

Searching for Frataxin Function: Exploring the Analogy with Nqo15, the Frataxin-like Protein of Respiratory Complex I from Thermus thermophilus

 , , , , , , ,
, , , , , , ,  , ,
, , 
Abstract
1. Introduction
2. Results
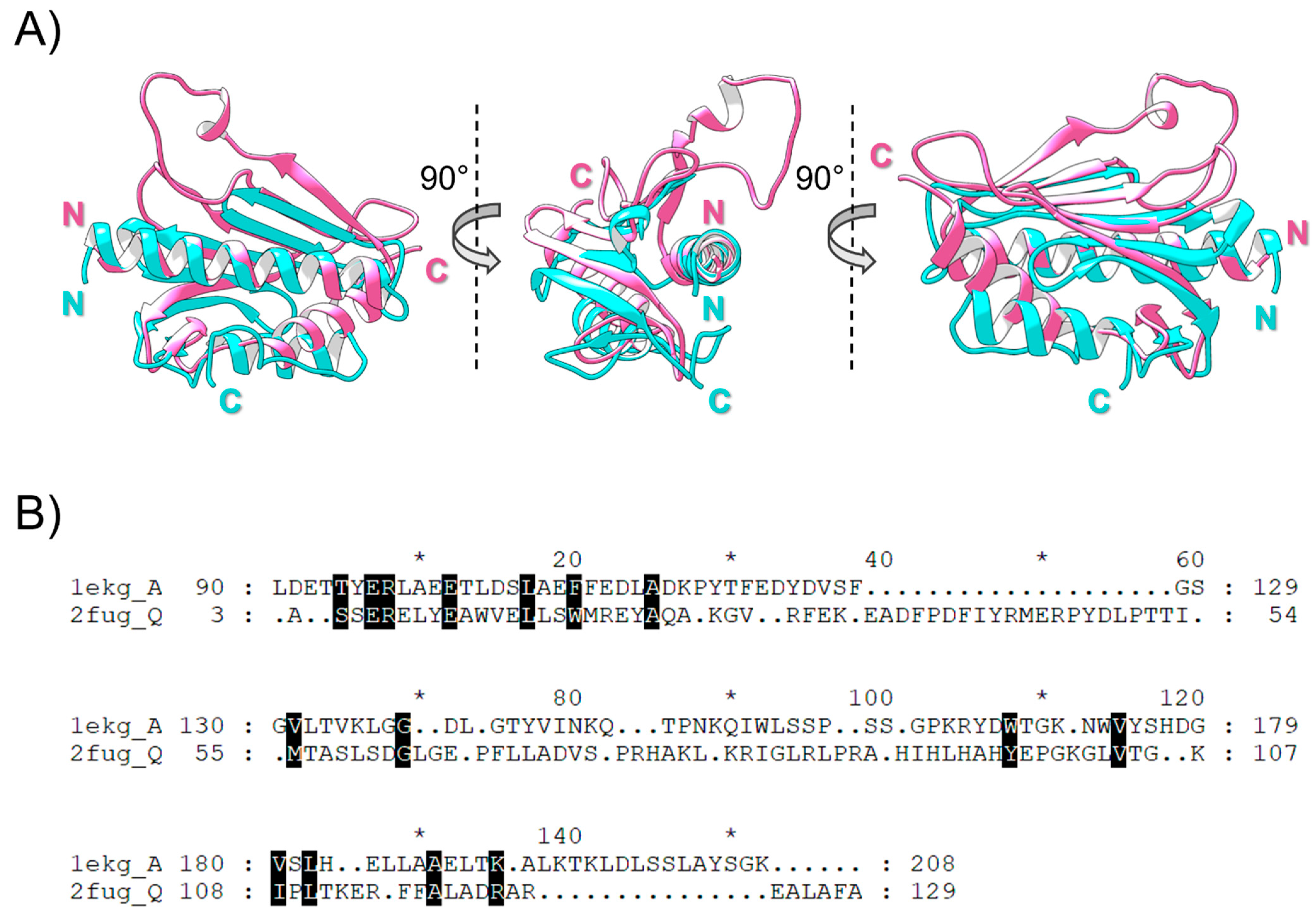
2.1. Searching for Nqo15 and FXN Homology
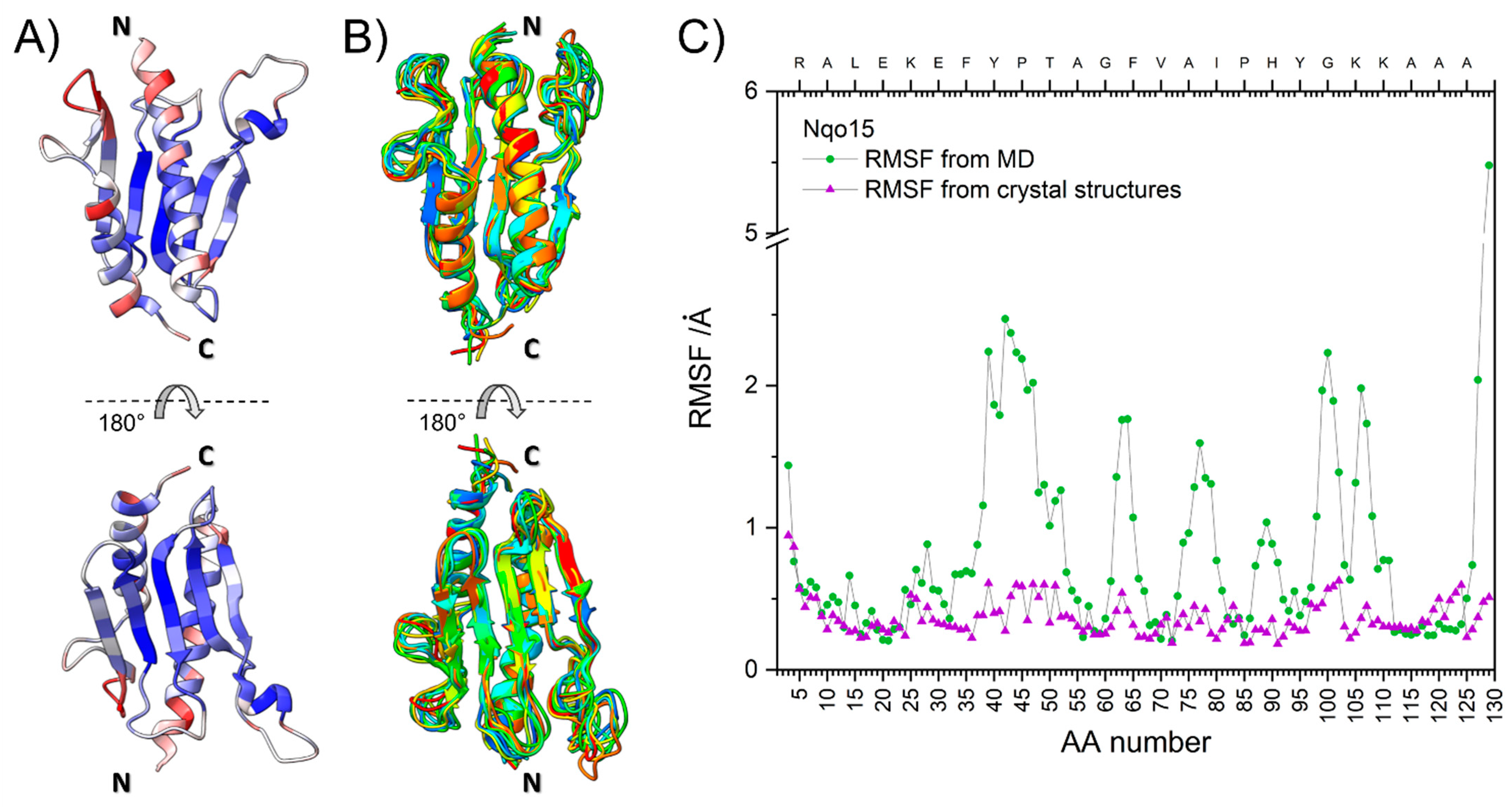
2.2. Structural Integrity, Solubility and Stability of Standalone Nqo15
2.3. Nqo15 Does Not Bind Iron
2.4. Nqo15 Does Not Activate Human Cysteine Desulfurase
3. Discussion
4. Materials and Methods
4.1. Protein Expression and Purification
- FwNqo15: 5′-GGAGATATACATATGAGCGCGTCTTCCGAGCGCGAACTC-3′;
- RevNqo15: 5′-GCAGCCGGATCCTCAGGCGAAGGCCAAAGCCTCCCGC-3′;
- FwpET-9b: 5′-TGAGGATCCGGCTGCTAACAAAG-3′;
- RevpET-9b: 5′-CATATGTATATCTCCTTCTTAAAG-3′.
4.2. Static Light Scattering (SLS) Measurements
4.3. Circular Dichroism (CD) Spectroscopy
4.4. Nuclear Magnetic Resonance (NMR) Spectroscopy
4.5. Fluorescence Spectroscopy
4.6. Iron (III)-Binding Capability Assay
4.7. Cysteine Desulfurase Activity Measurements
4.8. In Silico Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Campuzano, V.; Montermini, L.; Moltò, M.D.; Pianese, L.; Cossée, M.; Cavalcanti, F.; Monros, E.; Rodius, F.; Duclos, F.; Monticelli, A.; et al. Friedreich’s Ataxia: Autosomal Recessive Disease Caused by an Intronic GAA Triplet Repeat Expansion. Science 1996, 271, 1423–1427. [Google Scholar] [CrossRef]
- Dürr, A.; Cossee, M.; Agid, Y.; Campuzano, V.; Mignard, C.; Penet, C.; Mandel, J.-L.; Brice, A.; Koenig, M. Clinical and Genetic Abnormalities in Patients with Friedreich’s Ataxia. N. Engl. J. Med. 1996, 335, 1169–1175. [Google Scholar] [CrossRef]
- Pandolfo, M. Friedreich Ataxia: The Clinical Picture. J. Neurol. 2009, 256 (Suppl. 1), 3–8. [Google Scholar] [CrossRef]
- Payne, R.M.; Wagner, G.R. Cardiomyopathy in Friedreich Ataxia: Clinical Findings and Research. J. Child. Neurol. 2012, 27, 1179–1186. [Google Scholar] [CrossRef]
- Campuzano, V.; Montermini, L.; Lutz, Y.; Cova, L.; Hindelang, C.; Jiralerspong, S.; Trottier, Y.; Kish, S.J.; Faucheux, B.; Trouillas, P.; et al. Frataxin Is Reduced in Friedreich Ataxia Patients and Is Associated with Mitochondrial Membranes. Hum. Mol. Genet. 1997, 6, 1771–1780. [Google Scholar] [CrossRef]
- Cossée, M.; Dürr, A.; Schmitt, M.; Dahl, N.; Trouillas, P.; Allinson, P.; Kostrzewa, M.; Nivelon-Chevallier, A.; Gustavson, K.H.; Kohlschütter, A.; et al. Friedreich’s Ataxia: Point Mutations and Clinical Presentation of Compound Heterozygotes. Ann. Neurol. 1999, 45, 200–206. [Google Scholar] [CrossRef]
- Galea, C.A.; Huq, A.; Lockhart, P.J.; Tai, G.; Corben, L.A.; Yiu, E.M.; Gurrin, L.C.; Lynch, D.R.; Gelbard, S.; Durr, A.; et al. Compound Heterozygous FXN Mutations and Clinical Outcome in Friedreich Ataxia. Ann. Neurol. 2016, 79, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Puccio, H.; Simon, D.; Cossée, M.; Criqui-Filipe, P.; Tiziano, F.; Melki, J.; Hindelang, C.; Matyas, R.; Rustin, P.; Koenig, M. Mouse Models for Friedreich Ataxia Exhibit Cardiomyopathy, Sensory Nerve Defect and Fe-S Enzyme Deficiency Followed by Intramitochondrial Iron Deposits. Nat. Genet. 2001, 27, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Schulz, J.B.; Dehmer, T.; Schöls, L.; Mende, H.; Hardt, C.; Vorgerd, M.; Bürk, K.; Matson, W.; Dichgans, J.; Beal, M.F.; et al. Oxidative Stress in Patients with Friedreich Ataxia. Neurology 2000, 55, 1719–1721. [Google Scholar] [CrossRef] [PubMed]
- Rötig, A.; de Lonlay, P.; Chretien, D.; Foury, F.; Koenig, M.; Sidi, D.; Munnich, A.; Rustin, P. Aconitase and Mitochondrial Iron-Sulphur Protein Deficiency in Friedreich Ataxia. Nat. Genet. 1997, 17, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Huynen, M.A.; Snel, B.; Bork, P.; Gibson, T.J. The Phylogenetic Distribution of Frataxin Indicates a Role in Iron-Sulfur Cluster Protein Assembly. Hum. Mol. Genet. 2001, 10, 2463–2468. [Google Scholar] [CrossRef]
- Yoon, T.; Cowan, J.A. Iron-Sulfur Cluster Biosynthesis. Characterization of Frataxin as an Iron Donor for Assembly of [2Fe-2S] Clusters in ISU-Type Proteins. J. Am. Chem. Soc. 2003, 125, 6078–6084. [Google Scholar] [CrossRef] [PubMed]
- Gerber, J.; Mühlenhoff, U.; Lill, R. An Interaction between Frataxin and Isu1/Nfs1 That Is Crucial for Fe/S Cluster Synthesis on Isu1. EMBO Rep. 2003, 4, 906–911. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.; Wattenhofer-Donzé, M.; Schmucker, S.; Bouvet, S.; Reutenauer, L.; Puccio, H. Frataxin Is Essential for Extramitochondrial Fe-S Cluster Proteins in Mammalian Tissues. Hum. Mol. Genet. 2007, 16, 2651–2658. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Napoli, E.; Cortopassi, G. Mitochondrial Frataxin Interacts with ISD11 of the NFS1/ISCU Complex and Multiple Mitochondrial Chaperones. Hum. Mol. Genet. 2007, 16, 929–941. [Google Scholar] [CrossRef]
- Schmucker, S.; Martelli, A.; Colin, F.; Page, A.; Wattenhofer-Donzé, M.; Reutenauer, L.; Puccio, H. Mammalian Frataxin: An Essential Function for Cellular Viability through an Interaction with a Preformed ISCU/NFS1/ISD11 Iron-Sulfur Assembly Complex. PLoS ONE 2011, 6, e16199. [Google Scholar] [CrossRef]
- Boniecki, M.T.; Freibert, S.A.; Mühlenhoff, U.; Lill, R.; Cygler, M. Structure and Functional Dynamics of the Mitochondrial Fe/S Cluster Synthesis Complex. Nat. Commun. 2017, 8, 1287. [Google Scholar] [CrossRef]
- Lill, R.; Freibert, S.-A. Mechanisms of Mitochondrial Iron-Sulfur Protein Biogenesis. Annu. Rev. Biochem. 2020, 89, 471–499. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-L.; Barondeau, D.P. Human Frataxin Is an Allosteric Switch That Activates the Fe-S Cluster Biosynthetic Complex. Biochemistry 2010, 49, 9132–9139. [Google Scholar] [CrossRef] [PubMed]
- Colin, F.; Martelli, A.; Clémancey, M.; Latour, J.-M.; Gambarelli, S.; Zeppieri, L.; Birck, C.; Page, A.; Puccio, H.; Ollagnier de Choudens, S. Mammalian Frataxin Controls Sulfur Production and Iron Entry during de Novo Fe4S4 Cluster Assembly. J. Am. Chem. Soc. 2013, 135, 733–740. [Google Scholar] [CrossRef]
- Bridwell-Rabb, J.; Fox, N.G.; Tsai, C.-L.; Winn, A.M.; Barondeau, D.P. Human Frataxin Activates Fe-S Cluster Biosynthesis by Facilitating Sulfur Transfer Chemistry. Biochemistry 2014, 53, 4904–4913. [Google Scholar] [CrossRef] [PubMed]
- Parent, A.; Elduque, X.; Cornu, D.; Belot, L.; Le Caer, J.-P.; Grandas, A.; Toledano, M.B.; D’Autréaux, B. Mammalian Frataxin Directly Enhances Sulfur Transfer of NFS1 Persulfide to Both ISCU and Free Thiols. Nat. Commun. 2015, 6, 5686. [Google Scholar] [CrossRef] [PubMed]
- Fox, N.G.; Das, D.; Chakrabarti, M.; Lindahl, P.A.; Barondeau, D.P. Frataxin Accelerates [2Fe-2S] Cluster Formation on the Human Fe-S Assembly Complex. Biochemistry 2015, 54, 3880–3889. [Google Scholar] [CrossRef] [PubMed]
- Gervason, S.; Larkem, D.; Mansour, A.B.; Botzanowski, T.; Müller, C.S.; Pecqueur, L.; Le Pavec, G.; Delaunay-Moisan, A.; Brun, O.; Agramunt, J.; et al. Physiologically Relevant Reconstitution of Iron-Sulfur Cluster Biosynthesis Uncovers Persulfide-Processing Functions of Ferredoxin-2 and Frataxin. Nat. Commun. 2019, 10, 3566. [Google Scholar] [CrossRef] [PubMed]
- Musco, G.; Stier, G.; Kolmerer, B.; Adinolfi, S.; Martin, S.; Frenkiel, T.; Gibson, T.; Pastore, A. Towards a Structural Understanding of Friedreich’s Ataxia: The Solution Structure of Frataxin. Structure 2000, 8, 695–707. [Google Scholar] [CrossRef] [PubMed]
- Dhe-Paganon, S.; Shigeta, R.; Chi, Y.I.; Ristow, M.; Shoelson, S.E. Crystal Structure of Human Frataxin. J. Biol. Chem. 2000, 275, 30753–30756. [Google Scholar] [CrossRef]
- Adinolfi, S.; Trifuoggi, M.; Politou, A.S.; Martin, S.; Pastore, A. A Structural Approach to Understanding the Iron-Binding Properties of Phylogenetically Different Frataxins. Hum. Mol. Genet. 2002, 11, 1865–1877. [Google Scholar] [CrossRef]
- Cavadini, P.; O’Neill, H.A.; Benada, O.; Isaya, G. Assembly and Iron-Binding Properties of Human Frataxin, the Protein Deficient in Friedreich Ataxia. Hum. Mol. Genet. 2002, 11, 217–227. [Google Scholar] [CrossRef]
- Nair, M.; Adinolfi, S.; Pastore, C.; Kelly, G.; Temussi, P.; Pastore, A. Solution Structure of the Bacterial Frataxin Ortholog, CyaY: Mapping the Iron Binding Sites. Structure 2004, 12, 2037–2048. [Google Scholar] [CrossRef]
- He, Y.; Alam, S.L.; Proteasa, S.V.; Zhang, Y.; Lesuisse, E.; Dancis, A.; Stemmler, T.L. Yeast Frataxin Solution Structure, Iron Binding, and Ferrochelatase Interaction. Biochemistry 2004, 43, 16254–16262. [Google Scholar] [CrossRef]
- Bou-Abdallah, F.; Adinolfi, S.; Pastore, A.; Laue, T.M.; Dennis Chasteen, N. Iron Binding and Oxidation Kinetics in Frataxin CyaY of Escherichia Coli. J. Mol. Biol. 2004, 341, 605–615. [Google Scholar] [CrossRef]
- Cook, J.D.; Bencze, K.Z.; Jankovic, A.D.; Crater, A.K.; Busch, C.N.; Bradley, P.B.; Stemmler, A.J.; Spaller, M.R.; Stemmler, T.L. Monomeric Yeast Frataxin Is an Iron-Binding Protein. Biochemistry 2006, 45, 7767–7777. [Google Scholar] [CrossRef]
- Yoon, T.; Dizin, E.; Cowan, J.A. N-Terminal Iron-Mediated Self-Cleavage of Human Frataxin: Regulation of Iron Binding and Complex Formation with Target Proteins. J. Biol. Inorg. Chem. 2007, 12, 535–542. [Google Scholar] [CrossRef]
- Gentry, L.E.; Thacker, M.A.; Doughty, R.; Timkovich, R.; Busenlehner, L.S. His86 from the N-Terminus of Frataxin Coordinates Iron and Is Required for Fe-S Cluster Synthesis. Biochemistry 2013, 52, 6085–6096. [Google Scholar] [CrossRef] [PubMed]
- Ahlgren, E.-C.; Fekry, M.; Wiemann, M.; Söderberg, C.A.; Bernfur, K.; Gakh, O.; Rasmussen, M.; Højrup, P.; Emanuelsson, C.; Isaya, G.; et al. Iron-Induced Oligomerization of Human FXN81-210 and Bacterial CyaY Frataxin and the Effect of Iron Chelators. PLoS One 2017, 12, e0188937. [Google Scholar] [CrossRef] [PubMed]
- Bellanda, M.; Maso, L.; Doni, D.; Bortolus, M.; De Rosa, E.; Lunardi, F.; Alfonsi, A.; Noguera, M.E.; Herrera, M.G.; Santos, J.; et al. Exploring Iron-Binding to Human Frataxin and to Selected Friedreich Ataxia Mutants by Means of NMR and EPR Spectroscopies. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 140254. [Google Scholar] [CrossRef]
- Doni, D.; Passerini, L.; Audran, G.; Marque, S.R.A.; Schulz, M.; Santos, J.; Costantini, P.; Bortolus, M.; Carbonera, D. Effects of Fe2+/Fe3+ Binding to Human Frataxin and Its D122Y Variant, as Revealed by Site-Directed Spin Labeling (SDSL) EPR Complemented by Fluorescence and Circular Dichroism Spectroscopies. Int. J. Mol. Sci. 2020, 21, 9619. [Google Scholar] [CrossRef] [PubMed]
- Doni, D.; Meggiolaro, M.; Santos, J.; Audran, G.; Marque, S.R.A.; Costantini, P.; Bortolus, M.; Carbonera, D. A Combined Spectroscopic and In Silico Approach to Evaluate the Interaction of Human Frataxin with Mitochondrial Superoxide Dismutase. Biomedicines 2021, 9, 1763. [Google Scholar] [CrossRef]
- Doni, D.; Rigoni, G.; Palumbo, E.; Baschiera, E.; Peruzzo, R.; De Rosa, E.; Caicci, F.; Passerini, L.; Bettio, D.; Russo, A.; et al. The Displacement of Frataxin from the Mitochondrial Cristae Correlates with Abnormal Respiratory Supercomplexes Formation and Bioenergetic Defects in Cells of Friedreich Ataxia Patients. FASEB J. 2021, 35, e21362. [Google Scholar] [CrossRef] [PubMed]
- Doni, D.; Cavion, F.; Bortolus, M.; Baschiera, E.; Muccioli, S.; Tombesi, G.; d’Ettorre, F.; Ottaviani, D.; Marchesan, E.; Leanza, L.; et al. Human Frataxin, the Friedreich Ataxia Deficient Protein, Interacts with Mitochondrial Respiratory Chain. Cell Death Dis. 2023, 14, 805. [Google Scholar] [CrossRef]
- Sazanov, L.A.; Hinchliffe, P. Structure of the Hydrophilic Domain of Respiratory Complex I from Thermus Thermophilus. Science 2006, 311, 1430–1436. [Google Scholar] [CrossRef]
- Hinchliffe, P.; Carroll, J.; Sazanov, L.A. Identification of a Novel Subunit of Respiratory Complex I from Thermus Thermophilus. Biochemistry 2006, 45, 4413–4420. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, A.G.; Landmann, H.; Nette, D.; Burghaus, O.; Peuckert, F.; Seubert, A.; Miethke, M.; Marahiel, M.A. The Frataxin Homologue Fra Plays a Key Role in Intracellular Iron Channeling in Bacillus Subtilis. ChemBioChem 2011, 12, 2052–2061. [Google Scholar] [CrossRef] [PubMed]
- Monzon, A.M.; Rohr, C.O.; Fornasari, M.S.; Parisi, G. CoDNaS 2.0: A Comprehensive Database of Protein Conformational Diversity in the Native State. Database 2016, 2016, baw038. [Google Scholar] [CrossRef] [PubMed]
- Kmiecik, S.; Gront, D.; Kolinski, M.; Wieteska, L.; Dawid, A.E.; Kolinski, A. Coarse-Grained Protein Models and Their Applications. Chem. Rev. 2016, 116, 7898–7936. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, Q.; Qu, G.; Feng, Y.; Reetz, M.T. Utility of B-Factors in Protein Science: Interpreting Rigidity, Flexibility, and Internal Motion and Engineering Thermostability. Chem. Rev. 2019, 119, 1626–1665. [Google Scholar] [CrossRef]
- Schanda, P.; Brutscher, B. Very Fast Two-Dimensional NMR Spectroscopy for Real-Time Investigation of Dynamic Events in Proteins on the Time Scale of Seconds. J. Am. Chem. Soc. 2005, 127, 8014–8015. [Google Scholar] [CrossRef]
- Micsonai, A.; Moussong, É.; Wien, F.; Boros, E.; Vadászi, H.; Murvai, N.; Lee, Y.-H.; Molnár, T.; Réfrégiers, M.; Goto, Y.; et al. BeStSel: Webserver for Secondary Structure and Fold Prediction for Protein CD Spectroscopy. Nucleic Acids Res. 2022, 50, W90–W98. [Google Scholar] [CrossRef]
- Drew, E.D.; Janes, R.W. PDBMD2CD: Providing Predicted Protein Circular Dichroism Spectra from Multiple Molecular Dynamics-Generated Protein Structures. Nucleic Acids Res. 2020, 48, W17–W24. [Google Scholar] [CrossRef]
- Faraj, S.E.; Roman, E.A.; Aran, M.; Gallo, M.; Santos, J. The Alteration of the C-Terminal Region of Human Frataxin Distorts Its Structural Dynamics and Function. FEBS J. 2014, 281, 3397–3419. [Google Scholar] [CrossRef]
- Patra, S.; Barondeau, D.P. Mechanism of Activation of the Human Cysteine Desulfurase Complex by Frataxin. Proc. Natl. Acad. Sci. USA 2019, 116, 19421–19430. [Google Scholar] [CrossRef]
- Marelja, Z.; Stöcklein, W.; Nimtz, M.; Leimkühler, S. A Novel Role for Human Nfs1 in the Cytoplasm: Nfs1 Acts as a Sulfur Donor for MOCS3, a Protein Involved in Molybdenum Cofactor Biosynthesis. J. Biol. Chem. 2008, 283, 25178–25185. [Google Scholar] [CrossRef]
- Siegel, L.M. A DIRECT MICRODETERMINATION FOR SULFIDE. Anal. Biochem. 1965, 11, 126–132. [Google Scholar] [CrossRef]
- Cai, K.; Frederick, R.O.; Tonelli, M.; Markley, J.L. ISCU(M108I) and ISCU(D39V) Differ from Wild-Type ISCU in Their Failure To Form Cysteine Desulfurase Complexes Containing Both Frataxin and Ferredoxin. Biochemistry 2018, 57, 1491–1500. [Google Scholar] [CrossRef]
- Chothia, C.; Lesk, A.M. The Relation between the Divergence of Sequence and Structure in Proteins. EMBO J. 1986, 5, 823–826. [Google Scholar] [CrossRef]
- Koehl, P. Protein Structure Similarities. Curr. Opin. Struct. Biol. 2001, 11, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Szczepanowska, K.; Senft, K.; Heidler, J.; Herholz, M.; Kukat, A.; Höhne, M.N.; Hofsetz, E.; Becker, C.; Kaspar, S.; Giese, H.; et al. A Salvage Pathway Maintains Highly Functional Respiratory Complex I. Nat. Commun. 2020, 11, 1643. [Google Scholar] [CrossRef] [PubMed]
- Berrisford, J.M.; Sazanov, L.A. Structural Basis for the Mechanism of Respiratory Complex I. J. Biol. Chem. 2009, 284, 29773–29783. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Cowan, J.A. A Structural and Functional Homolog Supports a General Role for Frataxin in Cellular Iron Chemistry. Chem. Commun. 2010, 46, 719–721. [Google Scholar] [CrossRef] [PubMed]
- Fox, N.G.; Yu, X.; Feng, X.; Bailey, H.J.; Martelli, A.; Nabhan, J.F.; Strain-Damerell, C.; Bulawa, C.; Yue, W.W.; Han, S. Structure of the Human Frataxin-Bound Iron-Sulfur Cluster Assembly Complex Provides Insight into Its Activation Mechanism. Nat. Commun. 2019, 10, 2210. [Google Scholar] [CrossRef]
- Tsai, C.-L.; Bridwell-Rabb, J.; Barondeau, D.P. Friedreich’s Ataxia Variants I154F and W155R Diminish Frataxin-Based Activation of the Iron-Sulfur Cluster Assembly Complex. Biochemistry 2011, 50, 6478–6487. [Google Scholar] [CrossRef]
- Campbell, C.J.; Pall, A.E.; Naik, A.R.; Thompson, L.N.; Stemmler, T.L. Molecular Details of the Frataxin-Scaffold Interaction during Mitochondrial Fe-S Cluster Assembly. Int. J. Mol. Sci. 2021, 22, 6006. [Google Scholar] [CrossRef]
- Sewell, K.E.; Gola, G.F.; Pignataro, M.F.; Herrera, M.G.; Noguera, M.E.; Olmos, J.; Ramírez, J.A.; Capece, L.; Aran, M.; Santos, J. Direct Cysteine Desulfurase Activity Determination by NMR and the Study of the Functional Role of Key Structural Elements of Human NFS1. ACS Chem. Biol. 2023, 18, 1534–1547. [Google Scholar] [CrossRef]
- Marczenko, Z. Spectrophotometric Determination of Elements; Chalmer, R.A., Ed.; Wiley: New York, NY, USA, 1976. [Google Scholar]
- Gabler, F.; Nam, S.-Z.; Till, S.; Mirdita, M.; Steinegger, M.; Söding, J.; Lupas, A.N.; Alva, V. Protein Sequence Analysis Using the MPI Bioinformatics Toolkit. Curr. Protoc. Bioinform. 2020, 72, e108. [Google Scholar] [CrossRef]
- van Kempen, M.; Kim, S.S.; Tumescheit, C.; Mirdita, M.; Lee, J.; Gilchrist, C.L.M.; Söding, J.; Steinegger, M. Fast and Accurate Protein Structure Search with Foldseek. Nat. Biotechnol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Holm, L.; Laiho, A.; Törönen, P.; Salgado, M. DALI Shines a Light on Remote Homologs: One Hundred Discoveries. Protein Sci. 2023, 32, e4519. [Google Scholar] [CrossRef]
- Andreeva, A.; Kulesha, E.; Gough, J.; Murzin, A.G. The SCOP Database in 2020: Expanded Classification of Representative Family and Superfamily Domains of Known Protein Structures. Nucleic Acids Res. 2020, 48, D376–D382. [Google Scholar] [CrossRef] [PubMed]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- Grant, B.J.; Rodrigues, A.P.C.; ElSawy, K.M.; McCammon, J.A.; Caves, L.S.D. Bio3d: An R Package for the Comparative Analysis of Protein Structures. Bioinformatics 2006, 22, 2695–2696. [Google Scholar] [CrossRef] [PubMed]
- Kuriata, A.; Gierut, A.M.; Oleniecki, T.; Ciemny, M.P.; Kolinski, A.; Kurcinski, M.; Kmiecik, S. CABS-Flex 2.0: A Web Server for Fast Simulations of Flexibility of Protein Structures. Nucleic Acids Res. 2018, 46, W338–W343. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Organism | Representative PDB ID |
|---|---|---|
| Frataxin-like | Saccharomyces cerevisiae S288C | 5TRE_C |
| Frataxin-like | Saccharomyces cerevisiae S288C | 2FQL_A |
| Frataxin-like | Saccharomyces cerevisiae S288C | 4EC2_A |
| Frataxin-like | Saccharomyces cerevisiae S288C | 3OER_A |
| Frataxin-like | Homo sapiens | 3S5F_A |
| Frataxin-like | Homo sapiens | 3T3T_C |
| Frataxin-like | Homo sapiens | 1EKG_A |
| Frataxin-like | Homo sapiens | 6NZU_I |
| Frataxin-like | Homo sapiens | 3S4M_A |
| Frataxin-like | Psychromonas ingrahamii 37 | 4HS5_A |
| Frataxin-like | Psychromonas ingrahamii 37 | 4LK8_A |
| Frataxin-like | Psychromonas ingrahamii 37 | 4LP1_A |
| Frataxin-like | Chaetomium thermophilum var. thermophilum DSM 1495 | 6FCO_C |
| Frataxin-like | Escherichia coli K-12 | 1EW4_A |
| Frataxin-like | Escherichia coli K-12 | 2EFF_A |
| Frataxin-like | Escherichia coli K-12 | 2P1X_A |
| Frataxin-like | Escherichia coli K-12 | 1SOY_A |
| Frataxin-like | Burkholderia cenocepacia J2315 | 4JPD_A |
| Nqo15-like | Thermus thermophilus HB8 | 2ZJL_7 |
| Nqo15-like | Thermus thermophilus HB8 | 6I0D_I |
| Nqo15-like | Thermus thermophilus HB8 | 6Q8X_7 |
| Nqo15-like | Thermus thermophilus HB8 | 6I1P_7 |
| Nqo15-like | Thermus thermophilus HB8 | 6I1P_I |
| Nqo15-like | Thermus thermophilus HB8 | 6Y11_I |
| Nqo15-like | Thermus thermophilus HB8 | 3IAS_Q |
| Nqo15-like | Thermus thermophilus HB8 | 2FUG_Q |
| Nqo15-like | Thermus thermophilus HB8 | 3M9S_7 |
| Nqo15-like | Thermus thermophilus HB8 | 3IAM_7 |
| Nqo15-like | Thermus thermophilus HB8 | 4HEA_7 |
| Nqo15-like | Thermus thermophilus HB8 | 6Y11_7 |
| YdhG-like | Lactobacillus paracasei ATCC 334 | 2I8D_A |
| YdhG-like | Bacillus subtilis subsp. subtilis str. 168 | 2OC6_A |
| YdhG-like | Bacillus halodurans C-125 | 2KL4_A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doni, D.; Cavallari, E.; Noguera, M.E.; Gentili, H.G.; Cavion, F.; Parisi, G.; Fornasari, M.S.; Sartori, G.; Santos, J.; Bellanda, M.; et al. Searching for Frataxin Function: Exploring the Analogy with Nqo15, the Frataxin-like Protein of Respiratory Complex I from Thermus thermophilus. Int. J. Mol. Sci. 2024, 25, 1912. https://doi.org/10.3390/ijms25031912
Doni D, Cavallari E, Noguera ME, Gentili HG, Cavion F, Parisi G, Fornasari MS, Sartori G, Santos J, Bellanda M, et al. Searching for Frataxin Function: Exploring the Analogy with Nqo15, the Frataxin-like Protein of Respiratory Complex I from Thermus thermophilus. International Journal of Molecular Sciences. 2024; 25(3):1912. https://doi.org/10.3390/ijms25031912
Chicago/Turabian StyleDoni, Davide, Eva Cavallari, Martin Ezequiel Noguera, Hernan Gustavo Gentili, Federica Cavion, Gustavo Parisi, Maria Silvina Fornasari, Geppo Sartori, Javier Santos, Massimo Bellanda, and et al. 2024. "Searching for Frataxin Function: Exploring the Analogy with Nqo15, the Frataxin-like Protein of Respiratory Complex I from Thermus thermophilus" International Journal of Molecular Sciences 25, no. 3: 1912. https://doi.org/10.3390/ijms25031912
APA StyleDoni, D., Cavallari, E., Noguera, M. E., Gentili, H. G., Cavion, F., Parisi, G., Fornasari, M. S., Sartori, G., Santos, J., Bellanda, M., Carbonera, D., Costantini, P., & Bortolus, M. (2024). Searching for Frataxin Function: Exploring the Analogy with Nqo15, the Frataxin-like Protein of Respiratory Complex I from Thermus thermophilus. International Journal of Molecular Sciences, 25(3), 1912. https://doi.org/10.3390/ijms25031912