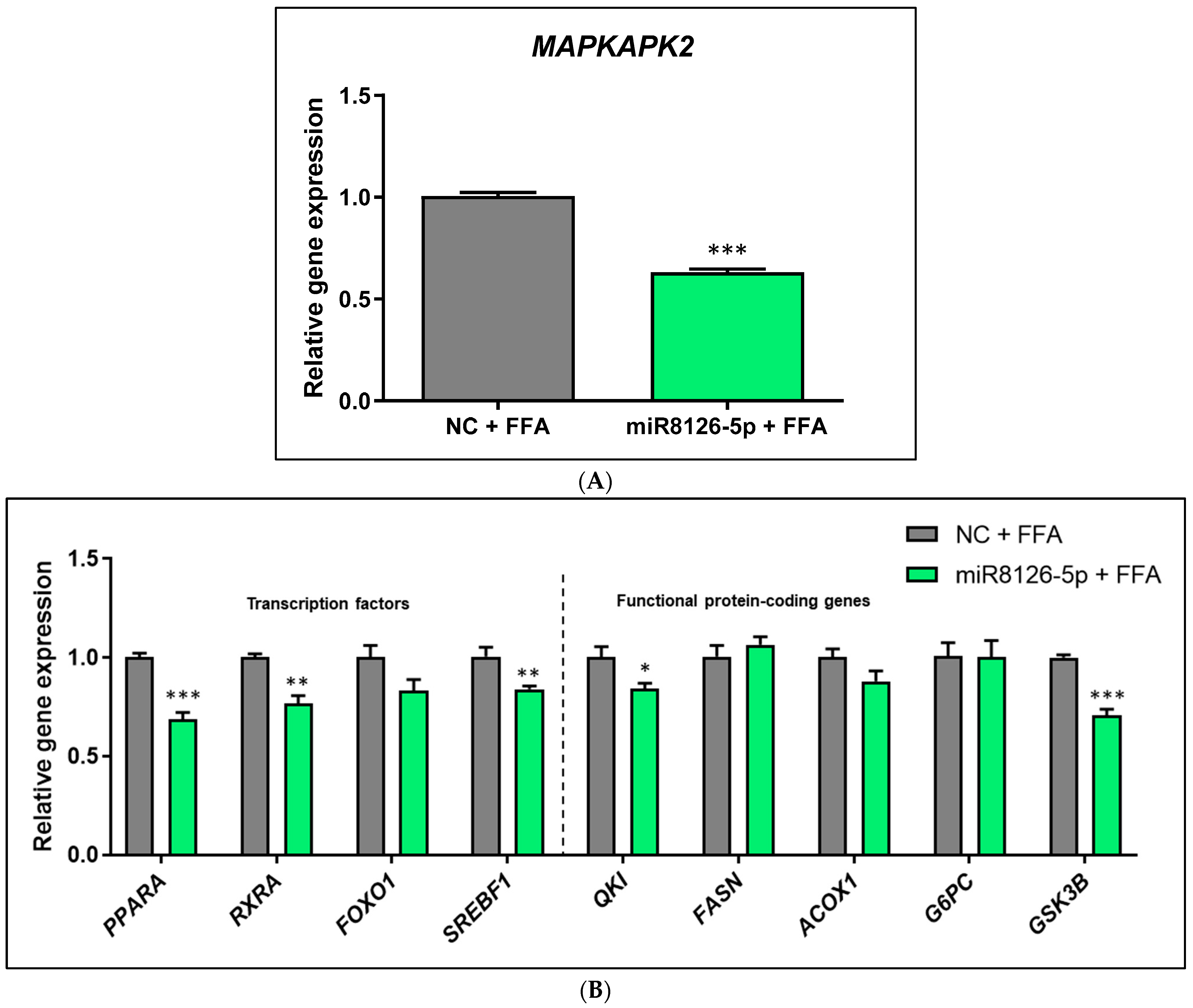
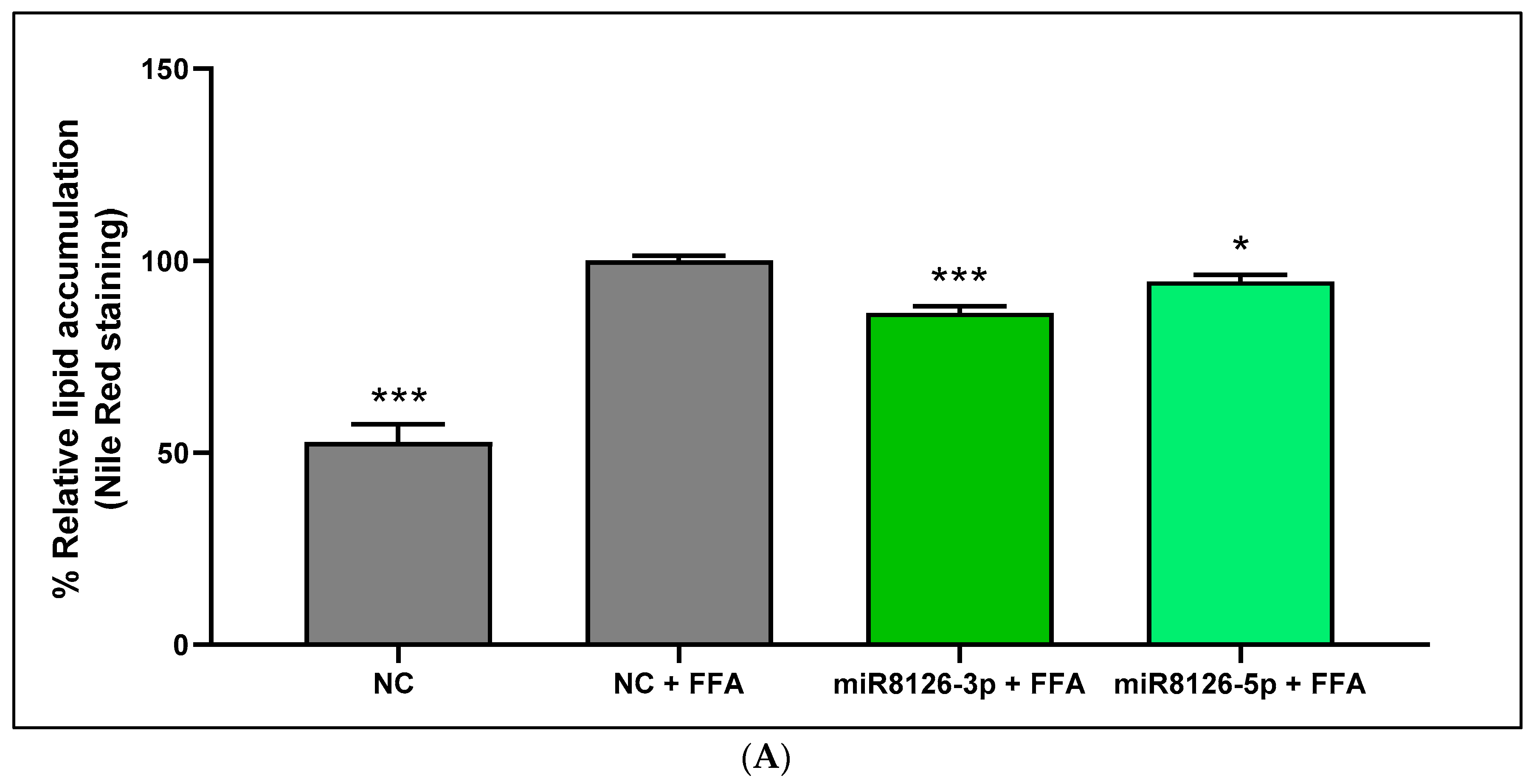
2.4. Plant miR8126-3p and miR8126-5p Modulated the Expression of Putative Target Genes and Metabolism-Associated Genes in HepG2 Cells
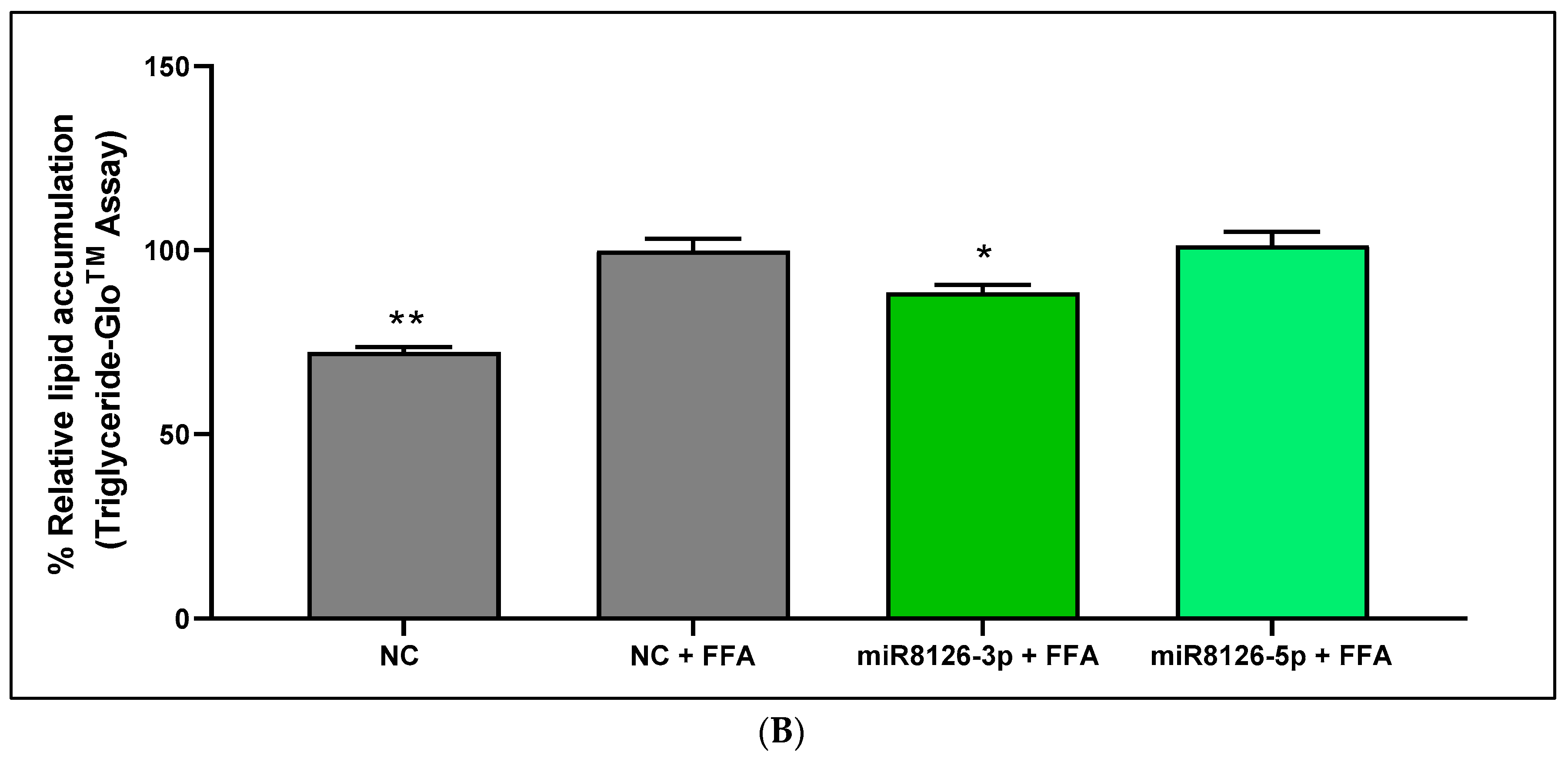
2.5. Plant miR8126-3p and miR8126-5p Decreased Lipid Accumulation in a Human Hepatocyte Cell Model That Mimicked Steatosis
3. Discussion
4. Materials and Methods
4.1. Bioinformatic Approach to Identify Putative Human Targets for Plant miRNAs
4.2. Cell Culture and Plant miRNA Mimic Transfection
4.3. RNA Isolation and Gene Expression Analyses
4.4. Cytotoxicity Assays
4.5. Intracellular Triglyceride Quantification
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Golabi, P.; Paik, J.M.; Henry, A.; Van Dongen, C.; Henry, L. The global epidemiology of nonalcoholic fatty liver disease (NAFLD) and nonalcoholic steatohepatitis (NASH): A systematic review. Hepatology 2023, 77, 1335–1347. [Google Scholar] [CrossRef] [PubMed]
- Huh, Y.; Cho, Y.J.; Nam, G.E. Recent Epidemiology and Risk Factors of Nonalcoholic Fatty Liver Disease. J. Obes. Metab. Syndr. 2022, 31, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.Q.; El-Serag, H.B.; Loomba, R. Global epidemiology of NAFLD-related HCC: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 223–238. [Google Scholar] [CrossRef]
- Liu, Y.; Zhong, G.-C.; Tan, H.-Y.; Hao, F.-B.; Hu, J.-J. Nonalcoholic fatty liver disease and mortality from all causes, cardiovascular disease, and cancer: A meta-analysis. Sci. Rep. 2019, 9, 11124. [Google Scholar] [CrossRef]
- Lazarus, J.V.; Mark, H.E.; Anstee, Q.M.; Arab, J.P.; Batterham, R.L.; Castera, L.; Cortez-Pinto, H.; Crespo, J.; Cusi, K.; Dirac, M.A.; et al. Advancing the global public health agenda for NAFLD: A consensus statement. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 60–78. [Google Scholar] [CrossRef]
- Raza, S.; Rajak, S.; Upadhyay, A.; Tewari, A.; Anthony Sinha, R. Current treatment paradigms and emerging therapies for NAFLD/NASH. Front. Biosci. 2021, 26, 206–237. [Google Scholar] [CrossRef]
- Berná, G.; Romero-Gomez, M. The role of nutrition in non-alcoholic fatty liver disease: Pathophysiology and management. Liver Int. 2020, 40 (Suppl. 1), 102–108. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.P.; Cunningham, R.P.; Dashek, R.J.; Mucinski, J.M.; Rector, R.S. A Fad too Far? Dietary Strategies for the Prevention and Treatment of NAFLD. Obesity 2020, 28, 1843–1852. [Google Scholar] [CrossRef]
- Yaskolka Meir, A.; Rinott, E.; Tsaban, G.; Zelicha, H.; Kaplan, A.; Rosen, P.; Shelef, I.; Youngster, I.; Shalev, A.; Blüher, M.; et al. Effect of green-Mediterranean diet on intrahepatic fat: The DIRECT PLUS randomised controlled trial. Gut 2021, 70, 2085–2095. [Google Scholar] [CrossRef]
- Li, X.; Peng, Z.; Li, M.; Zeng, X.; Li, H.; Zhu, Y.; Chen, H.; Hu, A.; Zhao, Q.; Zhang, Z.; et al. A Healthful Plant-Based Diet Is Associated with Lower Odds of Nonalcoholic Fatty Liver Disease. Nutrients 2022, 14, 4099. [Google Scholar] [CrossRef]
- Bagherniya, M.; Nobili, V.; Blesso, C.N.; Sahebkar, A. Medicinal plants and bioactive natural compounds in the treatment of non-alcoholic fatty liver disease: A clinical review. Pharmacol. Res. 2018, 130, 213–240. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Xin, X.; An, Z.; Hu, Y.; Feng, Q. Therapeutic Potential of Natural Plants Against Non-Alcoholic Fatty Liver Disease: Targeting the Interplay Between Gut Microbiota and Bile Acids. Front. Cell. Infect. Microbiol. 2022, 12, 854879. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-Y.; Gan, R.-Y.; Shang, A.; Mao, Q.-Q.; Sun, Q.-C.; Wu, D.-T.; Geng, F.; He, X.-Q.; Li, H.-B. Plant-Based Foods and Their Bioactive Compounds on Fatty Liver Disease: Effects, Mechanisms, and Clinical Application. Oxid. Med. Cell. Longev. 2021, 2021, 6621644. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Weng, A.; Melzig, M.F. MicroRNAs as New Bioactive Components in Medicinal Plants. Planta Med. 2016, 82, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yang, J.; Yang, Y.; Liu, J.; Li, H.; Li, R.; Cao, C.; Shi, L.; Wu, W.; He, K. A Timely Review of Cross-Kingdom Regulation of Plant-Derived MicroRNAs. Front. Genet. 2021, 12, 613197. [Google Scholar] [CrossRef]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef]
- Owusu Adjei, M.; Zhou, X.; Mao, M.; Rafique, F.; Ma, J. MicroRNAs Roles in Plants Secondary Metabolism. Plant Signal. Behav. 2021, 16, 1915590. [Google Scholar] [CrossRef]
- Dong, Q.; Hu, B.; Zhang, C. microRNAs and Their Roles in Plant Development. Front. Plant Sci. 2022, 13, 824240. [Google Scholar] [CrossRef]
- Zhang, F.; Yang, J.; Zhang, N.; Wu, J.; Si, H. Roles of microRNAs in abiotic stress response and characteristics regulation of plant. Front. Plant Sci. 2022, 13, 919243. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, Y.; Zhang, N.; Hu, M.; Zhang, H.; Joshi, T.; Xu, D. Evidence for plant-derived xenomiRs based on a large-scale analysis of public small RNA sequencing data from human samples. PLoS ONE 2018, 13, e0187519. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, P.; Wang, X.; Wang, Y.; Mu, Z.; Li, Q.; Fu, Y.; Xiao, J.; Li, G.; Ma, Y.; et al. Detection of dietetically absorbed maize-derived microRNAs in pigs. Sci. Rep. 2017, 7, 645. [Google Scholar] [CrossRef] [PubMed]
- Díez-Sainz, E.; Milagro, F.I.; Riezu-Boj, J.I.; Lorente-Cebrián, S. Effects of gut microbiota–derived extracellular vesicles on obesity and diabetes and their potential modulation through diet. J. Physiol. Biochem. 2021, 78, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, B.; Zhang, Y.; Petrick, J.S.; Heck, G.; Ivashuta, S.; Marshall, W.S. Lack of detectable oral bioavailability of plant microRNAs after feeding in mice. Nat. Biotechnol. 2013, 31, 965–967. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hou, D.; Chen, X.; Li, D.; Zhu, L.; Zhang, Y.; Li, J.; Bian, Z.; Liang, X.; Cai, X.; et al. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: Evidence of cross-kingdom regulation by microRNA. Cell Res. 2012, 22, 107–126. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, L.; Chu, Q.; Sun, S.; Wu, Y.; Tong, Z.; Fang, W.; Timko, M.P.; Fan, L. Large-scale identification of extracellular plant miRNAs in mammals implicates their dietary intake. PLoS ONE 2021, 16, e0257878. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, T.; Yin, Y.; Zhang, C.-Y.; Zhang, Y.-L. Dietary microRNA-A Novel Functional Component of Food. Adv. Nutr. 2019, 10, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Saiyed, A.N.; Vasavada, A.R.; Johar, S.R.K. Recent trends in miRNA therapeutics and the application of plant miRNA for prevention and treatment of human diseases. Futur. J. Pharm. Sci. 2022, 8, 24. [Google Scholar] [CrossRef]
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, W155–W159. [Google Scholar] [CrossRef]
- Bonnet, E.; He, Y.; Billiau, K.; Van de Peer, Y. TAPIR, a web server for the prediction of plant microRNA targets, including target mimics. Bioinformatics 2010, 26, 1566–1568. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, P.K.; Moturu, T.R.; Pandey, P.; Baldwin, I.T.; Pandey, S.P. A comparison of performance of plant miRNA target prediction tools and the characterization of features for genome-wide target prediction. BMC Genom. 2014, 15, 348. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Shin, S.; He, C.; Zhang, Q.; Rasband, M.N.; Ren, J.; Dai, C.; Zorrilla-Veloz, R.I.; Shingu, T.; Yuan, L.; et al. Qki regulates myelinogenesis through Srebp2-dependent cholesterol biosynthesis. Elife 2021, 10, e60467. [Google Scholar] [CrossRef]
- Lu, H.; Ye, Z.; Zhai, Y.; Wang, L.; Liu, Y.; Wang, J.; Zhang, W.; Luo, W.; Lu, Z.; Chen, J. QKI regulates adipose tissue metabolism by acting as a brake on thermogenesis and promoting obesity. EMBO Rep. 2020, 21, e47929. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; He, C.; Ren, J.; Dai, C.; Stevens, S.R.; Wang, Q.; Zamler, D.; Shingu, T.; Yuan, L.; Chandregowda, C.R.; et al. Mature myelin maintenance requires Qki to coactivate PPARβ-RXRα-mediated lipid metabolism. J. Clin. Investig. 2020, 130, 2220–2236. [Google Scholar] [CrossRef]
- Ruiz, M.; Coderre, L.; Allen, B.G.; Des Rosiers, C. Protecting the heart through MK2 modulation, toward a role in diabetic cardiomyopathy and lipid metabolism. Biochim. Biophys. Acta. Mol. Basis Dis. 2018, 1864, 1914–1922. [Google Scholar] [CrossRef]
- Ozcan, L.; Xu, X.; Deng, S.-X.; Ghorpade, D.S.; Thomas, T.; Cremers, S.; Hubbard, B.; Serrano-Wu, M.H.; Gaestel, M.; Landry, D.W.; et al. Treatment of Obese Insulin-Resistant Mice with an Allosteric MAPKAPK2/3 Inhibitor Lowers Blood Glucose and Improves Insulin Sensitivity. Diabetes 2015, 64, 3396–3405. [Google Scholar] [CrossRef]
- Ruiz, M.; Coderre, L.; Lachance, D.; Houde, V.; Martel, C.; Thompson Legault, J.; Gillis, M.-A.; Bouchard, B.; Daneault, C.; Carpentier, A.C.; et al. MK2 Deletion in Mice Prevents Diabetes-Induced Perturbations in Lipid Metabolism and Cardiac Dysfunction. Diabetes 2016, 65, 381–392. [Google Scholar] [CrossRef]
- Darbelli, L.; Richard, S. Emerging functions of the Quaking RNA-binding proteins and link to human diseases. Wiley Interdiscip. Rev. RNA 2016, 7, 399–412. [Google Scholar] [CrossRef]
- Gaestel, M. MAPKAP kinases—MKs—two’s company, three’s a crowd. Nat. Rev. Mol. Cell Biol. 2006, 7, 120–130. [Google Scholar] [CrossRef]
- Trempolec, N.; Muñoz, J.P.; Slobodnyuk, K.; Marin, S.; Cascante, M.; Zorzano, A.; Nebreda, A.R. Induction of oxidative metabolism by the p38α/MK2 pathway. Sci. Rep. 2017, 7, 11367. [Google Scholar] [CrossRef]
- Eynaudi, A.; Díaz-Castro, F.; Bórquez, J.C.; Bravo-Sagua, R.; Parra, V.; Troncoso, R. Differential Effects of Oleic and Palmitic Acids on Lipid Droplet-Mitochondria Interaction in the Hepatic Cell Line HepG2. Front. Nutr. 2021, 8, 775382. [Google Scholar] [CrossRef]
- Gómez-Lechón, M.J.; Donato, M.T.; Martínez-Romero, A.; Jiménez, N.; Castell, J.V.; O’Connor, J.-E. A human hepatocellular in vitro model to investigate steatosis. Chem. Biol. Interact. 2007, 165, 106–116. [Google Scholar] [CrossRef]
- Kim, S.H.; Yun, C.; Kwon, D.; Lee, Y.-H.; Kwak, J.-H.; Jung, Y.-S. Effect of Isoquercitrin on Free Fatty Acid-Induced Lipid Accumulation in HepG2 Cells. Molecules 2023, 28, 1476. [Google Scholar] [CrossRef]
- Eberlé, D.; Hegarty, B.; Bossard, P.; Ferré, P.; Foufelle, F. SREBP transcription factors: Master regulators of lipid homeostasis. Biochimie 2004, 86, 839–848. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Shimano, H.; Amemiya-Kudo, M.; Yahagi, N.; Hasty, A.H.; Matsuzaka, T.; Okazaki, H.; Tamura, Y.; Iizuka, Y.; Ohashi, K.; et al. Identification of liver X receptor-retinoid X receptor as an activator of the sterol regulatory element-binding protein 1c gene promoter. Mol. Cell. Biol. 2001, 21, 2991–3000. [Google Scholar] [CrossRef]
- Gao, W.-Y.; Chen, P.-Y.; Hsu, H.-J.; Lin, C.-Y.; Wu, M.-J.; Yen, J.-H. Tanshinone IIA Downregulates Lipogenic Gene Expression and Attenuates Lipid Accumulation through the Modulation of LXRα/SREBP1 Pathway in HepG2 Cells. Biomedicines 2021, 9, 326. [Google Scholar] [CrossRef]
- Gao, M.; Bu, L.; Ma, Y.; Liu, D. Concurrent activation of liver X receptor and peroxisome proliferator-activated receptor alpha exacerbates hepatic steatosis in high fat diet-induced obese mice. PLoS ONE 2013, 8, e65641. [Google Scholar] [CrossRef] [PubMed]
- de Bruin, R.G.; Shiue, L.; Prins, J.; de Boer, H.C.; Singh, A.; Fagg, W.S.; van Gils, J.M.; Duijs, J.M.G.J.; Katzman, S.; Kraaijeveld, A.O.; et al. Quaking promotes monocyte differentiation into pro-atherogenic macrophages by controlling pre-mRNA splicing and gene expression. Nat. Commun. 2016, 7, 10846. [Google Scholar] [CrossRef] [PubMed]
- Gnoni, G.V.; Rochira, A.; Leone, A.; Damiano, F.; Marsigliante, S.; Siculella, L. 3,5,3′triiodo-L-thyronine induces SREBP-1 expression by non-genomic actions in human HEP G2 cells. J. Cell. Physiol. 2012, 227, 2388–2397. [Google Scholar] [CrossRef] [PubMed]
- Ferré, P.; Foufelle, F. Hepatic steatosis: A role for de novo lipogenesis and the transcription factor SREBP-1c. Diabetes. Obes. Metab. 2010, 12 (Suppl. 2), 83–92. [Google Scholar] [CrossRef]
- Almatrafi, M.M.; Vergara-Jimenez, M.; Murillo, A.G.; Norris, G.H.; Blesso, C.N.; Fernandez, M.L. Moringa Leaves Prevent Hepatic Lipid Accumulation and Inflammation in Guinea Pigs by Reducing the Expression of Genes Involved in Lipid Metabolism. Int. J. Mol. Sci. 2017, 18, 1330. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.Y.; Chen, Y.; Rui, B.B.; Hu, C.M. Resveratrol ameliorates lipid accumulation in HepG2 cells, associated with down-regulation of lipin1 expression. Can. J. Physiol. Pharmacol. 2016, 94, 185–189. [Google Scholar] [CrossRef]
- Fowler, S.D.; Greenspan, P. Application of Nile red, a fluorescent hydrophobic probe, for the detection of neutral lipid deposits in tissue sections: Comparison with oil red O. J. Histochem. Cytochem. Off. J. Histochem. Soc. 1985, 33, 833–836. [Google Scholar] [CrossRef]
- Hoang, N.A.; Richter, F.; Schubert, M.; Lorkowski, S.; Klotz, L.-O.; Steinbrenner, H. Differential capability of metabolic substrates to promote hepatocellular lipid accumulation. Eur. J. Nutr. 2019, 58, 3023–3034. [Google Scholar] [CrossRef] [PubMed]
- Minami, Y.; Hoshino, A.; Higuchi, Y.; Hamaguchi, M.; Kaneko, Y.; Kirita, Y.; Taminishi, S.; Taruno, A.; Fukui, M.; Arany, Z.; et al. Liver lipophagy ameliorates nonalcoholic steatohepatitis through lysosomal lipid exocytosis. Nat. Commun. 2023, 14, 4084. [Google Scholar] [CrossRef] [PubMed]
- Belfort, R.; Harrison, S.A.; Brown, K.; Darland, C.; Finch, J.; Hardies, J.; Balas, B.; Gastaldelli, A.; Tio, F.; Pulcini, J.; et al. A placebo-controlled trial of pioglitazone in subjects with nonalcoholic steatohepatitis. N. Engl. J. Med. 2006, 355, 2297–2307. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-L.; Fu, Y.-C.; Xu, W.-C.; Feng, Y.-Q.; Fang, S.-R.; Zhou, X.-H. Resveratrol inhibits the expression of SREBP1 in cell model of steatosis via Sirt1-FOXO1 signaling pathway. Biochem. Biophys. Res. Commun. 2009, 380, 644–649. [Google Scholar] [CrossRef]
- Akao, Y.; Kuranaga, Y.; Heishima, K.; Sugito, N.; Morikawa, K.; Ito, Y.; Soga, T.; Ito, T. Plant hvu-MIR168-3p enhances expression of glucose transporter 1 (SLC2A1) in human cells by silencing genes related to mitochondrial electron transport chain complex I. J. Nutr. Biochem. 2022, 101, 108922. [Google Scholar] [CrossRef]
- Chen, T.; Ma, F.; Peng, Y.; Sun, R.; Xi, Q.; Sun, J.; Zhang, J.; Zhang, Y.; Li, M. Plant miR167e-5p promotes 3T3-L1 adipocyte adipogenesis by targeting β-catenin. Vitr. Cell Dev. Biol. Anim. 2022, 58, 471–479. [Google Scholar] [CrossRef]
- Aquilano, K.; Ceci, V.; Gismondi, A.; De Stefano, S.; Iacovelli, F.; Faraonio, R.; Di Marco, G.; Poerio, N.; Minutolo, A.; Minopoli, G.; et al. Adipocyte metabolism is improved by TNF receptor-targeting small RNAs identified from dried nuts. Commun. Biol. 2019, 2, 317. [Google Scholar] [CrossRef] [PubMed]
- Minutolo, A.; Potestà, M.; Gismondi, A.; Pirrò, S.; Cirilli, M.; Gattabria, F.; Galgani, A.; Sessa, L.; Mattei, M.; Canini, A.; et al. Olea europaea small RNA with functional homology to human miR34a in cross-kingdom interaction of anti-tumoral response. Sci. Rep. 2018, 8, 12413. [Google Scholar] [CrossRef]
- Roglia, V.; Potestà, M.; Minchella, A.; Bruno, S.P.; Bernardini, R.; Lettieri-Barbato, D.; Iacovelli, F.; Gismondi, A.; Aquilano, K.; Canini, A.; et al. Exogenous miRNAs from Moringa oleifera Lam. recover a dysregulated lipid metabolism. Front. Mol. Biosci. 2022, 9, 1012359. [Google Scholar] [CrossRef] [PubMed]
- Cavalieri, D.; Rizzetto, L.; Tocci, N.; Rivero, D.; Asquini, E.; Si-Ammour, A.; Bonechi, E.; Ballerini, C.; Viola, R. Plant microRNAs as novel immunomodulatory agents. Sci. Rep. 2016, 6, 25761. [Google Scholar] [CrossRef] [PubMed]
- Marzano, F.; Caratozzolo, M.F.; Consiglio, A.; Licciulli, F.; Liuni, S.; Sbisà, E.; D’Elia, D.; Tullo, A.; Catalano, D. Plant miRNAs Reduce Cancer Cell Proliferation by Targeting MALAT1 and NEAT1: A Beneficial Cross-Kingdom Interaction. Front. Genet. 2020, 11, 552490. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lee, H.; Jin, Y. Delivery of Functional Small RNAs via Extracellular Vesicles In Vitro and In Vivo. Methods Mol. Biol. 2020, 2115, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Müller, F.A.; Sturla, S.J. Human in vitro models of nonalcoholic fatty liver disease. Curr. Opin. Toxicol. 2019, 16, 9–16. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Cannon, C.H.; Cobb, G.P.; Anderson, T.A. Conservation and divergence of plant microRNA genes. Plant J. 2006, 46, 243–259. [Google Scholar] [CrossRef]
- Alptekin, B.; Akpinar, B.A.; Budak, H. A Comprehensive Prescription for Plant miRNA Identification. Front. Plant Sci. 2016, 7, 2058. [Google Scholar] [CrossRef]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Saez, P.; Chagoyen, M.; Tirado, F.; Carazo, J.M.; Pascual-Montano, A. GENECODIS: A web-based tool for finding significant concurrent annotations in gene lists. Genome Biol. 2007, 8, R3. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2^(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinforma. Biomath. 2013, 3, 71–85. [Google Scholar] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miR8126-3p Putative Human Target Genes | ||||
|---|---|---|---|---|
| psRNATarget. Scoring Schema V1 | ||||
| Target Accession | Expectation | UPE | mRNA Target Aligned Fragment (5′-3′) | Inhibitory Effect |
| NM_052929|FHAD1 | 2.5 | 20.471 | 63-[AUCUGUGUCAAAAUACUGAG]-82 | Cleavage |
| NM_017864|INTS8 | 2.5 | 14.728 | 108-[UUUUCAGACAAAAUACUGAA]-127 | Cleavage |
| NM_001042625|CAPSL | 3.0 | 15.911 | 110-[UUCUGUGUCAAAAUAUUGCA]-129 | Cleavage |
| NM_007359|CASC3 | 3.0 | 20.567 | 276-[UUCUGAGUCUAGAUACAGAA]-295 | Translation |
| NM_199355|ADAMTS18 | 3.0 | 12.676 | 1713-[UUUUGAGAUAAAAUGUUGAA]-1732 | Cleavage |
| NM_001098634|RBM47 | 3.0 | 11.972 | 668-[UUCUGAGUCCAAACACUGGA]-687 | Translation |
| NM_006775|QKI | 3.0 | 9.969 | 695-[AUCUGAGUUAAAAUUACUGAA]-715 | Cleavage |
| NM_206853|QKI | 3.0 | 9.969 | 5434-[AUCUGAGUUAAAAUUACUGAA]-5454 | Cleavage |
| NM_206854|QKI | 3.0 | 9.969 | 6695-[AUCUGAGUUAAAAUUACUGAA]-6715 | Cleavage |
| NM_206855|QKI | 3.0 | 9.969 | 7660-[AUCUGAGUUAAAAUUACUGAA]-7680 | Cleavage |
| NM_001104545|TMEM255A | 3.0 | 14.245 | 2064-[UUCUGCUUUAAAGUACUGAA]-2083 | Cleavage |
| psRNATarget. Scoring Schema V2 | ||||
| Target Accession | Expectation | UPE | mRNA Target Aligned Fragment (5′-3′) | Inhibitory Effect |
| NM_052929|FHAD1 | 1.5 | N/A | 63-[AUCUGUGUCAAAAUACUGAG]-82 | Cleavage |
| NM_019644|ANKRD7 | 2.5 | N/A | 285-[AUUUGUGUCAAAAUGUUGAA]-304 | Cleavage |
| NM_001099294|KIAA1644 | 2.5 | N/A | 418-[AUUUGUGUCAGAAUAUUGAA]-437 | Cleavage |
| NM_001042625|CAPSL | 3.0 | N/A | 110-[UUCUGUGUCAAAAUAUUGCA]-129 | Cleavage |
| NM_017864|INTS8 | 3.0 | N/A | 108-[UUUUCAGACAAAAUACUGAA]-127 | Cleavage |
| NM_001270458|MAOA | 3.0 | N/A | 697-[AACUGAGUUAGAAUGUUGAA]-716 | Cleavage |
| NM_004690|LATS1 | 3.0 | N/A | 3251-[AUUUGGGUCAAAAUAUUGGU]-3270 | Cleavage |
| NM_001104545|TMEM255A | 3.0 | N/A | 2064-[UUCUGCUUUAAAGUACUGAA]-2083 | Cleavage |
| NM_007359|CASC3 | 3.5 | N/A | 276-[UUCUGAGUCUAGAUACAGAA]-295 | Translation |
| NM_199355|ADAMTS18 | 3.5 | N/A | 1713-[UUUUGAGAUAAAAUGUUGAA]-1732 | Cleavage |
| NM_001098634|RBM47 | 3.5 | N/A | 668-[UUCUGAGUCCAAACACUGGA]-687 | Translation |
| NM_006775|QKI | 3.5 | N/A | 695-[AUCUGAGUUAAAAUUACUGAA]-715 | Cleavage |
| NM_206853|QKI | 3.5 | N/A | 5434-[AUCUGAGUUAAAAUUACUGAA]-5454 | Cleavage |
| NM_206854|QKI | 3.5 | N/A | 6695-[AUCUGAGUUAAAAUUACUGAA]-6715 | Cleavage |
| NM_206855|QKI | 3.5 | N/A | 7660-[AUCUGAGUUAAAAUUACUGAA]-7680 | Cleavage |
| NM_000997|RPL37 | 3.5 | N/A | 752-[UUCUGUGUUAAAGCACUGAA]-771 | Cleavage |
| NM_001040455|SIDT2 | 3.5 | N/A | 688-[UUCUGAUUCAAGAGGCUGAA]-707 | Cleavage |
| NM_001039199|TTPAL | 3.5 | N/A | 4980-[UUCUAAGUCAAAGUGGUGAA]-4999 | Cleavage |
| NM_203395|IYD | 3.5 | N/A | 738-[CAGUGAGUCAAGAUGCUGAG]-757 | Cleavage |
| NM_001164694|IYD | 3.5 | N/A | 838-[CAGUGAGUCAAGAUGCUGAG]-857 | Cleavage |
| NM_001164695|IYD | 3.5 | N/A | 925-[CAGUGAGUCAAGAUGCUGAG]-944 | Cleavage |
| NM_012069|ATP1B4 | 4.0 | N/A | 3011-[UUUUGAGUUAAAAUAGUUAA]-3030 | Cleavage |
| NM_134262|RORA | 4.0 | N/A | 6935-[UUUUGAGUCAUAAUAUUUAA]-6954 | Translation |
| NM_006311|NCOR1 | 4.5 | N/A | 1907-[UUCUGAGUCAGAAUACAGUU]-1926 | Cleavage |
| TAPIR | ||||
| Target Accession | Score | MFE Ratio | mRNA Target Aligned Fragment (5′-3′) | |
| NM_052929|FHAD1 | 2.5 | 0.8 | 63-[AUCUGUGUCAAAAUACUGAG]-82 | |
| NM_001042625|CAPSL | 4.0 | 0.78 | 110-[UUCUGUGUCAAAAUAUUGCA]-129 | |
| NM_006775|QKI | 4.0 | 0.75 | 695-[AUCUGAGUUAAAAUUACUGAA]-715 | |
| NM_206853|QKI | 4.0 | 0.75 | 5434-[AUCUGAGUUAAAAUUACUGAA]-5454 | |
| NM_206854|QKI | 4.0 | 0.75 | 6695-[AUCUGAGUUAAAAUUACUGAA]-6715 | |
| NM_206855|QKI | 4.0 | 0.75 | 7660-[AUCUGAGUUAAAAUUACUGAA]-7680 | |
| miR8126-5p Putative Human Target Genes | ||||
|---|---|---|---|---|
| psRNATarget. Scoring Schema V1 | ||||
| Target Accession | Expectation | UPE | mRNA Target Aligned Fragment (5′-3′) | Inhibitory Effect |
| NM_001201365|RNF14 | 2.5 | 19.838 | 693-[AAUUUAGUGAUACUGGCUCAGA]-714 | Translation |
| NM_032960|MAPKAPK2 | 3.0 | 16.147 | 248-[UUAUCAGUAAUUUGACUUAGA]-268 | Cleavage |
| NM_004759|MAPKAPK2 | 3.0 | 16.147 | 875-[UUAUCAGUAAUUUGACUUAGA]-895 | Cleavage |
| NM_057162|KLHL4 | 3.0 | 14.77 | 30-[CAUUUAUCAAUCUGACUCAGG]-50 | Cleavage |
| NM_019117|KLHL4 | 3.0 | 17.45 | 2890-[CAUUUAUCAAUCUGACUCAGG]-2910 | Cleavage |
| psRNATarget. Scoring Schema V2 | ||||
| Target Accession | Expectation | UPE | mRNA Target Aligned Fragment (5′-3′) | Inhibitory Effect |
| NM_032960|MAPKAPK2 | 2.0 | N/A | 248-[UUAUCAGUAAUUUGACUUAGA]-268 | Cleavage |
| NM_004759|MAPKAPK2 | 2.0 | N/A | 875-[UUAUCAGUAAUUUGACUUAGA]-895 | Cleavage |
| NM_173654|EOGT | 2.5 | N/A | 1618-[ACUUGAGUAGUCUGAUUUAGA]-1638 | Cleavage |
| NM_057162|KLHL4 | 3.0 | N/A | 30-[CAUUUAUCAAUCUGACUCAGG]-50 | Cleavage |
| NM_019117|KLHL4 | 3.0 | N/A | 2890-[CAUUUAUCAAUCUGACUCAGG]-2910 | Cleavage |
| NM_006663|PPP1R13L | 3.5 | N/A | 177-[AAUUUAGUAAUCUGCCUUAGC]-197 | Cleavage |
| NM_001001343|FNDC9 | 3.5 | N/A | 708-[UAUUAAAUAAUCUGACUUAGC]-728 | Cleavage |
| NM_020803|KLHL8 | 3.5 | N/A | 1383-[UGGUCAGUAAUCUGGUUCAUA]-1403 | Cleavage |
| NM_002076|GNS | 3.5 | N/A | 3204-[AUCUCAGUCAUUUGACUUAGA]-3224 | Cleavage |
| NM_001144966|NEDD4L | 4.0 | N/A | 4700-[UAUUGAGUAAUCUGGUUUCGA]-4720 | Cleavage |
| NM_001146213|TBC1D15 | 4.0 | N/A | 1927-[GAUACAGUUAUUUGACUCAGU]-1947 | Cleavage |
| NM_001201365|RNF14 | 4.5 | N/A | 693-[AAUUUAGUGAUACUGGCUCAGA]-714 | Translation |
| NM_001168724|TMEM135 | 4.5 | N/A | 3417-[UAUUAAGUAGUUUGACACAGC]-3437 | Cleavage |
| NM_207304|MBNL2 | 4.5 | N/A | 2298-[UAUUCAGAAGUCUGACUAUGA]-2318 | Cleavage |
| NM_144778|MBNL2 | 4.5 | N/A | 2411-[UAUUCAGAAGUCUGACUAUGA]-2431 | Cleavage |
| NM_001202485|HSPE1-MOB4 | 4.5 | N/A | 1820-[GAUUUGGUGAUCUGGCUGAGU]-1840 | Cleavage |
| NM_001002909|GPATCH8 | 5.0 | N/A | 1921-[UGUUCAGUACUCAUCUGAUUCAGA]-1944 | Cleavage |
| NM_030920|ANP32E | 5.0 | N/A | 593-[UAUUCAGUAAUAUGGUUCAUG]-613 | Translation |
| NM_001171909|CXorf40A | 5.0 | N/A | 406-[UGUUCAGUGUUCUGACUCGCC]-426 | Cleavage |
| NM_001041|SI | 5.0 | N/A | 164-[UAUUAUAGUAAUGUGACUUGGA]-185 | Translation |
| TAPIR | ||||
| Target Accession | Score | MFE Ratio | mRNA Target Aligned Fragment (5′-3′) | |
| NM_032960|MAPKAPK2 | 4.0 | 0.76 | 248-[UUAUCAGUAAUUUGACUUAGA]-268 | |
| NM_004759|MAPKAPK2 | 4.0 | 0.76 | 875-[UUAUCAGUAAUUUGACUUAGA]-895 | |
| Gene Name | Assay ID | RefSeq |
|---|---|---|
| TBP | 1Hs00427620_m1 | NM_001172085.1NM_003194.4 |
| QKI | 2Hs.PT.58.2815647 | NM_006775(1) |
| MAPKAPK2 | 2Hs.PT.58.2443418 | NM_004759(2) |
| PPARA | 2Hs.PT.58.45310483 | NM_001001928(2) |
| RXRA | 2Hs.PT.58.3784663 | NM_002957(1) |
| FOXO1 | 2Hs.PT.58.40005627 | NM_002015(1) |
| SREBF1 | 2Hs.PT.58.3359761 | NM_001005291(2) |
| FASN | 1Hs01005622_m1 | NM_004104.4XM_011523538.2 |
| ACOX1 | 2Hs.PT.56a.3058584 | NM_001185039(3) |
| G6PC | 2Hs.PT.58.5006581 | NM_000151(2) |
| GSK3B | 2Hs.PT.58.40111551 | NM_001146156(2) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díez-Sainz, E.; Aranaz, P.; Amri, E.-Z.; Riezu-Boj, J.I.; Lorente-Cebrián, S.; Milagro, F.I. Plant miR8126-3p and miR8126-5p Decrease Lipid Accumulation through Modulation of Metabolic Genes in a Human Hepatocyte Model That Mimics Steatosis. Int. J. Mol. Sci. 2024, 25, 1721. https://doi.org/10.3390/ijms25031721
Díez-Sainz E, Aranaz P, Amri E-Z, Riezu-Boj JI, Lorente-Cebrián S, Milagro FI. Plant miR8126-3p and miR8126-5p Decrease Lipid Accumulation through Modulation of Metabolic Genes in a Human Hepatocyte Model That Mimics Steatosis. International Journal of Molecular Sciences. 2024; 25(3):1721. https://doi.org/10.3390/ijms25031721
Chicago/Turabian StyleDíez-Sainz, Ester, Paula Aranaz, Ez-Zoubir Amri, José I. Riezu-Boj, Silvia Lorente-Cebrián, and Fermín I. Milagro. 2024. "Plant miR8126-3p and miR8126-5p Decrease Lipid Accumulation through Modulation of Metabolic Genes in a Human Hepatocyte Model That Mimics Steatosis" International Journal of Molecular Sciences 25, no. 3: 1721. https://doi.org/10.3390/ijms25031721