
Rosavin Alleviates LPS-Induced Acute Lung Injure by Modulating the TLR-4/NF-κB/MAPK Singnaling Pathways
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
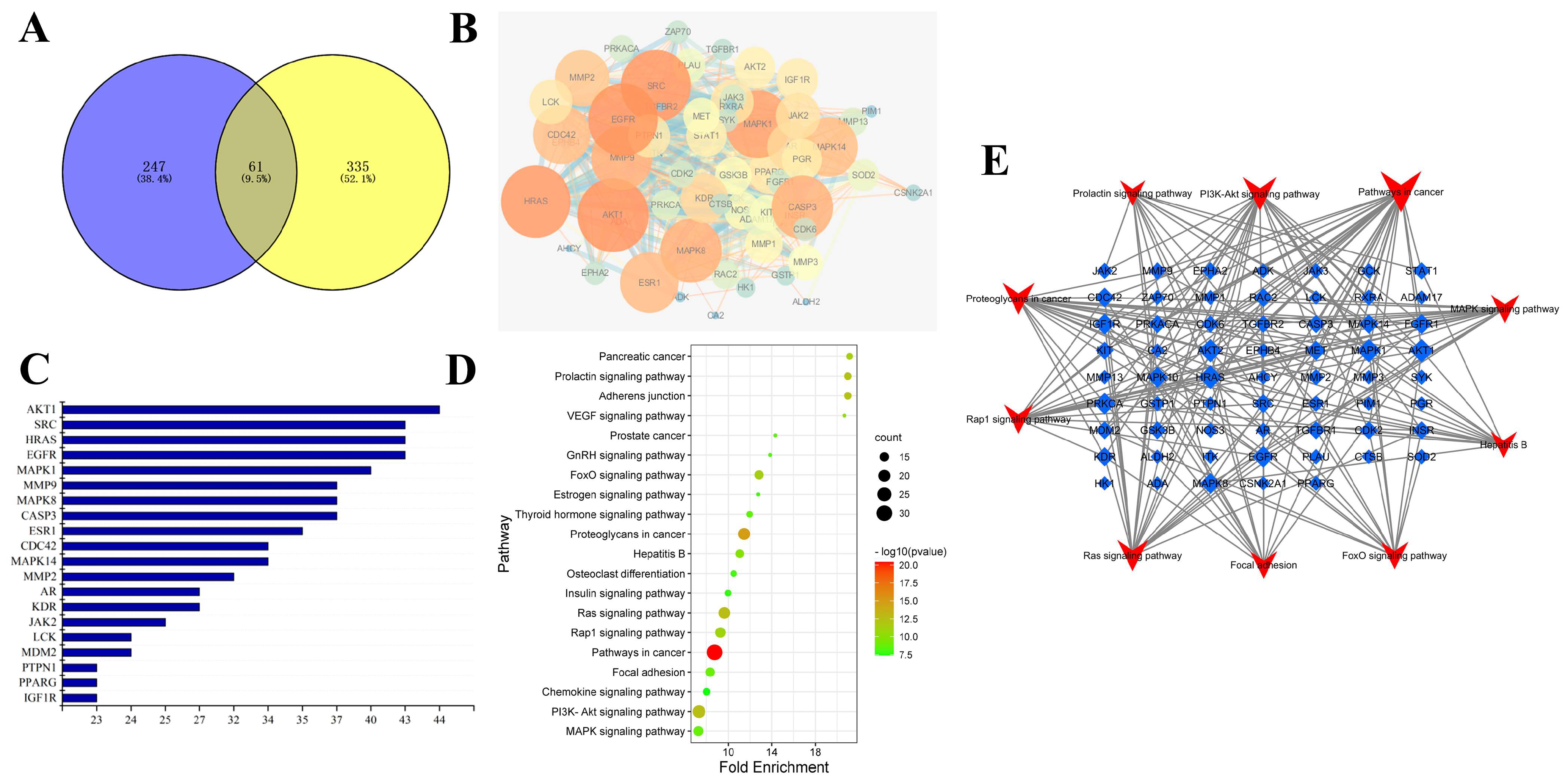
2.1. Prediction and Analysis of Potential Targets for Rosavin and ALI
2.2. In Vitro Results
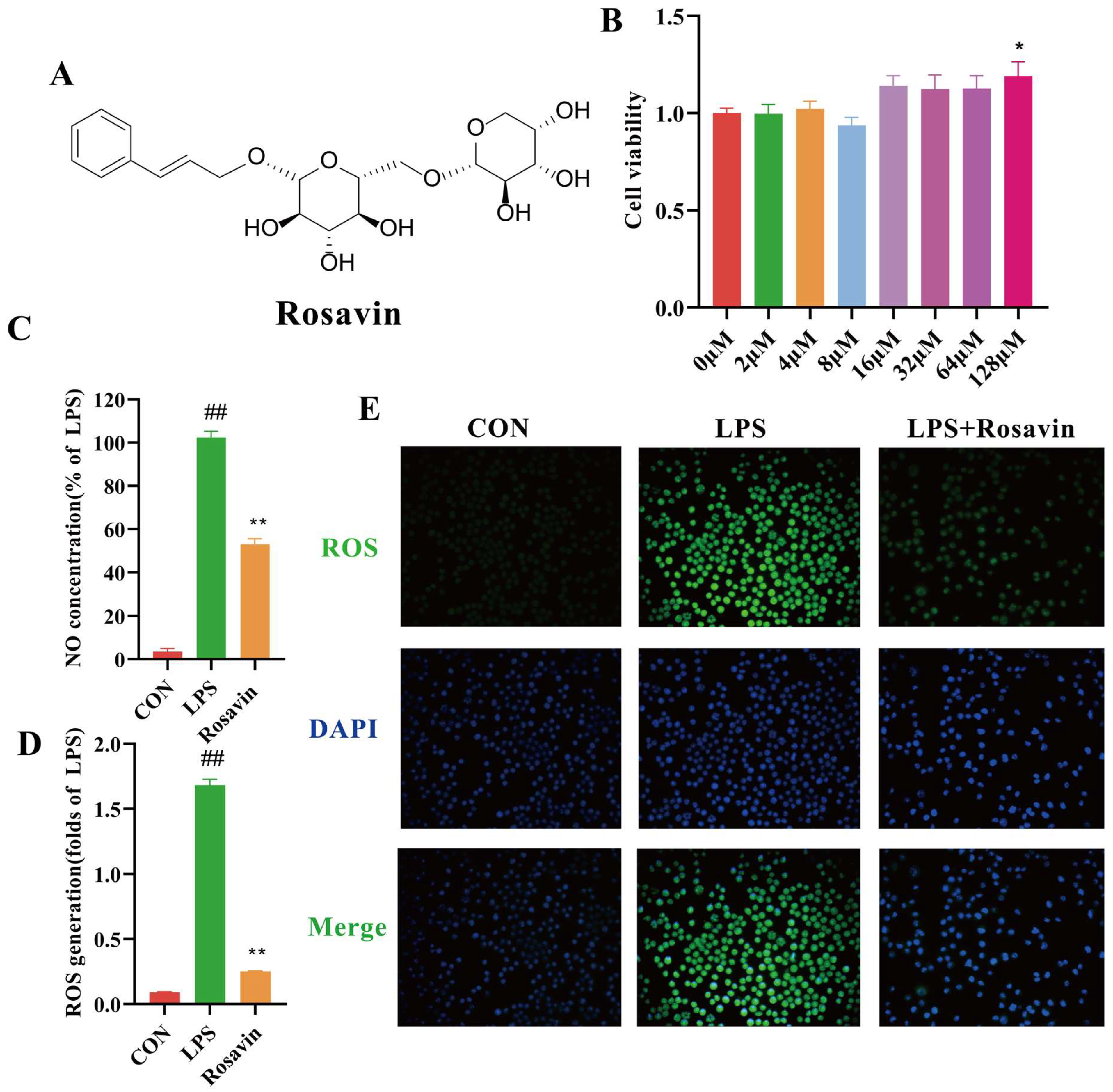
2.2.1. Rosavin Inhibits Pro-Inflammatory Mediator Release and Oxidative Stress Generated by LPS in RAW264.7 Cells
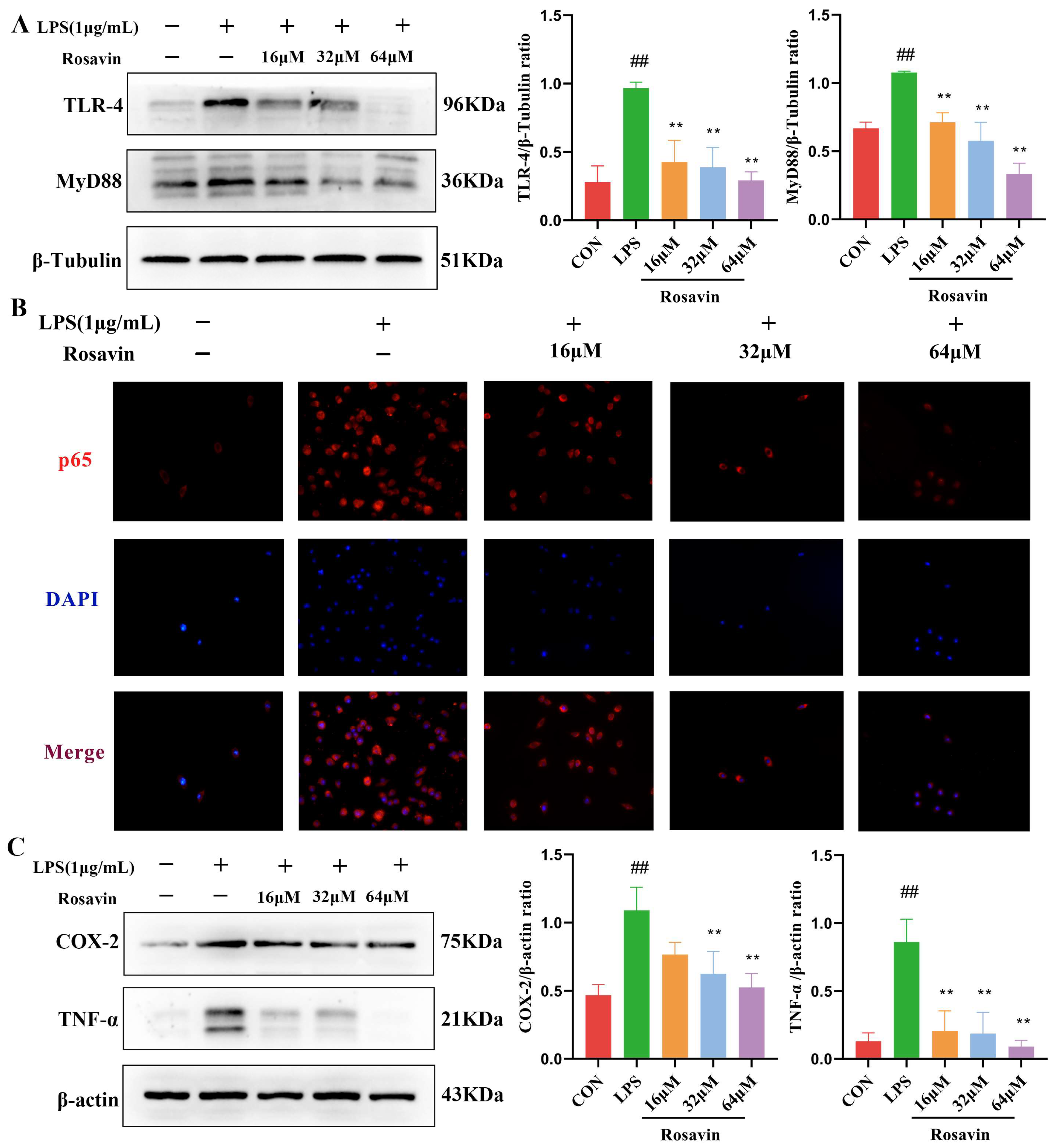
2.2.2. Rosavin Inhibits the TLR-4/NF-κB Signaling Cascade Generated by LPS in RAW264.7 Cells
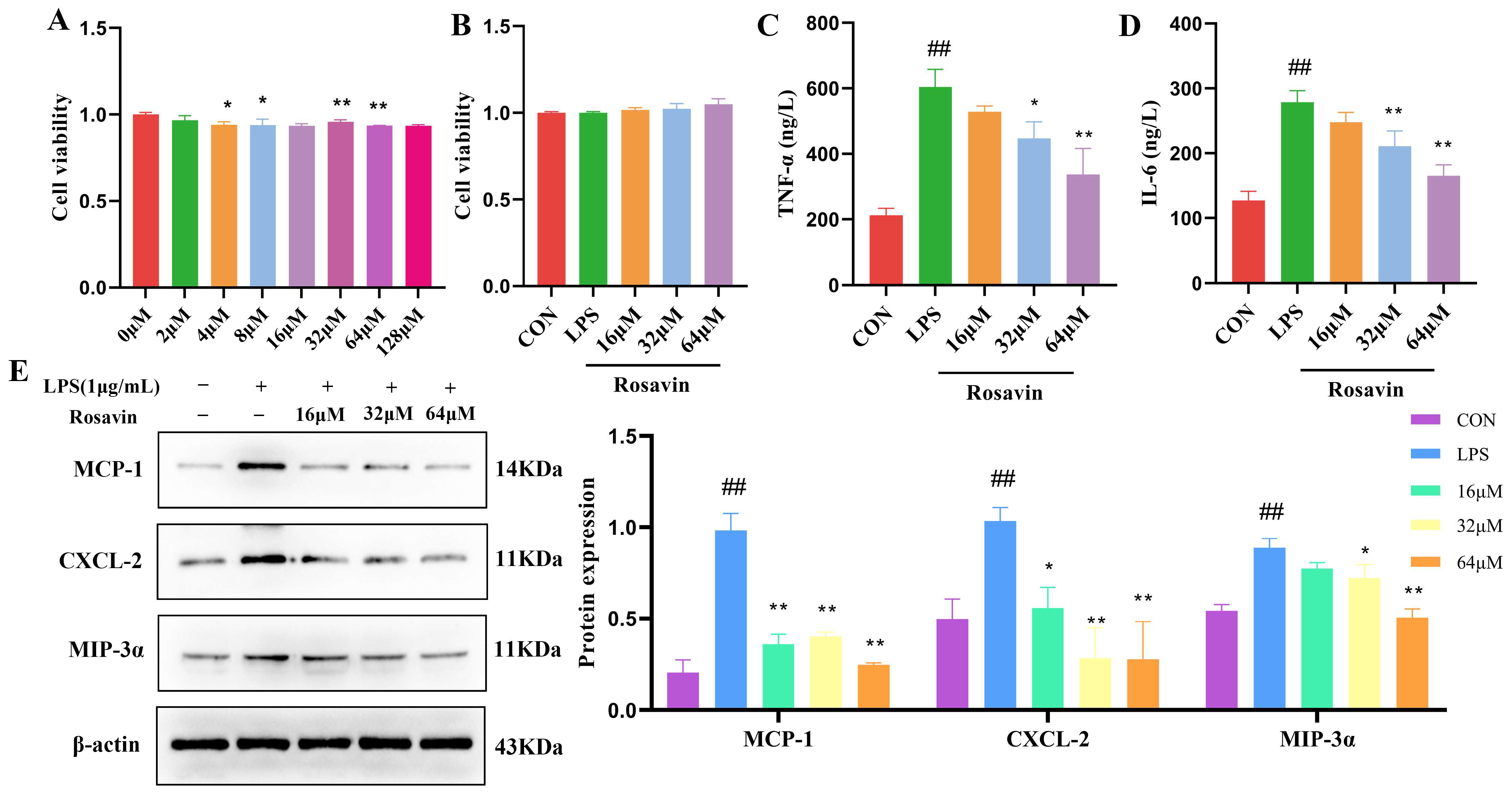
2.2.3. Rosavin Attenuated the Inflammation Response Cascade of LPS-Induced in A549 Cells
2.2.4. Rosavin Inhibits LPS-Induced Activation of the MAPK Signaling Pathway in A549 Cells
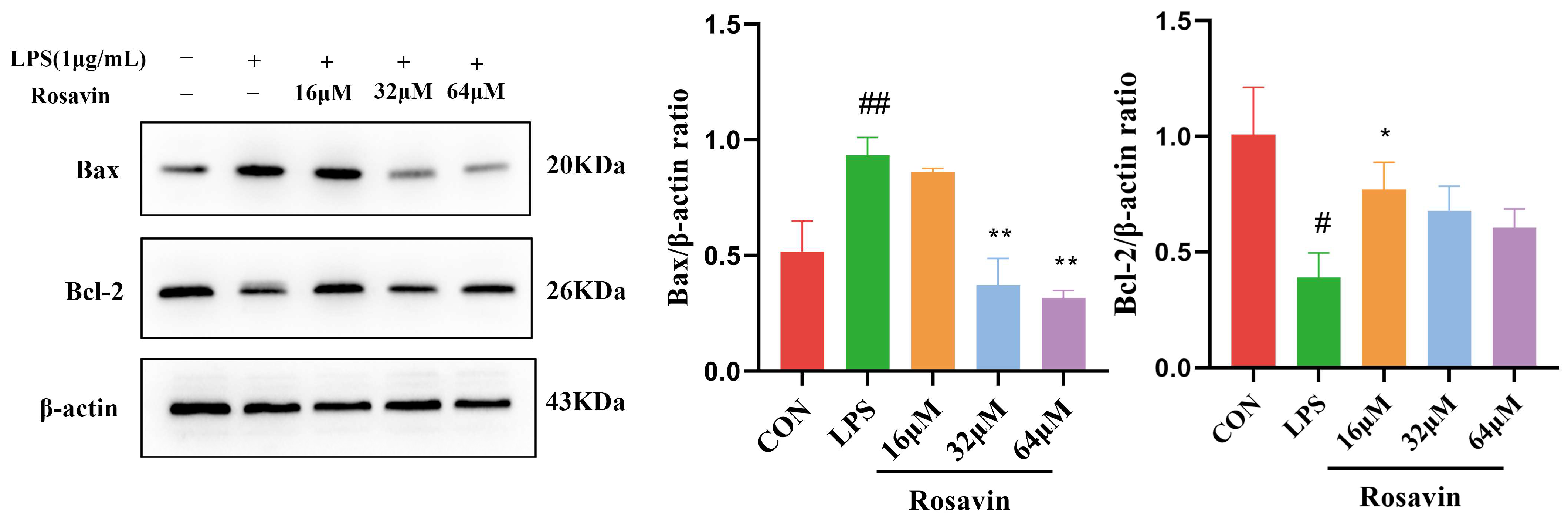
2.2.5. Rosavin Suppresses LPS-Induced Apoptosis in A549 Cells
2.3. In Vivo Results
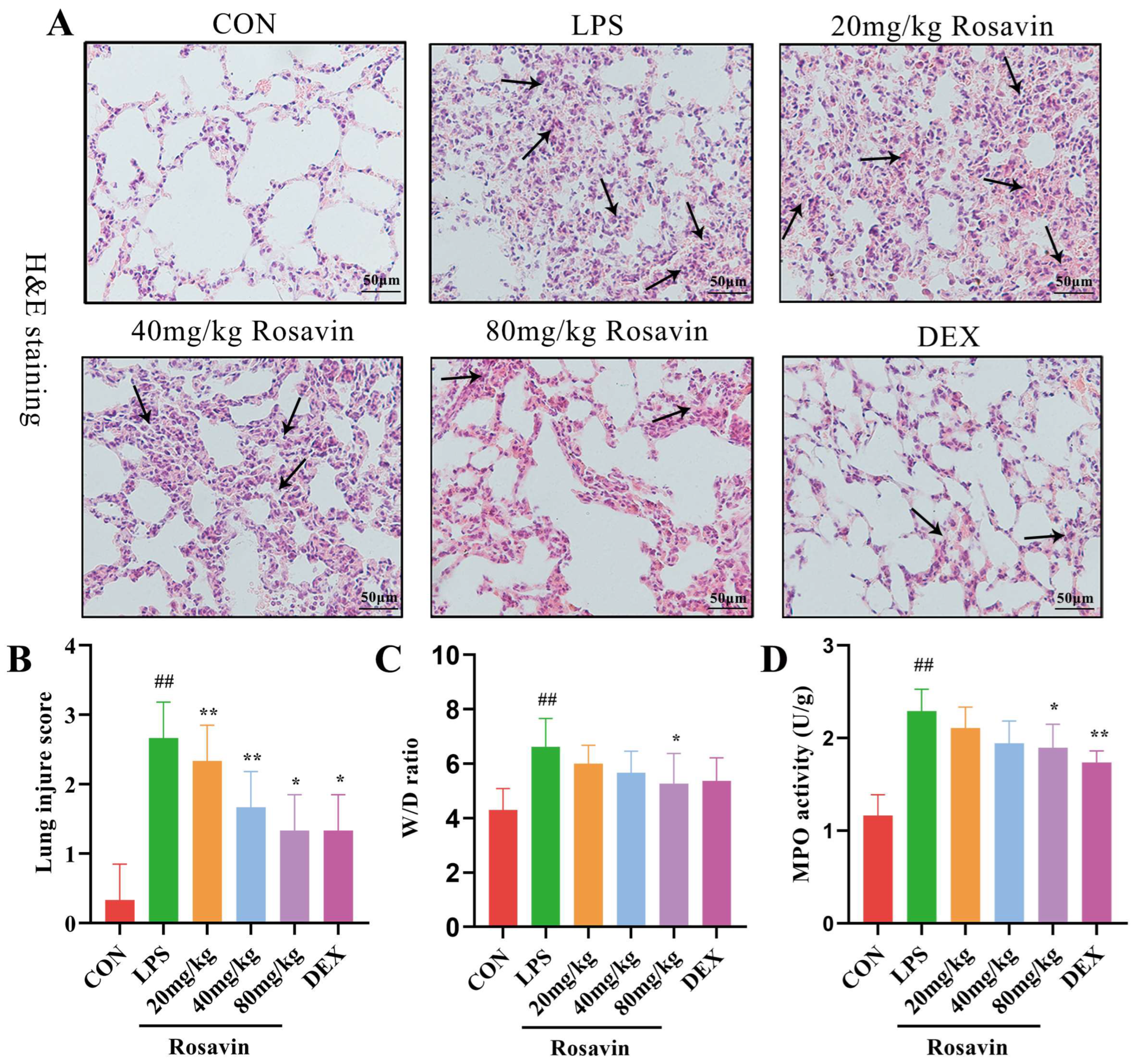
2.3.1. Rosavin Protects Mice from ALI Induced by LPS
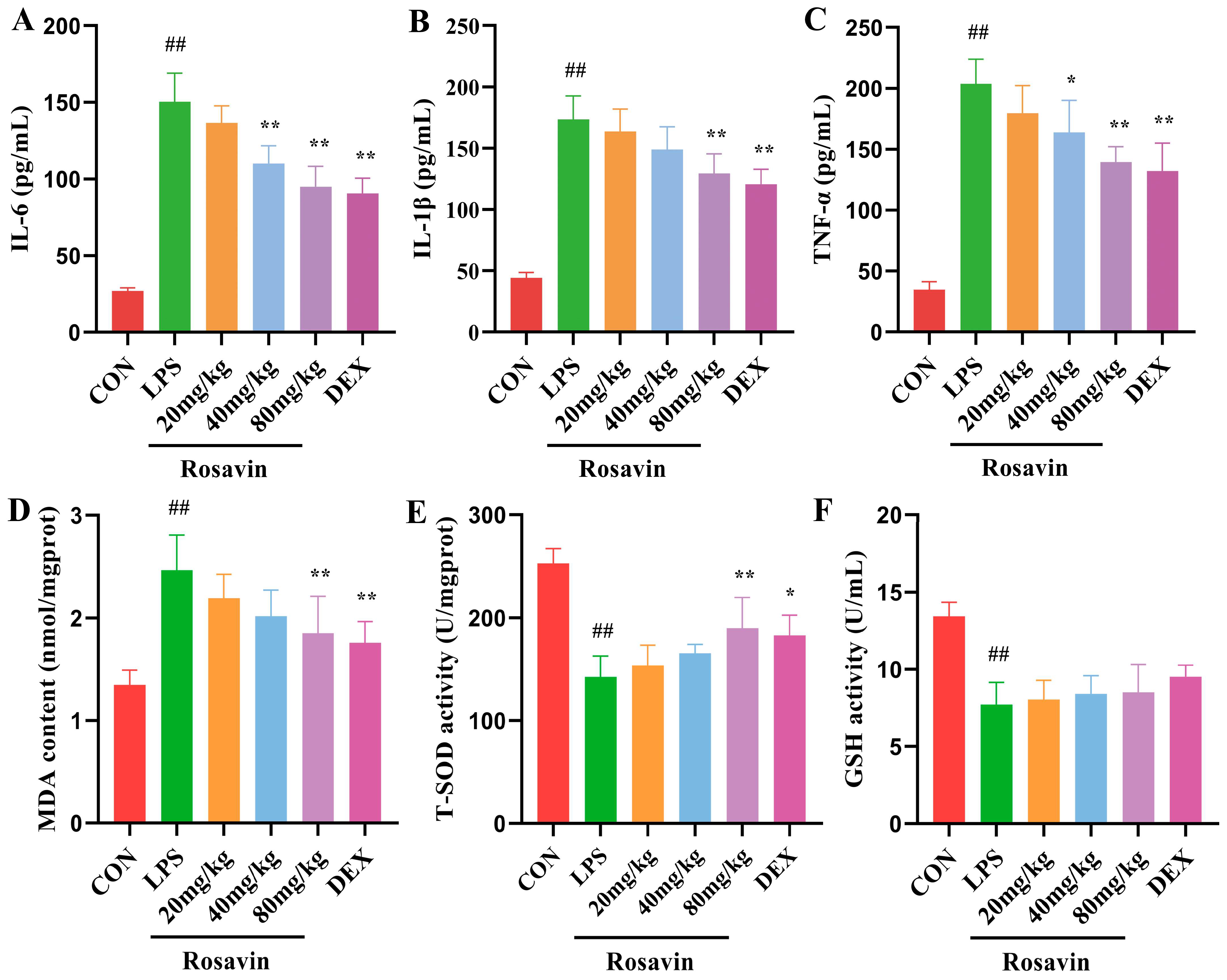
2.3.2. Rosavin Ameliorates LPS-Induced Release of Inflammatory Mediators and Oxidative Stress Injury in Mice
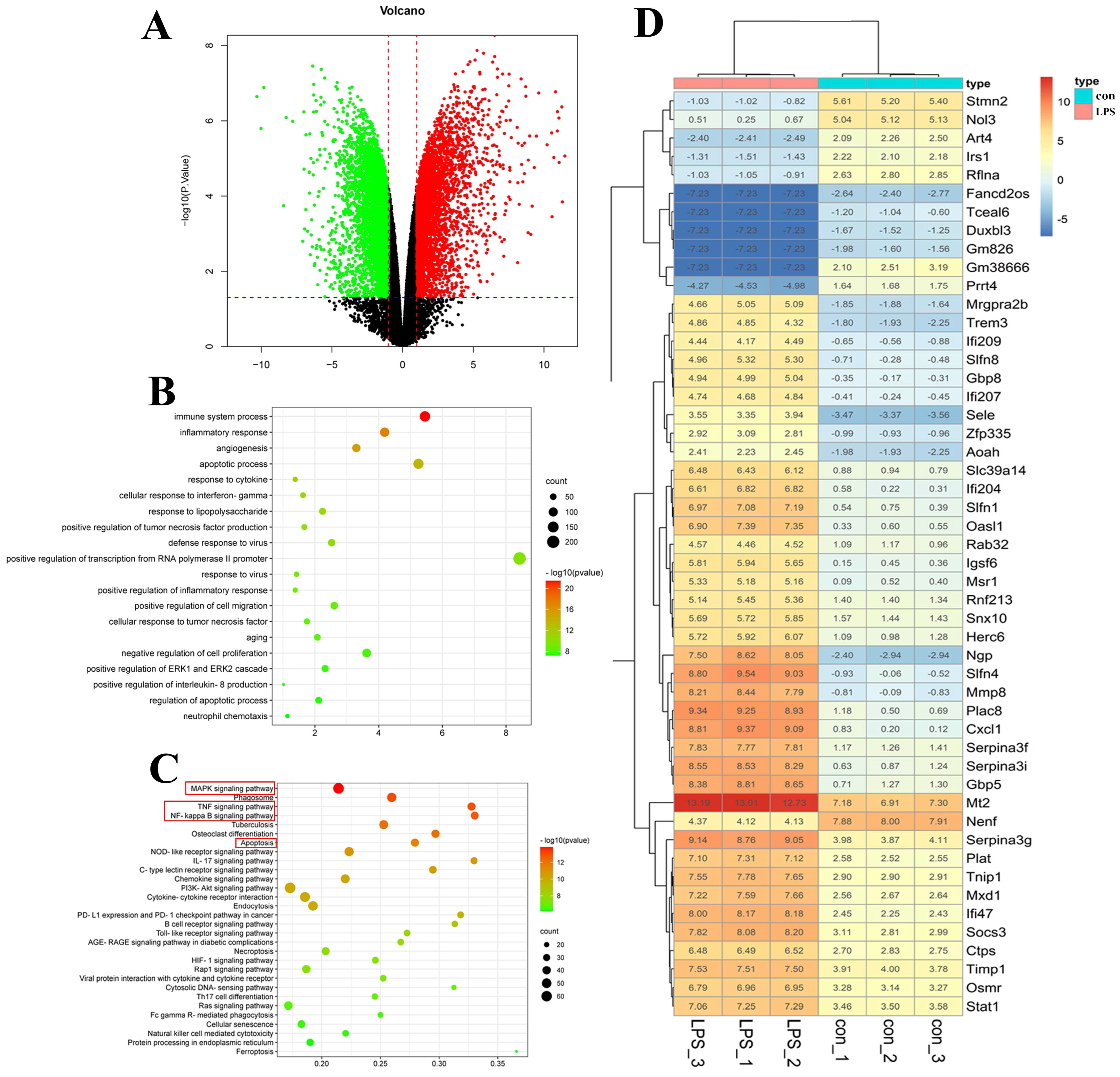
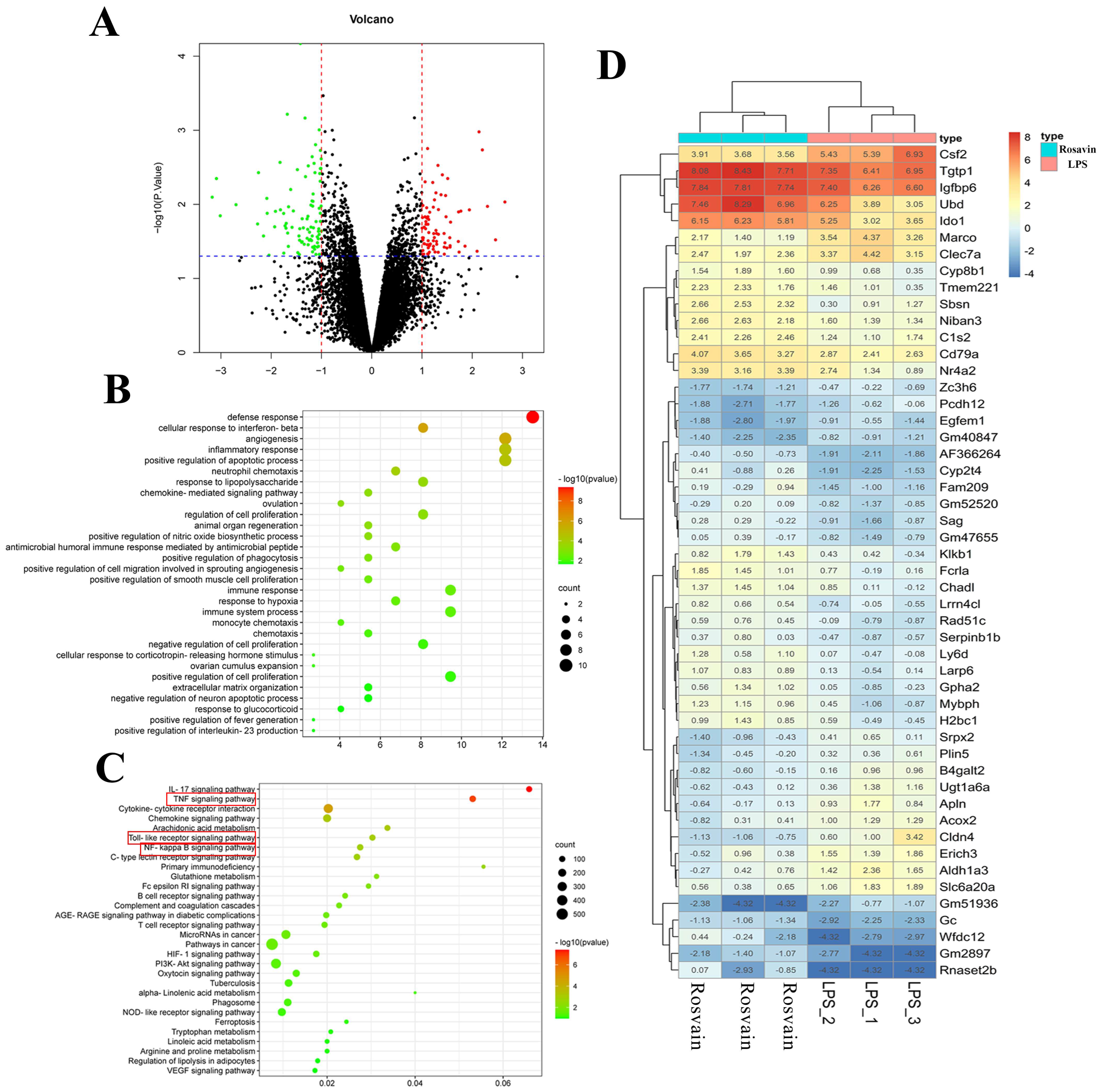
2.3.3. Transcriptomic Analysis
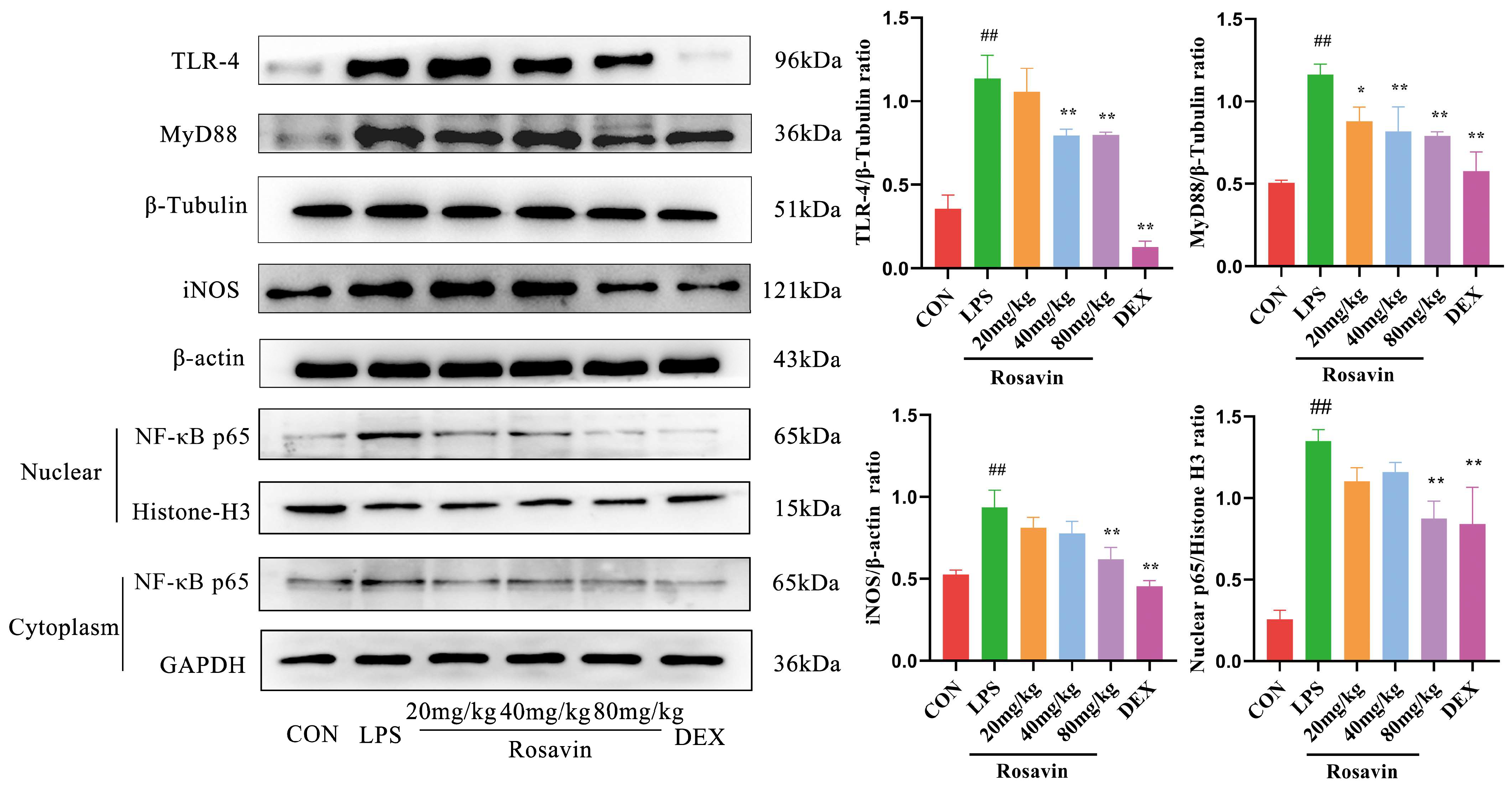
2.3.4. Rosavin Inhibits LPS-Induced Stimulation of the TLR-4/NF-κB Signaling Pathway in Mice
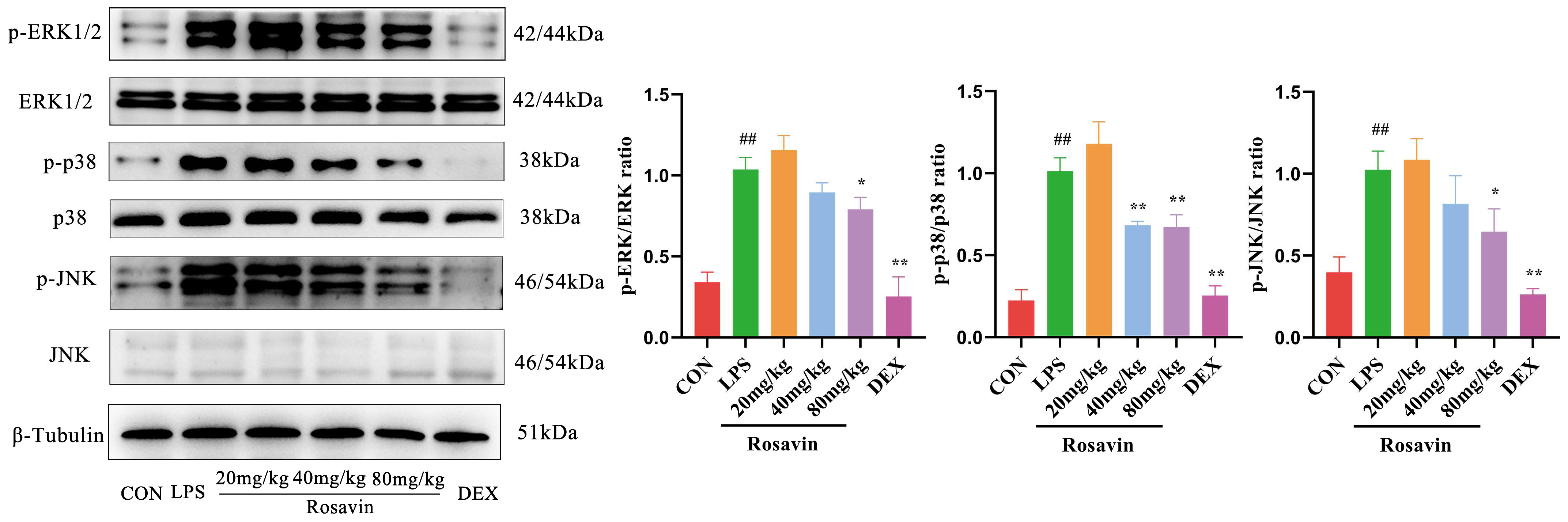
2.3.5. Rosavin Inhibits LPS-Induced Activation of the MAPK Signaling Pathway in Mice
2.3.6. Rosavin Suppresses LPS-Induced Apoptosis in Mice
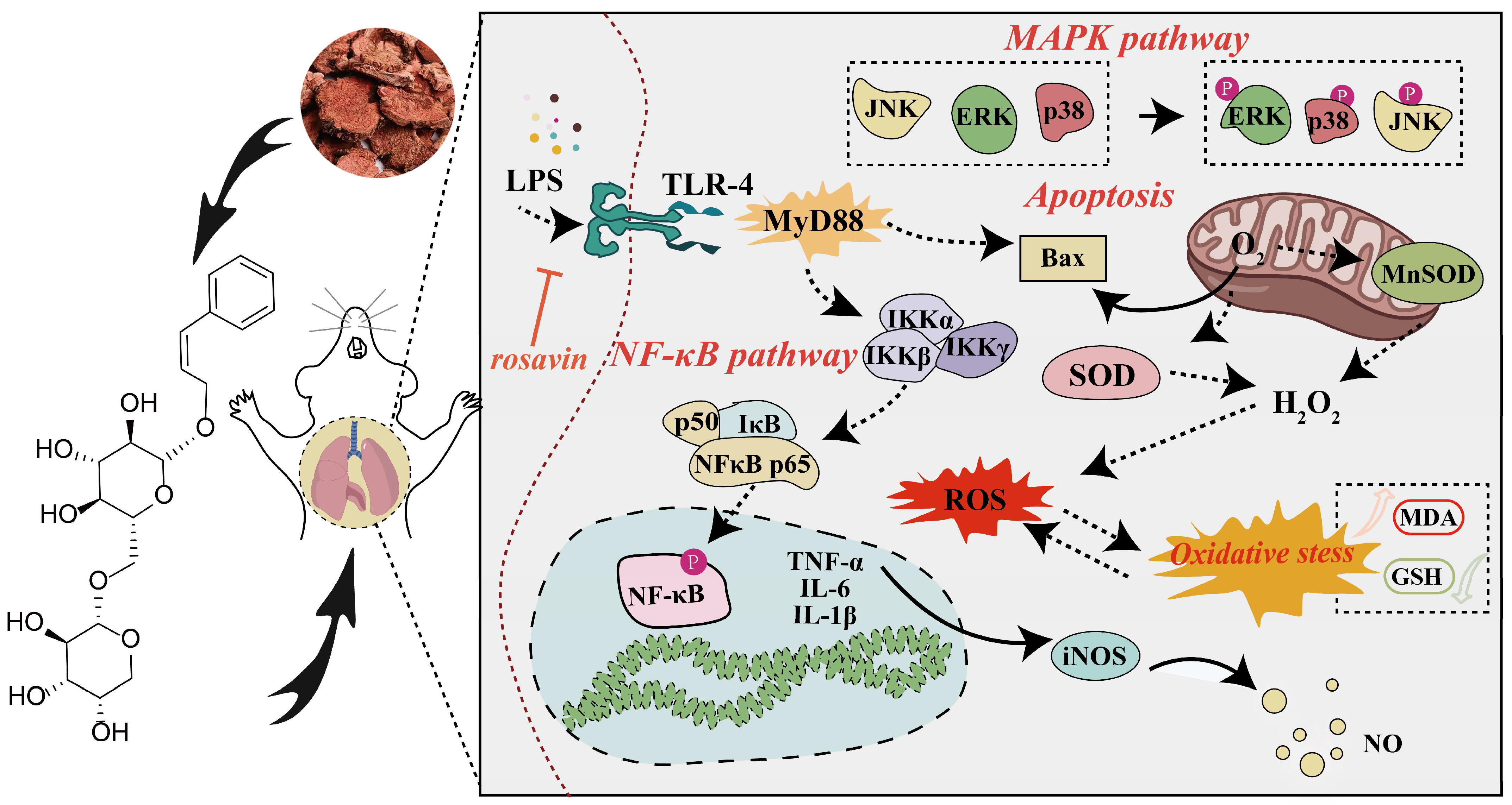
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Network Pharmacology Analysis
4.3. In Vitro Study
4.3.1. Cell Culture and Treatment
4.3.2. MTT Assay
4.3.3. Nitric Oxide Detection
4.3.4. Dectection of ROS
4.3.5. Cytokines’ Assay
4.3.6. Immunofluorescence
4.4. In Vivo Study
4.4.1. Animal Groups and Treatment
4.4.2. BALF Preparation and Examination
4.4.3. Lung Wet/Dry Weight Proportion Analysis
4.4.4. Histopathological Analysis
4.4.5. Enzyme-Linked Immunosorbent (ELISA) Assay
4.4.6. Determination of Myeloperoxidase (MPO), Superoxide Dismutase (SOD), Malondialdehyde (MDA) and Glutathione (GSH) in Lung Tissue
4.4.7. Western Blotting
4.4.8. Nucleocytoplasmic Separation
4.4.9. Transcriptome Sequencing
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ding, Z.; Zhong, R.; Yang, Y.; Xia, T.; Wang, W.; Wang, Y.; Xing, N.; Luo, Y.; Li, S.; Shang, L.; et al. Systems pharmacology reveals the mechanism of activity of Ge-Gen-Qin-Lian decoction against LPS-induced acute lung injury: A novel strategy for exploring active components and effective mechanism of TCM formulae. Pharmacol. Res. 2020, 156, 104759. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Deng, J.-S.; Huang, W.-C.; Jiang, W.-P.; Huang, G.-J. Attenuation of Lipopolysaccharide-Induced Acute Lung Injury by Hispolon in Mice, Through Regulating the TLR4/PI3K/Akt/mTOR and Keap1/Nrf2/HO-1 Pathways, and Suppressing Oxidative Stress-Mediated ER Stress-Induced Apoptosis and Autophagy. Nutrients 2020, 12, 1742. [Google Scholar] [CrossRef]
- Qian, Y.; Wang, Z.; Lin, H.; Lei, T.; Zhou, Z.; Huang, W.; Wu, X.; Zuo, L.; Wu, J.; Liu, Y.; et al. TRIM47 is a novel endothelial activation factor that aggravates lipopolysaccharide-induced acute lung injury in mice via K63-linked ubiquitination of TRAF2. Signal Transduct. Target. Ther. 2022, 7, 148. [Google Scholar] [CrossRef]
- Zhao, R.; Wang, B.; Wang, D.; Wu, B.; Ji, P.; Tan, D. Oxyberberine Prevented Lipopolysaccharide-Induced Acute Lung Injury through Inhibition of Mitophagy. Oxidative Med. Cell. Longev. 2021, 2021, 6675264. [Google Scholar] [CrossRef]
- Long, M.E.; Mallampalli, R.K.; Horowitz, J.C. Pathogenesis of pneumonia and acute lung injury. Clin. Sci. 2022, 136, 747–769. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Zhang, W.-H.; Zhu, Q.-M.; Ning, J.; Huo, X.-K.; Xiao, H.-T.; Sun, C.-P. Total terpenoids of Inula japonica activated the Nrf2 receptor to alleviate the inflammation and oxidative stress in LPS-induced acute lung injury. Phytomedicine 2022, 107, 154377. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, J.; Luo, Y.; Li, J.; Shang, L.; Zhou, F.; Yang, S. Honokiol alleviates LPS-induced acute lung injury by inhibiting NLRP3 inflammasome-mediated pyroptosis via Nrf2 activation in vitro and in vivo. Chin. Med. 2021, 16, 127. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Song, C.; Liu, Y.; Qu, J.; Li, H.; Xiao, W.; Kong, L.; Ge, H.; Sun, Y.; Lv, W. Re-Du-Ning injection ameliorates LPS-induced lung injury through inhibiting neutrophil extracellular traps formation. Phytomedicine 2021, 90, 153635. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; Li, Y.; Xue, Z.; Shao, R.; Li, L.; Zhu, Y.; Zhang, H.; Yang, J. Xuanfei Baidu Decoction reduces acute lung injury by regulating infiltration of neutrophils and macrophages via PD-1/IL17A pathway. Pharmacol. Res. 2022, 176, 106083. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, M.; Huo, X.-K.; Ning, J.; Yu, Z.-L.; Morisseau, C.; Sun, C.-P.; Hammock, B.D.; Ma, X.-C. Macrophage Inactivation by Small Molecule Wedelolactone via Targeting sEH for the Treatment of LPS-Induced Acute Lung Injury. ACS Cent. Sci. 2023, 9, 440–456. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yin, Z.; Feng, T.; Zhang, M.; Zhou, Z.; Zhou, Y. An integrated network pharmacology and RNA-Seq approach for exploring the preventive effect of Lonicerae japonicae flos on LPS-induced acute lung injury. J. Ethnopharmacol. 2021, 264, 113364. [Google Scholar] [CrossRef]
- Zhang, H.; Lang, W.; Wang, S.; Li, B.; Li, G.; Shi, Q. Echinacea polysaccharide alleviates LPS-induced lung injury via inhibiting inflammation, apoptosis and activation of the TLR4/NF-κB signal pathway. Int. Immunopharmacol. 2020, 88, 106974. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, Y.; Mak, M.; Tao, Z. Translational medicine for acute lung injury. J. Transl. Med. 2024, 22, 25. [Google Scholar] [CrossRef]
- Matute-Bello, G.; Downey, G.; Moore, B.B.; Groshong, S.D.; Matthay, M.A.; Slutsky, A.S.; Kuebler, W.M. An Official American Thoracic Society Workshop Report: Features and Measurements of Experimental Acute Lung Injury in Animals. Am. J. Respir. Cell Mol. Biol. 2011, 44, 725–738. [Google Scholar] [CrossRef]
- Wang, Y.-W.; Wu, Y.-H.; Zhang, J.-Z.; Tang, J.-H.; Fan, R.-P.; Li, F.; Yu, B.-Y.; Kou, J.-P.; Zhang, Y.-Y. Ruscogenin attenuates particulate matter-induced acute lung injury in mice via protecting pulmonary endothelial barrier and inhibiting TLR4 signaling pathway. Acta Pharmacol. Sin. 2020, 42, 726–734. [Google Scholar] [CrossRef]
- Miao, J.; Shen, J.; Yan, C.; Ren, J.; Liu, H.; Qiao, Y.; Li, Q. The protective effects of Mai-Luo-Ning injection against LPS-induced acute lung injury via the TLR4/NF-κB signalling pathway. Phytomedicine 2022, 104, 154290. [Google Scholar] [CrossRef]
- Cai, J.; Wang, Y.-l.; Sheng, X.-d.; Zhang, L.; Lv, X. Shufeng Jiedu capsule inhibits inflammation and apoptosis by activating A2AAR and inhibiting NF-κB to alleviate LPS-induced ALI. J. Ethnopharmacol. 2022, 298, 115661. [Google Scholar] [CrossRef]
- Hong, H.; Lou, S.; Zheng, F.; Gao, H.; Wang, N.; Tian, S.; Huang, G.; Zhao, H. Hydnocarpin D attenuates lipopolysaccharide-induced acute lung injury via MAPK/NF-κB and Keap1/Nrf2/HO-1 pathway. Phytomedicine 2022, 101, 154143. [Google Scholar] [CrossRef]
- Zhu, W.; Wang, M.; Jin, L.; Yang, B.; Bai, B.; Mutsinze, R.N.; Zuo, W.; Chattipakorn, N.; Huh, J.Y.; Liang, G.; et al. Licochalcone A protects against LPS-induced inflammation and acute lung injury by directly binding with myeloid differentiation factor 2 (MD2). Br. J. Pharmacol. 2022, 180, 1114–1131. [Google Scholar] [CrossRef]
- Shi, K.; Xiao, Y.; Dong, Y.; Wang, D.; Xie, Y.; Tu, J.; Xu, K.; Zhou, Z.; Cao, G.; Liu, Y. Protective Effects of Atractylodis lancea Rhizoma on Lipopolysaccharide-Induced Acute Lung Injury via TLR4/NF-κB and Keap1/Nrf2 Signaling Pathways In Vitro and In Vivo. Int. J. Mol. Sci. 2022, 23, 6134. [Google Scholar] [CrossRef]
- He, Y.-Q.; Zhou, C.-C.; Yu, L.-Y.; Wang, L.; Deng, J.-L.; Tao, Y.-L.; Zhang, F.; Chen, W.-S. Natural product derived phytochemicals in managing acute lung injury by multiple mechanisms. Pharmacol. Res. 2021, 163, 105224. [Google Scholar] [CrossRef]
- Li, C.; Huang, Y.; Yao, X.; Hu, B.; Wu, S.; Chen, G.; Lv, X.; Tian, F. Lugrandoside attenuates LPS-induced acute respiratory distress syndrome by anti-inflammation and anti-apoptosis in mice. Am. J. Transl. Res. 2016, 8, 5557–5568. [Google Scholar]
- Qi, Z.; Qi, S.; Ling, L.; Lv, J.; Feng, Z. Salidroside attenuates inflammatory response via suppressing JAK2-STAT3 pathway activation and preventing STAT3 transfer into nucleus. Int. Immunopharmacol. 2016, 35, 265–271. [Google Scholar] [CrossRef]
- Zhang, H.; Dong, W.; Li, S.; Zhang, Y.; Lv, Z.; Yang, L.; Jiang, L.; Wu, T.; Wang, Y. Salidroside protects against ventilation-induced lung injury by inhibiting the expression of matrix metalloproteinase-9. Pharm. Biol. 2021, 59, 760–768. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Y.; Zhang, P.; Ruan, W.; Zhang, L.; Yuan, S.; Pang, T.; Jia, A.-Q. Smiglaside A ameliorates LPS-induced acute lung injury by modulating macrophage polarization via AMPK-PPARγ pathway. Biochem. Pharmacol. 2018, 156, 385–395. [Google Scholar] [CrossRef]
- Gao, T.; Li, J.; Shi, L.; Hu, B. Rosavin inhibits neutrophil extracellular traps formation to ameliorate sepsis-induced lung injury by regulating the MAPK pathway. Allergol. Et Immunopathol. 2023, 51, 46–54. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, S.; Jia, N.; Shen, Z.; Huang, D.; Wang, X.; Wu, Y.; Pei, C.; Shi, S.; He, Y.; et al. Pretreatment with rosavin attenuates PM2.5-induced lung injury in rats through antiferroptosis via PI3K/Akt/Nrf2 signaling pathway. Phytother. Res. 2022, 37, 195–210. [Google Scholar] [CrossRef]
- Xin, X.; Yao, D.; Zhang, K.; Han, S.; Liu, D.; Wang, H.; Liu, X.; Li, G.; Huang, J.; Wang, J. Protective effects of Rosavin on bleomycin-induced pulmonary fibrosis via suppressing fibrotic and inflammatory signaling pathways in mice. Biomed. Pharmacother. 2019, 115, 108870. [Google Scholar] [CrossRef]
- Rao, Z.; Li, X.; Zhang, X.; Zeng, J.; Wang, B.; Yang, R.; Zeng, N. Fengreqing Oral Liquid Exerts Anti-Inflammatory Effects by Promoting Apoptosis and Inhibiting PI3K/AKT and NF-κB Signaling Pathways. Front. Pharmacol. 2022, 13, 824579. [Google Scholar] [CrossRef]
- Yu, Y.-Y.; Li, X.-Q.; Hu, W.-P.; Cu, S.-C.; Dai, J.-J.; Gao, Y.-N.; Zhang, Y.-T.; Bai, X.-Y.; Shi, D.-Y. Self-developed NF-κB inhibitor 270 protects against LPS-induced acute kidney injury and lung injury through improving inflammation. Biomed. Pharmacother. 2022, 147, 112615. [Google Scholar] [CrossRef]
- Wang, J.; Yang, H.; Zheng, D.; Sun, Y.; An, L.; Li, G.; Zhao, Z. Integrating network pharmacology and pharmacological evaluation to reveal the therapeutic effects and potential mechanism of S-allylmercapto-N-acetylcysteine on acute respiratory distress syndrome. Int. Immunopharmacol. 2023, 121, 110516. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Luo, W.; Han, J.; Zhang, Q.; Ji, L.; Samorodov, A.V.; Pavlov, V.N.; Zhuang, Z.; Yang, D.; Yin, L.; et al. Schisandrin B protects against LPS-induced inflammatory lung injury by targeting MyD88. Phytomedicine 2023, 108, 154489. [Google Scholar] [CrossRef]
- Han, S.; Yuan, R.; Cui, Y.; He, J.; Wang, Q.-Q.; Zhuo, Y.; Yang, S.; Gao, H. Hederasaponin C Alleviates Lipopolysaccharide-Induced Acute Lung Injury In Vivo and In Vitro Through the PIP2/NF-κB/NLRP3 Signaling Pathway. Front. Immunol. 2022, 13, 846384. [Google Scholar] [CrossRef]
- Jia, X.; Zhang, K.; Feng, S.; Li, Y.; Yao, D.; Liu, Q.; Liu, D.; Li, X.; Huang, J.; Wang, H.; et al. Total glycosides of Rhodiola rosea L. attenuate LPS-induced acute lung injury by inhibiting TLR4/NF-κB pathway. Biomed. Pharmacother. 2023, 158, 114186. [Google Scholar] [CrossRef]
- Yang, L.; Chen, H.; Hu, Q.; Liu, L.; Yuan, Y.; Zhang, C.; Tang, J.; Shen, X. Eupalinolide B attenuates lipopolysaccharide-induced acute lung injury through inhibition of NF-κB and MAPKs signaling by targeting TAK1 protein. Int. Immunopharmacol. 2022, 111, 109148. [Google Scholar] [CrossRef]
- Chen, J.; Huang, Y.; Bian, X.; He, Y. Berberine Ameliorates Inflammation in Acute Lung Injury via NF-κB/Nlrp3 Signaling Pathway. Front. Nutr. 2022, 9, 851255. [Google Scholar] [CrossRef]
- Liu, L.; Chen, X.; Jiang, Y.; Yuan, Y.; Yang, L.; Hu, Q.; Tang, J.; Meng, X.; Xie, C.; Shen, X. Brevilin A Ameliorates Acute Lung Injury and Inflammation Through Inhibition of NF-κB Signaling via Targeting IKKα/β. Front. Pharmacol. 2022, 13, 911157. [Google Scholar] [CrossRef]
- Liao, X.; Zhang, W.; Dai, H.; Jing, R.; Ye, M.; Ge, W.; Pei, S.; Pan, L. Neutrophil-Derived IL-17 Promotes Ventilator-Induced Lung Injury via p38 MAPK/MCP-1 Pathway Activation. Front. Immunol. 2021, 12, 768813. [Google Scholar] [CrossRef]
- Kim, S.-H.; Hong, J.-H.; Yang, W.-K.; Geum, J.-H.; Kim, H.-R.; Choi, S.-Y.; Kang, Y.-M.; An, H.-J.; Lee, Y.-C. Herbal Combinational Medication of Glycyrrhiza glabra, Agastache rugosa Containing Glycyrrhizic Acid, Tilianin Inhibits Neutrophilic Lung Inflammation by Affecting CXCL2, Interleukin-17/STAT3 Signal Pathways in a Murine Model of COPD. Nutrients 2020, 12, 926. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, M.; Xu, Y.; Liao, J.; Li, X.; Zhou, Y.; Dai, J.; Li, X.; Chen, P.; Chen, G.; et al. Discovery of 4-oxo-N-phenyl-1,4-dihydroquinoline-3-carboxamide derivatives as novel anti-inflammatory agents for the treatment of acute lung injury and sepsis. Eur. J. Med. Chem. 2023, 249, 115144. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.L.; Kim, J.M.; Go, M.J.; Kim, T.Y.; Joo, S.G.; Kim, J.H.; Lee, H.S.; Kim, H.-J.; Heo, H.J. Protective Effect of Lonicera japonica on PM2.5-Induced Pulmonary Damage in BALB/c Mice via the TGF-β and NF-κB Pathway. Antioxidants 2023, 12, 968. [Google Scholar] [CrossRef]
- Liao, J.; Yang, J.; Li, X.; Hu, C.; Zhu, W.; Zhou, Y.; Zou, Y.; Guo, M.; Chen, Z.; Li, X.; et al. Discovery of the Diphenyl 6-Oxo-1,6-dihydropyridazine-3-carboxylate/carboxamide Analogue J27 for the Treatment of Acute Lung Injury and Sepsis by Targeting JNK2 and Inhibiting the JNK2-NF-κB/MAPK Pathway. J. Med. Chem. 2023, 66, 12304–12323. [Google Scholar] [CrossRef]
- Cui, B.; Liu, Y.; Chen, J.; Chen, H.; Feng, Y.; Zhang, P. Small molecule inhibitor CRT0066101 inhibits cytokine storm syndrome in a mouse model of lung injury. Int. Immunopharmacol. 2023, 120, 110240. [Google Scholar] [CrossRef]
- Liang, J.; Liu, J.; Tang, Y.; Peng, Q.; Zhang, L.; Ma, X.; Xu, N.; Wei, J.; Han, H. Sophoridine inhibits endotoxin-induced acute lung injury by enhancing autophagy of macrophage and reducing inflammation. J. Leukoc. Biol. 2022, 112, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhao, Y.; Wang, X.; Lin, Y.; Zhao, W.; Wu, D.; Pan, J.; Luo, W.; Wang, Y.; Liang, G. FAK mediates LPS-induced inflammatory lung injury through interacting TAK1 and activating TAK1-NFκB pathway. Cell Death Dis. 2022, 13, 589. [Google Scholar] [CrossRef]
- Jangam, A.; Tirunavalli, S.K.; Adimoolam, B.M.; Kasireddy, B.; Patnaik, S.S.; Erukkambattu, J.; Thota, J.R.; Andugulapati, S.B.; Addlagatta, A. Anti-inflammatory and antioxidant activities of Gymnema Sylvestre extract rescue acute respiratory distress syndrome in rats via modulating the NF-κB/MAPK pathway. Inflammopharmacology 2023, 31, 823–844. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Bai, B.; Ge, X.; Zheng, B.; Xiao, Z.; Tang, Y.; Fang, L.; Tang, Y.; Dai, Y.; Zhang, B.; et al. Costunolide attenuates LPS-induced inflammation and lung injury through inhibiting IKK/NF-κB signaling. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2023. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Z.; Li, P.; Gao, Y.; Shi, Y. Bakuchiol regulates TLR4/MyD88/NF-κB and Keap1/Nrf2/HO-1 pathways to protect against LPS-induced acute lung injury in vitro and in vivo. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2023. [Google Scholar] [CrossRef]
- Li, Y.-L.; Qin, S.-Y.; Li, Q.; Song, S.-J.; Xiao, W.; Yao, G.-D. Jinzhen Oral Liquid alleviates lipopolysaccharide-induced acute lung injury through modulating TLR4/MyD88/NF-κB pathway. Phytomedicine 2023, 114, 154744. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, H.; Lv, Z.; Du, Y.; Li, D.; Fang, H.; You, J.; Yu, L.; Li, R. Up-regulated CD38 by daphnetin alleviates lipopolysaccharide-induced lung injury via inhibiting MAPK/NF-κB/NLRP3 pathway. Cell Commun. Signal. 2023, 21, 66. [Google Scholar] [CrossRef] [PubMed]
- Pei, X.; Zhang, Z.; Wang, N.; Huang, G.; Min, X.; Yang, Y.; Cao, J. Onychiol B attenuates lipopolysaccharide-induced inflammation via MAPK/NF-κB pathways and acute lung injury in vivo. Bioorg. Chem. 2023, 132, 106351. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.-R.; Qu, F.; Zhong, W.-J.; Yang, H.-H.; Zeng, J.; Huang, J.-H.; Liu, J.; Zhang, M.-Y.; Zhou, Y.; Guan, C.-X. Beneficial effects of aloperine on inflammation and oxidative stress by suppressing necroptosis in lipopolysaccharide-induced acute lung injury mouse model. Phytomedicine 2022, 100, 154074. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.J.; Zhang, M.L.; Hou, Y.M.; Zhang, K.; Yao, D.H.; Li, G.Y.; Kou, W.B.; Wang, H.Y.; Wang, J.H. The Amomum tsao-ko Essential Oils Inhibited Inflammation and Apoptosis through p38/JNK MAPK Signaling Pathway and Alleviated Gentamicin-Induced Acute Kidney Injury. Molecules 2022, 27, 7121. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, D.; Chen, Y.; Abudou, H.; Wang, H.; Cai, J.; Wang, Y.; Liu, Z.; Liu, Y.; Fan, H. Classic Signaling Pathways in Alveolar Injury and Repair Involved in Sepsis-Induced ALI/ARDS: New Research Progress and Prospect. Dis. Markers 2022, 2022, 6362344. [Google Scholar] [CrossRef] [PubMed]
- Bos, L.D.J.; Ware, L.B. Acute respiratory distress syndrome: Causes, pathophysiology, and phenotypes. Lancet 2022, 400, 1145–1156. [Google Scholar] [CrossRef]
- Wang, J.; Ren, C.; Bi, W.; Batu, W. Glycyrrhizin mitigates acute lung injury by inhibiting the NLRP3 inflammasome in vitro and in vivo. J. Ethnopharmacol. 2023, 303, 115948. [Google Scholar] [CrossRef]
- Xi, X.; Yao, Y.; Liu, N.; Li, P. MiR-297 alleviates LPS-induced A549 cell and mice lung injury via targeting cyclin dependent kinase 8. Int. Immunopharmacol. 2020, 80, 106197. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.-H.; Zhang, K.; Feng, S.-S.; Zhang, L.-J.; Li, S.-Y.; Wang, H.-Y.; Wang, J.-H. Rosavin Alleviates LPS-Induced Acute Lung Injure by Modulating the TLR-4/NF-κB/MAPK Singnaling Pathways. Int. J. Mol. Sci. 2024, 25, 1875. https://doi.org/10.3390/ijms25031875
Liu Q-H, Zhang K, Feng S-S, Zhang L-J, Li S-Y, Wang H-Y, Wang J-H. Rosavin Alleviates LPS-Induced Acute Lung Injure by Modulating the TLR-4/NF-κB/MAPK Singnaling Pathways. International Journal of Molecular Sciences. 2024; 25(3):1875. https://doi.org/10.3390/ijms25031875
Chicago/Turabian StyleLiu, Qiao-Hui, Ke Zhang, Shu-Shu Feng, Li-Juan Zhang, Shun-Ying Li, Hang-Yu Wang, and Jin-Hui Wang. 2024. "Rosavin Alleviates LPS-Induced Acute Lung Injure by Modulating the TLR-4/NF-κB/MAPK Singnaling Pathways" International Journal of Molecular Sciences 25, no. 3: 1875. https://doi.org/10.3390/ijms25031875
APA StyleLiu, Q.-H., Zhang, K., Feng, S.-S., Zhang, L.-J., Li, S.-Y., Wang, H.-Y., & Wang, J.-H. (2024). Rosavin Alleviates LPS-Induced Acute Lung Injure by Modulating the TLR-4/NF-κB/MAPK Singnaling Pathways. International Journal of Molecular Sciences, 25(3), 1875. https://doi.org/10.3390/ijms25031875