
Members of the Capsicum annuum CaTrxh Family Respond to High Temperature and Exhibit Dynamic Hetero/Homo Interactions


Abstract
1. Introduction
2. Results
2.1. Genome-Wide Identification and Characterization of the CaTrxh-Type Family in Pepper
2.2. Cis-Regulatory Element Analysis of CaTrxh-Type Gene Promoter
2.3. Tissue-Specific Gene Expression of CaTrxh Family
2.4. Expression Analysis of CaTrxh Genes in Response to Heat and Cold Stresses
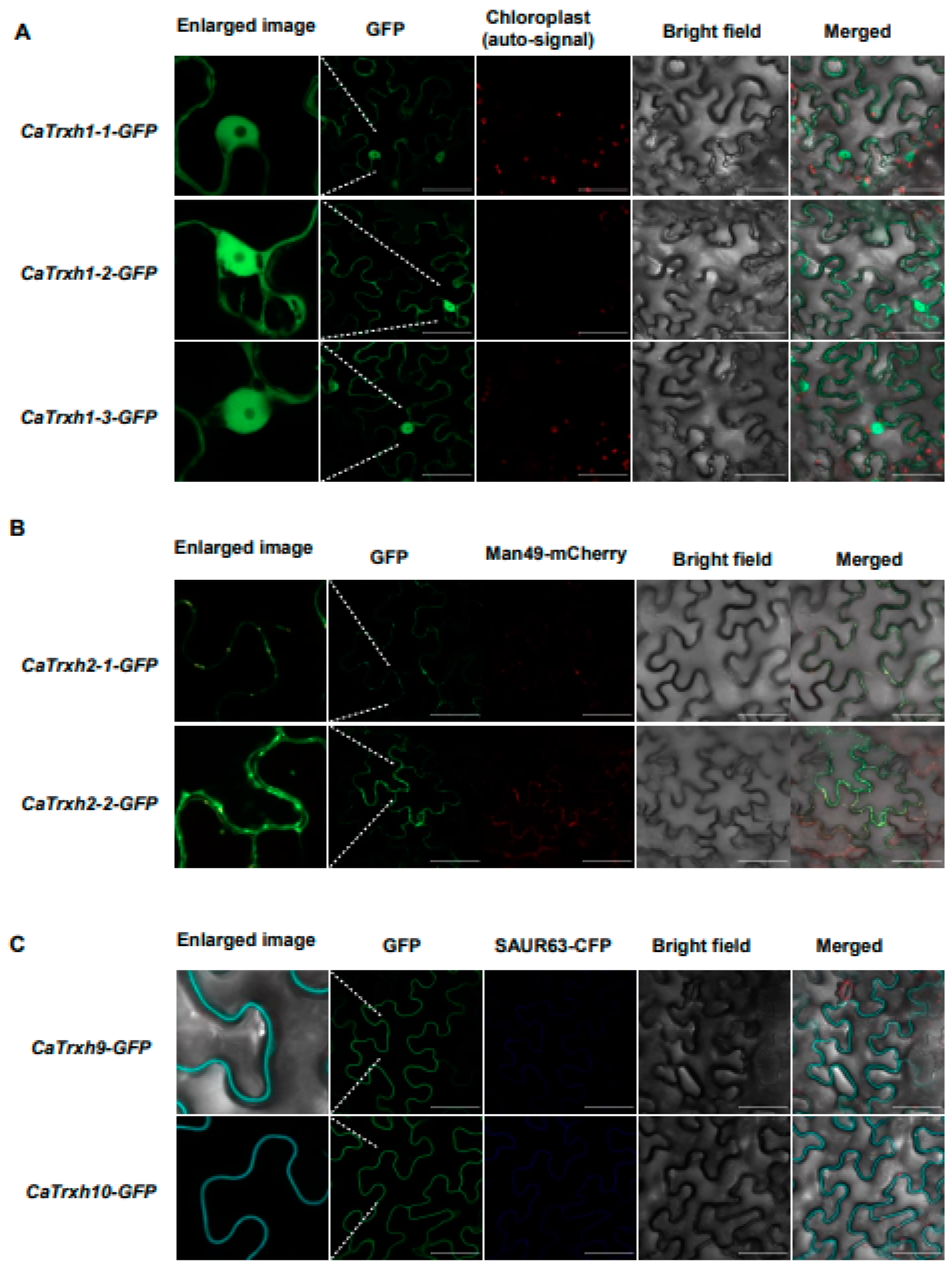
2.5. Subcellular Localization of the CaTrxh Family
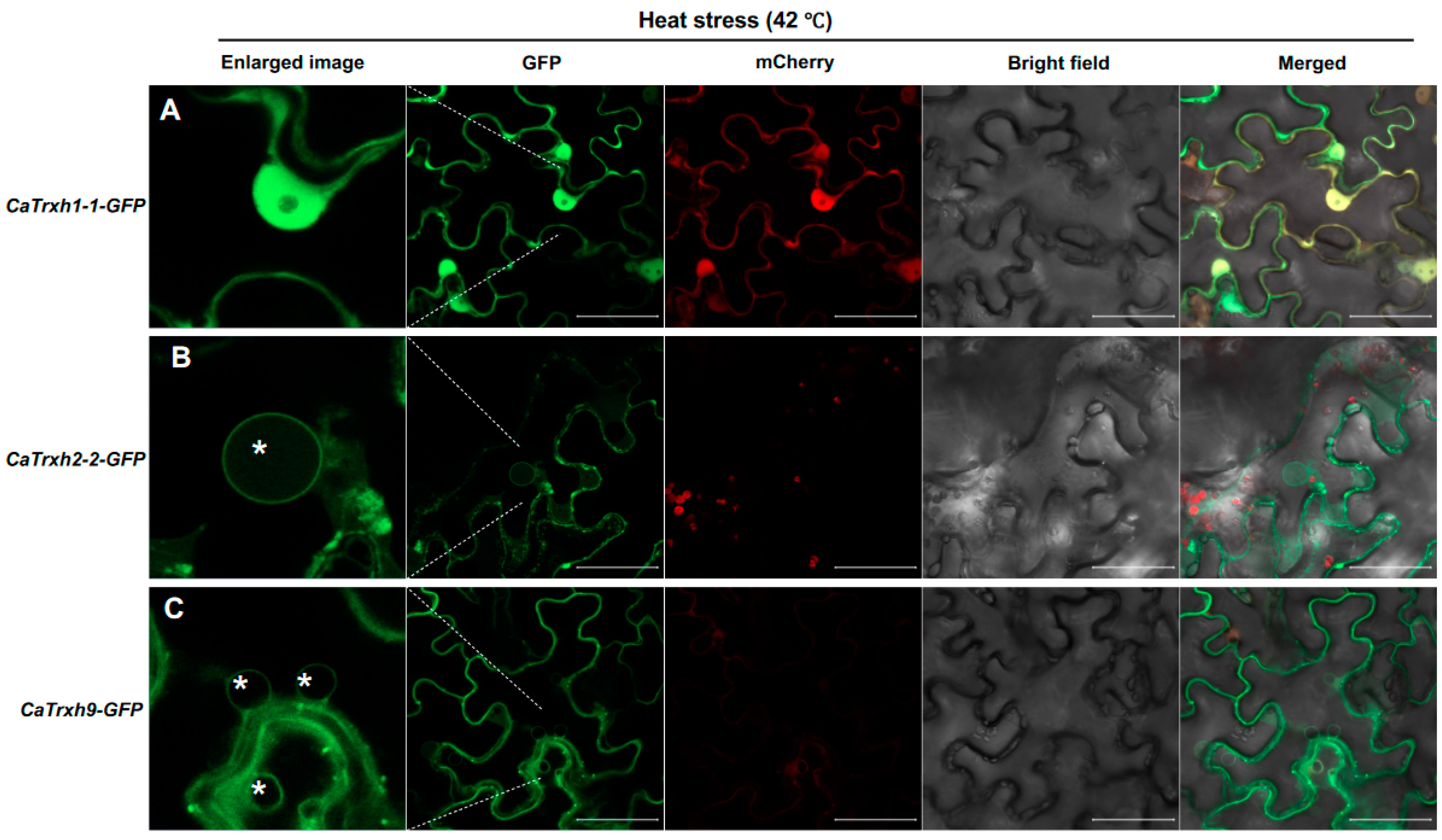
2.6. CaTrxh Protein Changes in Subcellular Localization under Heat Stress
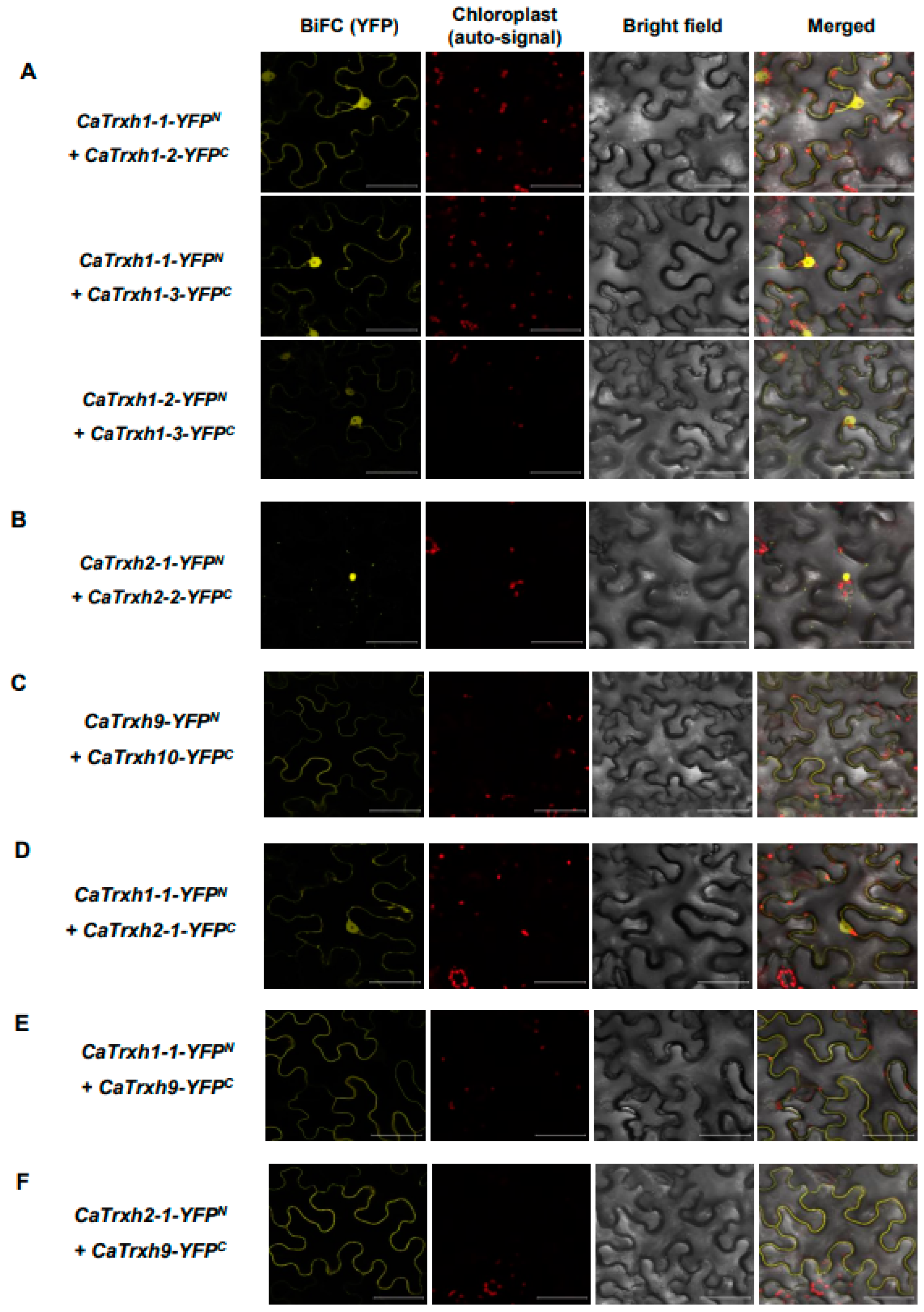
2.7. CaTrxh Proteins Were Able to Make Complexes via Homomeric Association
2.8. CaTrxh Proteins Exhibited Heteromeric Protein Complexes
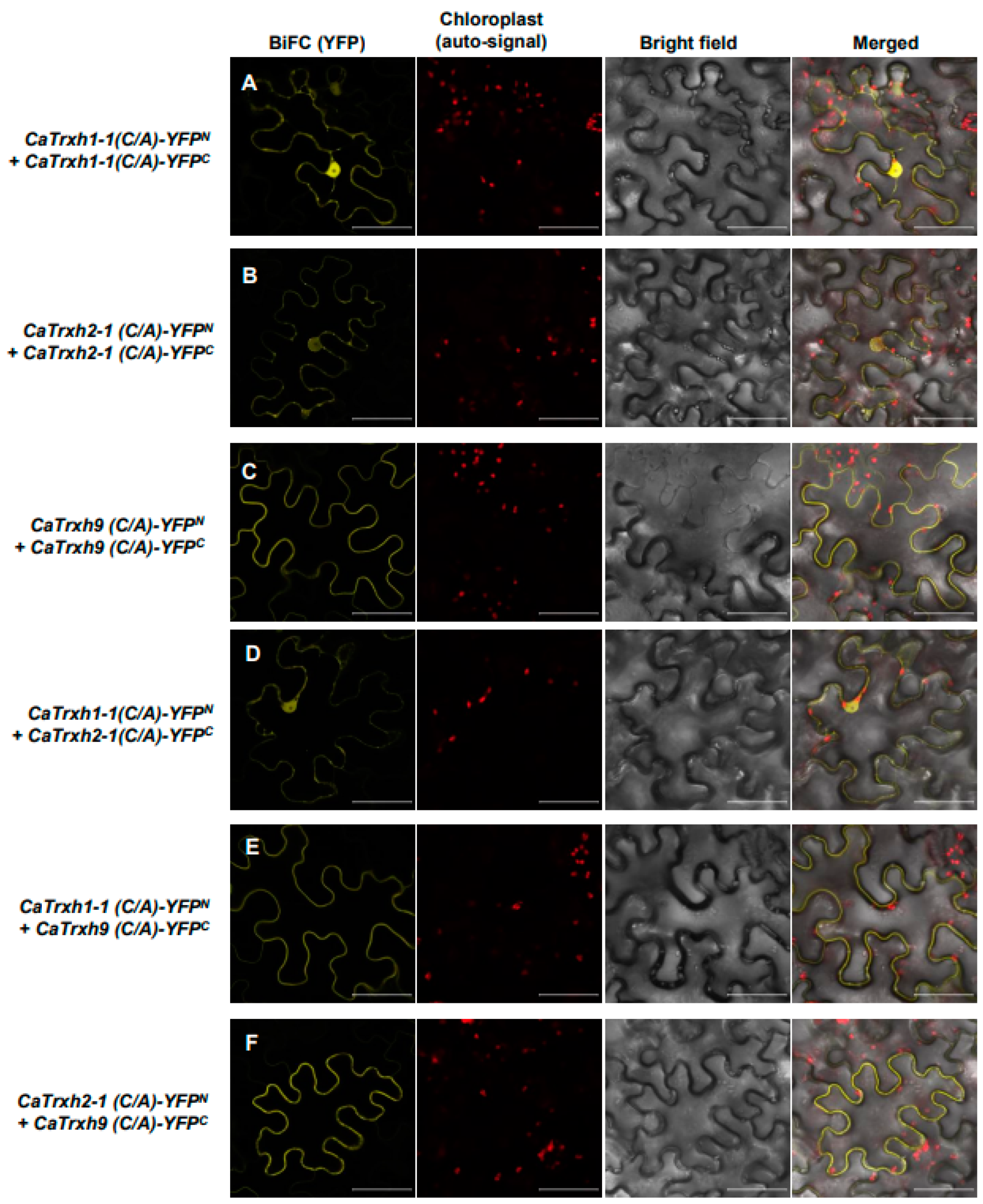
2.9. Thioredoxin Protein Activity Is Not Related with CaTrxh Protein Complex and Relocalization
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Sample RNA Extraction, Semi-Quantitative RT-PCR, and qRT-PCR
4.3. Identification, Protein Sequence Alignment, and Phylogenetic Tree of CaTrxh-Type Subfamily
4.4. Plasmid Construct of CaTrx-h-Type Genes and Site-Direct Mutagenesis
4.5. BiFC Analysis and Subcellular Localization Analysis
4.6. Promoter Analysis of CaTrxh-Type Genes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, L.; Kantar, M.B.; Moxley, D.; Ortiz-Barrientos, D.; Rieseberg, L.H. Crop adaptation to climate change: An evolutionary perspective. Mol. Plant 2023, 16, 1518–1546. [Google Scholar] [CrossRef] [PubMed]
- Wani, K.I.; Naeem, M.; Castroverde, C.D.M.; Kalaji, H.M.; Albaqami, M.; Aftab, T. Molecular Mechanisms of Nitric Oxide (NO) Signaling and Reactive Oxygen Species (ROS) Homeostasis during Abiotic Stresses in Plants. Int. J. Mol. Sci. 2021, 22, 9656. [Google Scholar] [CrossRef] [PubMed]
- Devireddy, A.R.; Rivero, R.M.; Zandalinas, S.I. Editorial: Rising stars in plant ROS/redox biology under abiotic stress conditions. Front. Plant Sci. 2023, 14, 1207275. [Google Scholar] [CrossRef]
- Meyer, Y.; Buchanan, B.B.; Vignols, F.; Reichheld, J.P. Thioredoxins and glutaredoxins: Unifying elements in redox biology. Annu. Rev. Genet. 2009, 43, 335–367. [Google Scholar] [CrossRef]
- Sevilla, F.; Marti, M.C.; De Brasi-Velasco, S.; Jimenez, A. Redox regulation, thioredoxins, and glutaredoxins in retrograde signalling and gene transcription. J. Exp. Bot. 2023, 74, 5955–5969. [Google Scholar] [CrossRef]
- Geigenberger, P.; Thormahlen, I.; Daloso, D.M.; Fernie, A.R. The Unprecedented Versatility of the Plant Thioredoxin System. Trends Plant Sci. 2017, 22, 249–262. [Google Scholar] [CrossRef]
- Ji, M.G.; Park, H.J.; Cha, J.-Y.; Kim, J.A.; Shin, G.-I.; Jeong, S.Y.; Lee, E.S.; Yun, D.-J.; Lee, S.Y.; Kim, W.-Y. Expression of Arabidopsis thaliana Thioredoxin-h2 in Brassica napus enhances antioxidant defenses and improves salt tolerance. Plant Physiol. Biochem. 2020, 147, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Li, Q.; Yan, H.; Zhang, D.; Jiang, G.; Jiang, Y.; Duan, X. Characteristics of Three Thioredoxin Genes and Their Role in Chilling Tolerance of Harvested Banana Fruit. Int. J. Mol. Sci. 2016, 17, 1526. [Google Scholar] [CrossRef]
- Svensson, M.J.; Larsson, J. Thioredoxin-2 affects lifespan and oxidative stress in Drosophila. Hereditas 2007, 144, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Ogata, F.T.; Batista, W.L.; Sartori, A.; Gesteira, T.F.; Masutani, H.; Arai, R.J.; Yodoi, J.; Stern, A.; Monteiro, H.P. Nitrosative/oxidative stress conditions regulate thioredoxin-interacting protein (TXNIP) expression and thioredoxin-1 (TRX-1) nuclear localization. PLoS ONE 2013, 8, e84588. [Google Scholar] [CrossRef] [PubMed]
- Sugano, E.; Murayama, N.; Takahashi, M.; Tabata, K.; Tamai, M.; Tomita, H. Essential role of thioredoxin 2 in mitigating oxidative stress in retinal epithelial cells. J. Ophthalmol. 2013, 2013, 185825. [Google Scholar] [CrossRef] [PubMed]
- Chibani, K.; Pucker, B.; Dietz, K.J.; Cavanagh, A. Genome-wide analysis and transcriptional regulation of the typical and atypical thioredoxins in Arabidopsis thaliana. FEBS Lett. 2021, 595, 2715–2730. [Google Scholar] [CrossRef]
- Montrichard, F.; Alkhalfioui, F.; Yano, H.; Vensel, W.H.; Hurkman, W.J.; Buchanan, B.B. Thioredoxin targets in plants: The first 30 years. J. Proteom. 2009, 72, 452–474. [Google Scholar] [CrossRef] [PubMed]
- Sainz, M.M.; Filippi, C.V.; Eastman, G.; Sotelo-Silveira, J.; Borsani, O.; Sotelo-Silveira, M. Analysis of Thioredoxins and Glutaredoxins in Soybean: Evidence of Translational Regulation under Water Restriction. Antioxidants 2022, 11, 1622. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, Y.; Huang, X. Genome-Wide Analysis of the Thioredoxin Gene Family in Gossypium hirsutum L. and the Role of the Atypical Thioredoxin Gene GhTRXL3-2 in Flowering. J. Plant Biol. 2021, 64, 461–473. [Google Scholar] [CrossRef]
- Meyer, Y.; Siala, W.; Bashandy, T.; Riondet, C.; Vignols, F.; Reichheld, J.P. Glutaredoxins and thioredoxins in plants. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2008, 1783, 589–600. [Google Scholar] [CrossRef]
- Tada, Y.; Spoel, S.H.; Pajerowska-Mukhtar, K.; Mou, Z.; Song, J.; Wang, C.; Zuo, J.; Dong, X. Plant immunity requires conformational changes [corrected] of NPR1 via S-nitrosylation and thioredoxins. Science 2008, 321, 952–956. [Google Scholar] [CrossRef]
- Kneeshaw, S.; Gelineau, S.; Tada, Y.; Loake, G.J.; Spoel, S.H. Selective protein denitrosylation activity of Thioredoxin-h5 modulates plant Immunity. Mol. Cell 2014, 56, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.S.; Park, J.H.; Wi, S.D.; Chae, H.B.; Paeng, S.K.; Bae, S.B.; Phan, K.A.T.; Lee, S.Y. Arabidopsis Disulfide Reductase, Trx-h2, Functions as an RNA Chaperone under Cold Stress. Appl. Sci. 2021, 11, 6865. [Google Scholar] [CrossRef]
- Zhai, J.; Qi, Q.; Wang, M.; Yan, J.; Li, K.; Xu, H. Overexpression of tomato thioredoxin h (SlTrxh) enhances excess nitrate stress tolerance in transgenic tobacco interacting with SlPrx protein. Plant Sci. 2022, 315, 111137. [Google Scholar] [CrossRef]
- Chung, Y.B.; Lee, H.; Hwang, S.; Seo, H.Y.; Suh, H.J.; Jo, K. Effect of capsaicinoids in hot pepper powder on microbial community and free sugar during kimchi fermentation. J. Food Sci. 2021, 86, 3195–3204. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wan, Y.; Meng, Y.; Zhou, Q.; Li, B.; Chen, Y.; Wang, L. Capsaicin: A Novel Approach to the Treatment of Functional Dyspepsia. Mol. Nutr. Food Res. 2023, 67, e2200793. [Google Scholar] [CrossRef]
- Tiamiyu, Q.O.; Adebayo, S.E.; Ibrahim, N. Recent advances on postharvest technologies of bell pepper: A review. Heliyon 2023, 9, e15302. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Dai, T.; Sun, W.; Wei, Y.; Ren, J.; Zhang, L.; Zhang, M.; Zhou, F. Protein N-myristoylation: Functions and mechanisms in control of innate immunity. Cell. Mol. Immunol. 2021, 18, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Majeran, W.; Le Caer, J.P.; Ponnala, L.; Meinnel, T.; Giglione, C. Targeted Profiling of Arabidopsis thaliana Subproteomes Illuminates Co- and Posttranslationally N-Terminal Myristoylated Proteins. Plant Cell 2018, 30, 543–562. [Google Scholar] [CrossRef] [PubMed]
- Traverso, J.A.; Micalella, C.; Martinez, A.; Brown, S.C.; Satiat-Jeunemaître, B.; Meinnel, T.; Giglione, C. Roles of N-terminal fatty acid acylations in membrane compartment partitioning: Arabidopsis h-type thioredoxins as a case study. Plant Cell 2013, 25, 1056–1077. [Google Scholar] [CrossRef] [PubMed]
- Muhammad Aslam, M.; Waseem, M.; Jakada, B.H.; Okal, E.J.; Lei, Z.; Saqib, H.S.A.; Yuan, W.; Xu, W.; Zhang, Q. Mechanisms of Abscisic Acid-Mediated Drought Stress Responses in Plants. Int. J. Mol. Sci. 2022, 23, 1084. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Azhar, M.T.; Hinze, L.; Qayyum, A.; Li, H.; Peng, Z.; Qin, G.; Jia, Y.; Pan, Z.; He, S.; et al. Insight into abscisic acid perception and signaling to increase plant tolerance to abiotic stress. J. Plant Interact. 2021, 16, 222–237. [Google Scholar] [CrossRef]
- Marcotte, W.R., Jr.; Russell, S.H.; Quatrano, R.S. Abscisic acid-responsive sequences from the em gene of wheat. Plant Cell 1989, 1, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Guiltinan, M.J.; Marcotte, W.R., Jr.; Quatrano, R.S. A plant leucine zipper protein that recognizes an abscisic acid response element. Science 1990, 250, 267–271. [Google Scholar] [CrossRef]
- Agarwal, M.; Hao, Y.; Kapoor, A.; Dong, C.H.; Fujii, H.; Zheng, X.; Zhu, J.K. A R2R3 type MYB transcription factor is involved in the cold regulation of CBF genes and in acquired freezing tolerance. J. Biol. Chem. 2006, 281, 37636–37645. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.Q.; Shen, C.; Wu, L.H.; Tang, K.X.; Lin, J. CBF-dependent signaling pathway: A key responder to low temperature stress in plants. Crit. Rev. Biotechnol. 2011, 31, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Wang, Z.; Zhang, Y.; Li, H.; Yang, M.; Yin, H.; Cui, J.; Chai, H.; Gao, Y.; Hu, G.; et al. Overexpression of a Thioredoxin-Protein-Encoding Gene, MsTRX, from Medicago sativa Enhances Salt Tolerance to Transgenic Tobacco. Agronomy 2022, 12, 1467. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, E.S.; Chae, H.B.; Paeng, S.K.; Wi, S.D.; Bae, S.B.; Thi Phan, K.A.; Lee, S.Y. Disulfide reductase activity of thioredoxin-h2 imparts cold tolerance in Arabidopsis. Biochem. Biophys. Res. Commun. 2021, 568, 124–130. [Google Scholar] [CrossRef]
- Meyer, Y.; Reichheld, J.P.; Vignols, F. Thioredoxins in Arabidopsis and other plants. Photosynth. Res. 2005, 86, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Hara, S.; Hisabori, T. Thioredoxin Selectivity for Thiol-based Redox Regulation of Target Proteins in Chloroplasts. J. Biol. Chem. 2015, 290, 14278–14288. [Google Scholar] [CrossRef]
- Thormählen, I.; Zupok, A.; Rescher, J.; Leger, J.; Weissenberger, S.; Groysman, J.; Orwat, A.; Chatel-Innocenti, G.; Issakidis-Bourguet, E.; Armbruster, U.; et al. Thioredoxins Play a Crucial Role in Dynamic Acclimation of Photosynthesis in Fluctuating Light. Mol. Plant 2017, 10, 168–182. [Google Scholar] [CrossRef] [PubMed]
- Ying, Y.; Yue, W.; Wang, S.; Li, S.; Wang, M.; Zhao, Y.; Wang, C.; Mao, C.; Whelan, J.; Shou, H. Two h-Type Thioredoxins Interact with the E2 Ubiquitin Conjugase PHO2 to Fine-Tune Phosphate Homeostasis in Rice. Plant Physiol. 2017, 173, 812–824. [Google Scholar] [CrossRef]
- Cejudo, F.J.; González, M.C.; Pérez-Ruiz, J.M. Redox regulation of chloroplast metabolism. Plant Physiol. 2021, 186, 9–21. [Google Scholar] [CrossRef]
- Meng, L.; Wong, J.H.; Feldman, L.J.; Lemaux, P.G.; Buchanan, B.B. A membrane-associated thioredoxin required for plant growth moves from cell to cell, suggestive of a role in intercellular communication. Proc. Natl. Acad. Sci. USA 2010, 107, 3900–3905. [Google Scholar] [CrossRef]
- Lee, E.S.; Park, J.H.; Wi, S.D.; Kang, C.H.; Chi, Y.H.; Chae, H.B.; Paeng, S.K.; Ji, M.G.; Kim, W.Y.; Kim, M.G.; et al. Redox-dependent structural switch and CBF activation confer freezing tolerance in plants. Nat. Plants 2021, 7, 914–922. [Google Scholar] [CrossRef]
- Yoshida, K.; Hisabori, T. Current Insights into the Redox Regulation Network in Plant Chloroplasts. Plant Cell Physiol. 2023, 64, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Yano, H. Ongoing applicative studies of plant thioredoxins. Mol. Plant 2014, 7, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, C.; Cheng, F.F.; Guo, X.Y.; Li, Y.X.; Wang, A.Y.; Zhu, J.B. Molecular cloning and functional analysis of the thioredoxin gene SikTrxh from Saussurea involucrata. Biol. Plant. 2021, 65, 47–59. [Google Scholar] [CrossRef]
- Park, S.K.; Jung, Y.J.; Lee, J.R.; Lee, Y.M.; Jang, H.H.; Lee, S.S.; Park, J.H.; Kim, S.Y.; Moon, J.C.; Lee, S.Y.; et al. Heat-shock and redox-dependent functional switching of an h-type Arabidopsis thioredoxin from a disulfide reductase to a molecular chaperone. Plant Physiol. 2009, 150, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Arnaiz, A.; Romero-Puertas, M.C.; Santamaria, M.E.; Rosa-Diaz, I.; Arbona, V.; Munoz, A.; Grbic, V.; Gonzalez-Melendi, P.; Mar Castellano, M.; Sandalio, L.M.; et al. The Arabidopsis thioredoxin TRXh5regulates the S-nitrosylation pattern of the TIRK receptor being both proteins essential in the modulation of defences to Tetranychus urticae. Redox Biol. 2023, 67, 102902. [Google Scholar] [CrossRef]
- Kinkema, M.; Fan, W.; Dong, X. Nuclear localization of NPR1 is required for activation of PR gene expression. Plant Cell 2000, 12, 2339–2350. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Offenborn, J.N.; Waadt, R.; Kudla, J. Visualization and translocation of ternary Calcineurin-A/Calcineurin-B/Calmodulin-2 protein complexes by dual-color trimolecular fluorescence complementation. New Phytol. 2015, 208, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Huh, S.U. Optimization of immune receptor-related hypersensitive cell death response assay using agrobacterium-mediated transient expression in tobacco plants. Plant Methods 2022, 18, 57. [Google Scholar] [CrossRef]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | |
|---|---|---|
| Arabidopsis thaliana | AtTrxh1 | NP_190672.1 |
| AtTrxh2 | NP_198811.1 | |
| AtTrxh3 | NP_199112.1 | |
| AtTrxh4 | NP_173403.1 | |
| AtTrxh5 | NP_175128.1 | |
| AtTrxh7 | NP_176182.1 | |
| AtTrxh8 | NP_177146.1 | |
| AtTrxh9 | NP_001078124.1 | |
| AtTrxh10 | NP_001325846.1 | |
| AtCXXS1 | NP_172620.1 | |
| AtCXXS2 | NP_001318396.1 | |
| Oryza sativa | OsTrxh1 | XP_015647572.1 |
| OsTrxh2 | XP_015640796.1 | |
| OsTrxh4 | XP_015631704.1 | |
| OsTrxh5 | XP_015646978.1 | |
| OsTrxh7 | XP_015625017.1 | |
| OsTrxh8 | XP_015640764.1 | |
| OsTrxh9 | XP_015639270.1 | |
| Solanum tuberosum | StTrxh1 | XP_006341987.1 |
| StTrxh2 | NP_001275313.1 | |
| Solanum lycopersicum | SlTrxh1 | XP_004238306.1 |
| SlTrxh2 | XP_010316263.1 | |
| SlTrxh4 | XP_004235220.1 | |
| SlTrxh9 | XP_004246283.1 | |
| Nicotiana tabacum | NtTrxh1 | CAA41415.1 |
| NtTrxh2 | Q07090.1 | |
| NtTrxh3 | XP_016458904.1 | |
| Capsicum annuum | CaTrxh1-1 | XP_016568496.1 |
| CaTrxh1-2 | XP_016560703.1 | |
| CaTrxh1-3 | XP_016552903.1 | |
| CaTrxh2-1 | XP_016548658.2 | |
| CaTrxh2-2 | XP_016561441.1 | |
| CaTrxh9 | XP_016541800.1 | |
| CaTrxh10 | XP_016564926.1 |
| Gene Name | Accession Number | Chromosome | Length (aa) | Molecular Weight | Theoretical pI | Instability Index | Aliphatic Index | GRAVY | Stability |
|---|---|---|---|---|---|---|---|---|---|
| CaTrxh1-1 | XM_016713010.1 | 4 | 124 | 13,717.77 | 5.01 | 30.8 | 85.65 | 0.008 | YES |
| CaTrxh1-2 | XM_016705217.1 | 2 | 119 | 13,186.19 | 5.3 | 25.34 | 86.81 | −0.047 | YES |
| CaTrxh1-3 | XM_016697417.1 | 4 | 117 | 12,849.94 | 5.34 | 24.2 | 94.19 | 0.067 | YES |
| CaTrxh2-1 | XM_016693172.1 | 11 | 135 | 15,286.76 | 6.96 | 20.14 | 88.81 | −0.136 | YES |
| CaTrxh2-2 | XM_016705955.1 | 2 | 142 | 15,655.87 | 5.23 | 44.58 | 81.06 | −0.185 | NO |
| CaTrxh9 | XM_016686314.1 | 9 | 152 | 16,830.12 | 4.82 | 30.95 | 77.7 | −0.189 | YES |
| CaTrxh10 | XM_016709440.1 | 3 | 138 | 15,415.41 | 5.06 | 34.65 | 69.93 | −0.463 | YES |
| Cis-Element | Function | CaTrxh1-1 | CaTrxh1-2 | CaTrxh1-3 | CaTrxh2-1 | CaTrxh2-2 | CaTrxh9 | CaTrxh10 |
|---|---|---|---|---|---|---|---|---|
| Hormone-related | ||||||||
| ABRE | cis-element involved in the abscisic acid responsiveness | 3 | - | 2 | 2 | 1 | 5 | 1 |
| CGTCA-motif | cis-element involved in the MeJA-responsiveness | 1 | - | - | 1 | 1 | - | - |
| ERE | ethylene-responsive element | 7 | - | 1 | - | 2 | - | 1 |
| GARE-motif | gibberellin-responsive element | - | - | - | - | - | 1 | 1 |
| MYC | cis-acting element involved in the abscisic acid responsiveness | - | 6 | 1 | 3 | 3 | 3 | 3 |
| P-box | gibberellin-responsive element | 1 | 1 | - | - | - | 1 | - |
| TATC-box | cis-element involved in gibberellin-responsiveness | 1 | - | - | - | - | 1 | 1 |
| TCA-element | cis-element involved in salicylic acid responsiveness | 1 | 1 | 1 | - | 1 | 1 | 1 |
| TGA-box | part of an auxin-responsive element | - | - | - | 1 | - | - | - |
| TGA-element | auxin-responsive element | - | - | - | - | - | - | - |
| TGACG-motif | cis-acting regulatory element involved in the MeJA-responsiveness | 1 | - | - | 1 | 1 | - | - |
| Stress-related | ||||||||
| ARE | cis-element essential for the anaerobic induction | - | - | - | 1 | 1 | - | 1 |
| LTR | cis-element involved in low-temperature responsiveness | 1 | - | 1 | - | - | 1 | - |
| MBS | MYB binding site involved in drought-inducibility | - | 1 | 2 | - | 2 | - | - |
| TC-rich repeats | cis-element involved in defense and stress responsiveness | - | - | 2 | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, S.; Huh, S.U. Members of the Capsicum annuum CaTrxh Family Respond to High Temperature and Exhibit Dynamic Hetero/Homo Interactions. Int. J. Mol. Sci. 2024, 25, 1729. https://doi.org/10.3390/ijms25031729
Hong S, Huh SU. Members of the Capsicum annuum CaTrxh Family Respond to High Temperature and Exhibit Dynamic Hetero/Homo Interactions. International Journal of Molecular Sciences. 2024; 25(3):1729. https://doi.org/10.3390/ijms25031729
Chicago/Turabian StyleHong, Suji, and Sung Un Huh. 2024. "Members of the Capsicum annuum CaTrxh Family Respond to High Temperature and Exhibit Dynamic Hetero/Homo Interactions" International Journal of Molecular Sciences 25, no. 3: 1729. https://doi.org/10.3390/ijms25031729
APA StyleHong, S., & Huh, S. U. (2024). Members of the Capsicum annuum CaTrxh Family Respond to High Temperature and Exhibit Dynamic Hetero/Homo Interactions. International Journal of Molecular Sciences, 25(3), 1729. https://doi.org/10.3390/ijms25031729