
The Impact of 90 Parkinson’s Disease-Risk Single Nucleotide Polymorphisms on Urinary Bis(monoacylglycerol)phosphate Levels in the Prodromal and PD Cohorts

Abstract
:1. Introduction
2. Results
2.1. The Prodromal Cohort Had Higher Urinary BMP Levels than the PD and Healthy Controls
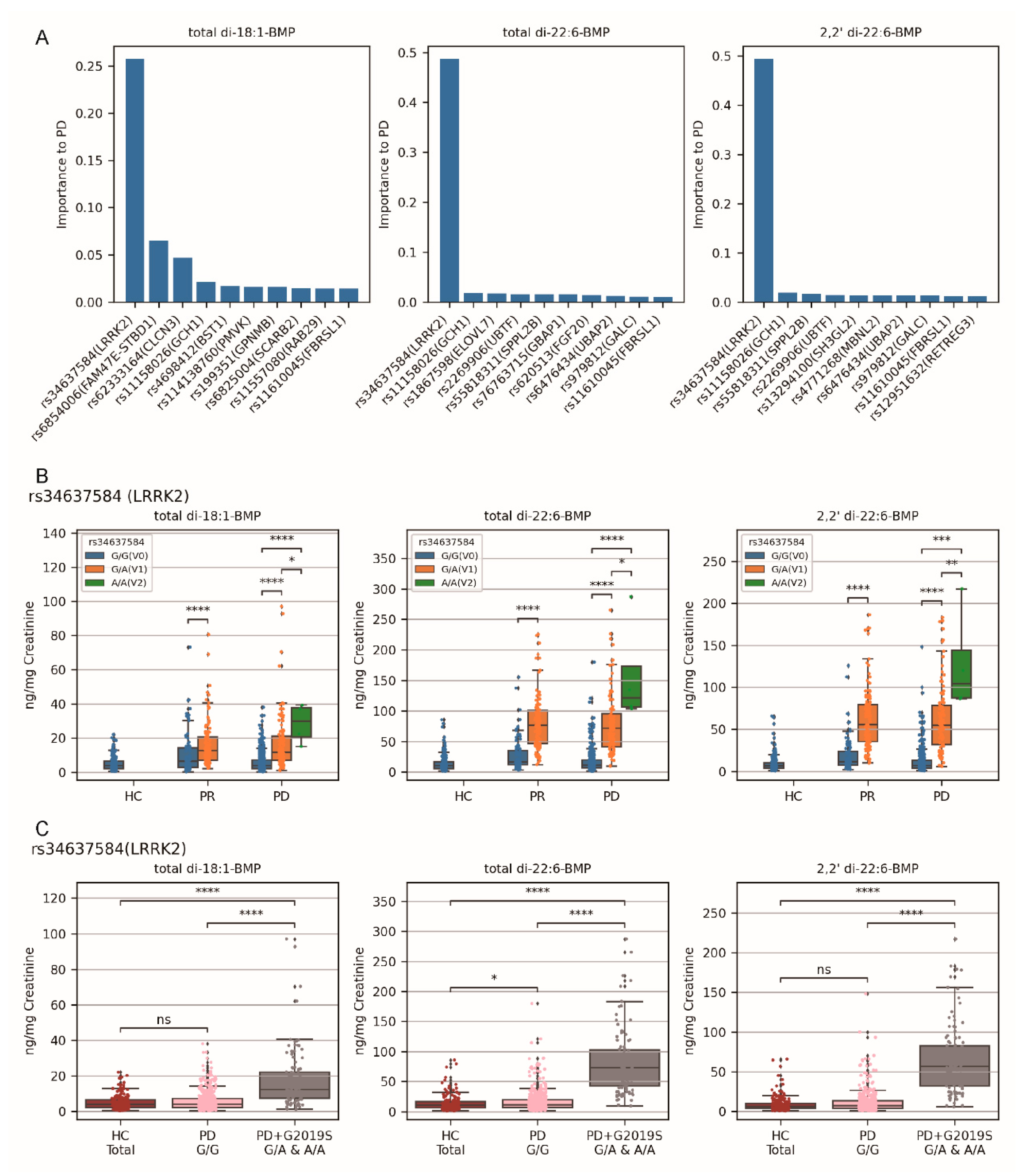
2.2. The Impact of Variants of 90 PD-Risk SNPs Varied in Different Cohorts of PD
2.3. Variants rs34637584 G/A and rs34637584 A/A (LRRK2 G2019S) Were the Primary Genetic Factors for Elevated Urinary BMP Levels in the PD Cohort than in Healthy Controls
2.4. Variant rs76763715 T/T (GBA1) Emerged as the Primary Factors to Elevated Urinary BMP Levels in the PR Cohort Compared to PD Cohort
2.5. The Overall Significance of Factors on Urinary BMP Levels
2.6. Pathway and Process-Enrichment Analysis for Alterations in BMP Level
3. Discussions
4. Materials and Methods
4.1. Data Source
4.2. Hypothesis Tests for Significance Check
4.3. Heatmap for p-Values of Impact of SNPs’ Variants on BMP Levels
4.4. Assessment of Factors Influencing Urinary BMP Levels
4.5. Proteomics Data Analysis
4.6. Data Calculation and Visualization
5. Conclusions
6. Innovations and Limitations of Our Work
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hayes, M.T. Parkinson’s Disease and Parkinsonism. Am. J. Med. 2019, 132, 802–807. [Google Scholar] [CrossRef]
- Lill, C.M. Genetics of Parkinson’s disease. Mol. Cell Probes 2016, 30, 386–396. [Google Scholar] [CrossRef]
- Balestrino, R.; Schapira, A.H.V. Parkinson disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Dorsey, E.R.; Constantinescu, R.; Thompson, J.P.; Biglan, K.M.; Holloway, R.G.; Kieburtz, K.; Marshall, F.J.; Ravina, B.M.; Schifitto, G.; Siderowf, A.; et al. Projected number of people with Parkinson disease in the most populous nations, 2005 through 2030. Neurology 2007, 68, 384–386. [Google Scholar] [CrossRef]
- Tysnes, O.B.; Storstein, A. Epidemiology of Parkinson’s disease. J. Neural Transm. 2017, 124, 901–905. [Google Scholar] [CrossRef] [PubMed]
- Kieburtz, K.; Wunderle, K.B. Parkinson’s disease: Evidence for environmental risk factors. Mov. Disord. 2013, 28, 8–13. [Google Scholar] [CrossRef]
- Noyce, A.J.; Bestwick, J.P.; Silveira-Moriyama, L.; Hawkes, C.H.; Giovannoni, G.; Lees, A.J.; Schrag, A. Meta-analysis of early nonmotor features and risk factors for Parkinson disease. Ann. Neurol. 2012, 72, 893–901. [Google Scholar] [CrossRef]
- Blauwendraat, C.; Nalls, M.A.; Singleton, A.B. The genetic architecture of Parkinson’s disease. Lancet Neurol. 2020, 19, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Cerri, S.; Mus, L.; Blandini, F. Parkinson’s Disease in Women and Men: What’s the Difference? J. Park. Dis. 2019, 9, 501–515. [Google Scholar] [CrossRef]
- Warren, N.; O’Gorman, C.; Lehn, A.; Siskind, D. Dopamine dysregulation syndrome in Parkinson’s disease: A systematic review of published cases. J. Neurol. Neurosurg. Psychiatry 2017, 88, 1060–1064. [Google Scholar] [CrossRef]
- Tolosa, E.; Garrido, A.; Scholz, S.W.; Poewe, W. Challenges in the diagnosis of Parkinson’s disease. Lancet Neurol. 2021, 20, 385–397. [Google Scholar] [CrossRef]
- Berg, D.; Borghammer, P.; Fereshtehnejad, S.M.; Heinzel, S.; Horsager, J.; Schaeffer, E.; Postuma, R.B. Prodromal Parkinson disease subtypes–key to understanding heterogeneity. Nat. Rev. Neurol. 2021, 17, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Mahlknecht, P.; Seppi, K.; Poewe, W. The Concept of Prodromal Parkinson’s Disease. J. Park. Dis. 2015, 5, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Berg, D.; Postuma, R.B.; Adler, C.H.; Bloem, B.R.; Chan, P.; Dubois, B.; Gasser, T.; Goetz, C.G.; Halliday, G.; Joseph, L.; et al. MDS research criteria for prodromal Parkinson’s disease. Mov. Disord. 2015, 30, 1600–1611. [Google Scholar] [CrossRef] [PubMed]
- Heinzel, S.; Berg, D.; Gasser, T.; Chen, H.; Yao, C.; Postuma, R.B.; the MDS Task Force on the Definition of Parkinson’s Disease. Update of the MDS research criteria for prodromal Parkinson’s disease. Mov. Disord. 2019, 34, 1464–1470. [Google Scholar] [CrossRef] [PubMed]
- Fayyad, M.; Salim, S.; Majbour, N.; Erskine, D.; Stoops, E.; Mollenhauer, B.; El-Agnaf, O.M.A. Parkinson’s disease biomarkers based on alpha-synuclein. J. Neurochem. 2019, 150, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Spillantini, M.G.; Goedert, M. Neurodegeneration and the ordered assembly of alpha-synuclein. Cell Tissue Res. 2018, 373, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Moors, T.; Paciotti, S.; Chiasserini, D.; Calabresi, P.; Parnetti, L.; Beccari, T.; van de Berg, W.D. Lysosomal Dysfunction and alpha-Synuclein Aggregation in Parkinson’s Disease: Diagnostic Links. Mov. Disord. 2016, 31, 791–801. [Google Scholar] [CrossRef]
- Parnetti, L.; Paciotti, S.; Eusebi, P.; Dardis, A.; Zampieri, S.; Chiasserini, D.; Tasegian, A.; Tambasco, N.; Bembi, B.; Calabresi, P.; et al. Cerebrospinal fluid beta-glucocerebrosidase activity is reduced in parkinson’s disease patients. Mov. Disord. 2017, 32, 1423–1431. [Google Scholar] [CrossRef]
- Lotankar, S.; Prabhavalkar, K.S.; Bhatt, L.K. Biomarkers for Parkinson’s Disease: Recent Advancement. Neurosci. Bull. 2017, 33, 585–597. [Google Scholar] [CrossRef]
- Parnetti, L.; Gaetani, L.; Eusebi, P.; Paciotti, S.; Hansson, O.; El-Agnaf, O.; Mollenhauer, B.; Blennow, K.; Calabresi, P. CSF and blood biomarkers for Parkinson’s disease. Lancet Neurol. 2019, 18, 573–586. [Google Scholar] [CrossRef]
- Ganguly, U.; Singh, S.; Pal, S.; Prasad, S.; Agrawal, B.K.; Saini, R.V.; Chakrabarti, S. Alpha-Synuclein as a Biomarker of Parkinson’s Disease: Good, but Not Good Enough. Front. Aging Neurosci. 2021, 13, 702639. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.T.; Bullock, K.M.; Erickson, M.A.; Zhang, J.; Banks, W.A. Alpha synuclein is transported into and out of the brain by the blood-brain barrier. Peptides 2014, 62, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Le, W. Biomarkers for Parkinson’s Disease: How Good Are They? Neurosci. Bull. 2020, 36, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Bloem, B.R.; Okun, M.S.; Klein, C. Parkinson’s disease. Lancet 2021, 397, 2284–2303. [Google Scholar] [CrossRef] [PubMed]
- Malpartida, A.B.; Williamson, M.; Narendra, D.P.; Wade-Martins, R.; Ryan, B.J. Mitochondrial Dysfunction and Mitophagy in Parkinson’s Disease: From Mechanism to Therapy. Trends Biochem. Sci. 2021, 46, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Muqit, M.M.K. Parkinson’s: A Disease of Aberrant Vesicle Trafficking. Annu. Rev. Cell Dev. Biol. 2020, 36, 237–264. [Google Scholar] [CrossRef]
- Nalls, M.A.; Blauwendraat, C.; Vallerga, C.L.; Heilbron, K.; Bandres-Ciga, S.; Chang, D.; Tan, M.; Kia, D.A.; Noyce, A.J.; Xue, A.; et al. Identification of novel risk loci, causal insights, and heritable risk for Parkinson’s disease: A meta-analysis of genome-wide association studies. Lancet Neurol. 2019, 18, 1091–1102. [Google Scholar] [CrossRef]
- Deng, H.; Wang, P.; Jankovic, J. The genetics of Parkinson disease. Ageing Res. Rev. 2018, 42, 72–85. [Google Scholar] [CrossRef]
- Scherer, M.; Schmitz, G. Metabolism, function and mass spectrometric analysis of bis(monoacylglycero)phosphate and cardiolipin. Chem. Phys. Lipids 2011, 164, 556–562. [Google Scholar] [CrossRef]
- Showalter, M.R.; Berg, A.L.; Nagourney, A.; Heil, H.; Carraway, K.L., III; Fiehn, O. The Emerging and Diverse Roles of Bis(monoacylglycero) Phosphate Lipids in Cellular Physiology and Disease. Int. J. Mol. Sci. 2020, 21, 8067. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.L.; Zhang, J.; Stewart, S.; Rosenzweig, B.A.; Shea, K.; Mans, D.; Colatsky, T. Comparison of urinary and serum levels of di-22:6-bis(monoacylglycerol)phosphate as noninvasive biomarkers of phospholipidosis in rats. Toxicol. Lett. 2012, 213, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Tengstrand, E.A.; Chourb, L.; Hsieh, F.Y. Di-22:6-bis(monoacylglycerol)phosphate: A clinical biomarker of drug-induced phospholipidosis for drug development and safety assessment. Toxicol. Appl. Pharmacol. 2014, 279, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Baronas, E.T.; Lee, J.W.; Alden, C.; Hsieh, F.Y. Biomarkers to monitor drug-induced phospholipidosis. Toxicol. Appl. Pharmacol. 2007, 218, 72–78. [Google Scholar] [CrossRef]
- Fuji, R.N.; Flagella, M.; Baca, M.; Baptista, M.A.; Brodbeck, J.; Chan, B.K.; Fiske, B.K.; Honigberg, L.; Jubb, A.M.; Katavolos, P.; et al. Effect of selective LRRK2 kinase inhibition on nonhuman primate lung. Sci. Transl. Med. 2015, 7, 273ra215. [Google Scholar] [CrossRef] [PubMed]
- Alcalay, R.N.; Hsieh, F.; Tengstrand, E.; Padmanabhan, S.; Baptista, M.; Kehoe, C.; Narayan, S.; Boehme, A.K.; Merchant, K. Higher Urine bis(Monoacylglycerol)Phosphate Levels in LRRK2 G2019S Mutation Carriers: Implications for Therapeutic Development. Mov. Disord. 2020, 35, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Merchant, K.M.; Simuni, T.; Fedler, J.; Caspell-Garcia, C.; Brumm, M.; Nudelman, K.N.H.; Tengstrandt, E.; Hsieh, F.; Alcalay, R.N.; Coffey, C.; et al. LRRK2 and GBA1 variant carriers have higher urinary bis(monacylglycerol) phosphate concentrations in PPMI cohorts. NPJ Park. Dis. 2023, 9, 30. [Google Scholar] [CrossRef] [PubMed]
- Parkinson Progression Marker, I. The Parkinson Progression Marker Initiative (PPMI). Prog. Neurobiol. 2011, 95, 629–635. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Behl, T.; Kumar, S.; Althafar, Z.M.; Sehgal, A.; Singh, S.; Sharma, N.; Badavath, V.N.; Yadav, S.; Bhatia, S.; Al-Harrasi, A.; et al. Exploring the Role of Ubiquitin-Proteasome System in Parkinson’s Disease. Mol. Neurobiol. 2022, 59, 4257–4273. [Google Scholar] [CrossRef]
- Wright, R. Mitochondrial dysfunction and Parkinson’s disease. Nat. Neurosci. 2022, 25, 2. [Google Scholar] [CrossRef]
- Navarro-Romero, A.; Montpeyo, M.; Martinez-Vicente, M. The Emerging Role of the Lysosome in Parkinson’s Disease. Cells 2020, 9, 2399. [Google Scholar] [CrossRef]
- Nguyen, M.; Wong, Y.C.; Ysselstein, D.; Severino, A.; Krainc, D. Synaptic, Mitochondrial, and Lysosomal Dysfunction in Parkinson’s Disease. Trends Neurosci. 2019, 42, 140–149. [Google Scholar] [CrossRef]
- Lu, J.; Wu, M.; Yue, Z. Autophagy and Parkinson’s Disease. Adv. Exp. Med. Biol. 2020, 1207, 21–51. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.R.; Lee, B.D. Pathological Functions of LRRK2 in Parkinson’s Disease. Cells 2020, 9, 2565. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.L.; Moore, D.J. LRRK2 and the Endolysosomal System in Parkinson’s Disease. J. Park. Dis. 2020, 10, 1271–1291. [Google Scholar] [CrossRef] [PubMed]
- Mamais, A.; Kluss, J.H.; Bonet-Ponce, L.; Landeck, N.; Langston, R.G.; Smith, N.; Beilina, A.; Kaganovich, A.; Ghosh, M.C.; Pellegrini, L.; et al. Mutations in LRRK2 linked to Parkinson disease sequester Rab8a to damaged lysosomes and regulate transferrin-mediated iron uptake in microglia. PLoS Biol. 2021, 19, e3001480. [Google Scholar] [CrossRef] [PubMed]
- Boecker, C.A.; Goldsmith, J.; Dou, D.; Cajka, G.G.; Holzbaur, E.L.F. Increased LRRK2 kinase activity alters neuronal autophagy by disrupting the axonal transport of autophagosomes. Curr. Biol. 2021, 31, 2140–2154 e2146. [Google Scholar] [CrossRef] [PubMed]
- Gegg, M.E.; Menozzi, E.; Schapira, A.H.V. Glucocerebrosidase-associated Parkinson disease: Pathogenic mechanisms and potential drug treatments. Neurobiol. Dis. 2022, 166, 105663. [Google Scholar] [CrossRef] [PubMed]
- Boer, D.E.C.; van Smeden, J.; Bouwstra, J.A.; Aerts, J. Glucocerebrosidase: Functions in and Beyond the Lysosome. J. Clin. Med. 2020, 9, 736. [Google Scholar] [CrossRef]
- Hruska, K.S.; LaMarca, M.E.; Scott, C.R.; Sidransky, E. Gaucher disease: Mutation and polymorphism spectrum in the glucocerebrosidase gene (GBA). Hum. Mutat. 2008, 29, 567–583. [Google Scholar] [CrossRef]
- Riboldi, G.M.; Di Fonzo, A.B. GBA, Gaucher Disease, and Parkinson’s Disease: From Genetic to Clinic to New Therapeutic Approaches. Cells 2019, 8, 364. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Sanz, P.; Orgaz, L.; Fuentes, J.M.; Vicario, C.; Moratalla, R. Cholesterol and multilamellar bodies: Lysosomal dysfunction in GBA-Parkinson disease. Autophagy 2018, 14, 717–718. [Google Scholar] [CrossRef] [PubMed]
- Rabia, M.; Leuzy, V.; Soulage, C.; Durand, A.; Fourmaux, B.; Errazuriz-Cerda, E.; Koffel, R.; Draeger, A.; Colosetti, P.; Jalabert, A.; et al. Bis(monoacylglycero)phosphate, a new lipid signature of endosome-derived extracellular vesicles. Biochimie 2020, 178, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Schulze, H.; Sandhoff, K. Lysosomal lipid storage diseases. Cold Spring Harb. Perspect. Biol. 2011, 3, a004804. [Google Scholar] [CrossRef] [PubMed]
- Bryant, N.; Malpeli, N.; Ziaee, J.; Blauwendraat, C.; Liu, Z.; Consortium, A.P.; West, A.B. Identification of LRRK2 missense variants in the accelerating medicines partnership Parkinson’s disease cohort. Hum. Mol. Genet. 2021, 30, 454–466. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Jian, B.; Liu, Z. Transmembrane Protein 175, a Lysosomal Ion Channel Related to Parkinson’s Disease. Biomolecules 2023, 13, 802. [Google Scholar] [CrossRef] [PubMed]
- Krohn, L.; Ozturk, T.N.; Vanderperre, B.; Ouled Amar Bencheikh, B.; Ruskey, J.A.; Laurent, S.B.; Spiegelman, D.; Postuma, R.B.; Arnulf, I.; Hu, M.T.M.; et al. Genetic, Structural, and Functional Evidence Link TMEM175 to Synucleinopathies. Ann. Neurol. 2020, 87, 139–153. [Google Scholar] [CrossRef]
- Yu, H.; Sang, P.; Huan, T. Adaptive Box-Cox Transformation: A Highly Flexible Feature-Specific Data Transformation to Improve Metabolomic Data Normality for Better Statistical Analysis. Anal. Chem. 2022, 94, 8267–8276. [Google Scholar] [CrossRef]
- Kim, Y.J.; Cribbie, R.A. ANOVA and the variance homogeneity assumption: Exploring a better gatekeeper. Br. J. Math. Stat. Psychol. 2018, 71, 1–12. [Google Scholar] [CrossRef]
- Dexter, F. Wilcoxon-Mann-Whitney test used for data that are not normally distributed. Anesth. Analg. 2013, 117, 537–538. [Google Scholar] [CrossRef]
- Armstrong, R.A. When to use the Bonferroni correction. Ophthalmic Physiol. Opt. 2014, 34, 502–508. [Google Scholar] [CrossRef]
- Rigatti, S.J. Random Forest. J. Insur. Med. 2017, 47, 31–39. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort | HC n = 190 | PR n = 210 | PD n = 554 | p-Value ofHC vs. PR | p-Value ofHC vs. PD | p-Value ofPR vs. PD |
|---|---|---|---|---|---|---|
| Female number, % | 67, 35.26% | 123, 58.57% | 215, 38.81% | − | − | − |
| Age (years),mean ± SD | 60.83 ± 11.18 | 62.61 ± 7.30 | 62.40 ± 9.63 | 0.4590 a | 0.8893 a | 0.4590 a |
| p-value of age between sex | 0.0549 a | 0.4051 a | 0.9245 a | − | − | − |
| p-value of total di-18:1-BMP between sex | 0.8208 b | 0.8559 b | 0.1410 b | − | − | − |
| p-value of total di-22:6-BMP between sex | 0.4892 b | 0.4892 b | 0.0577 b | − | − | − |
| p-value of 2,2’ di-22:6-BMP between sex | 0.2588 b | 0.2398 b | 0.1739 b | − | − | − |
| total di-18:1-BMP, mean ± SD | 4.99 ± 4.23 | 13.53 ± 13.92 | 8.05 ± 9.96 | <0.0001 b | <0.0001 b | <0.0001 b |
| total di-22:6-BMP, mean ± SD | 13.83 ± 13.02 | 56.94 ± 45.22 | 29.83 ± 38.79 | <0.0001 b | <0.0001 b | <0.0001 b |
| 2,2’ di-22:6-BM, mean ± SD | 9.11 ± 9.70 | 43.56 ± 37.24 | 22.07 ± 31.07 | <0.0001 b | <0.0001 b | <0.0001 b |
| Covariates | Cohort | Total di-18:1-BMP | Total di-22:6-BMP | 2,2’ di-22:6-BMP | rs34637584 (LRRK2) | rs76763715 (GBA1) | ||
|---|---|---|---|---|---|---|---|---|
| Sex and age | Cohort | corr | 1.000 | 0.033 | 0.046 | 0.051 | 0.073 | −0.023 |
| p | . | 0.367 | 0.201 | 0.162 | 0.044 | 0.522 | ||
| df | 0 | 758 | 758 | 758 | 758 | 758 | ||
| Total di-18:1-BMP | corr | 0.033 | 1.000 | 0.502 | 0.509 | 0.412 | −0.108 | |
| p | 0.367 | . | 0.000 | 0.000 | 0.000 | 0.003 | ||
| df | 758 | 0 | 758 | 758 | 758 | 758 | ||
| Total di-22:6-BMP | corr | 0.046 | 0.502 | 1.000 | 0.986 | 0.680 | −0.143 | |
| p | 0.201 | 0.000 | . | 0.000 | 0.000 | 0.000 | ||
| df | 758 | 758 | 0 | 758 | 758 | 758 | ||
| 2,2’ di-22:6-BMP | corr | 0.051 | 0.509 | 0.986 | 1.000 | 0.681 | −0.148 | |
| p | 0.162 | 0.000 | 0.000 | . | 0.000 | 0.000 | ||
| df | 758 | 758 | 758 | 0 | 758 | 758 | ||
| rs34637584(LRRK2) | corr | 0.073 | 0.412 | 0.680 | 0.681 | 1.000 | −0.220 | |
| p | 0.044 | 0.000 | 0.000 | 0.000 | . | 0.000 | ||
| df | 758 | 758 | 758 | 758 | 0 | 758 | ||
| rs76763715(GBA1) | corr | −0.023 | −0.108 | −0.143 | −0.148 | −0.220 | 1.000 | |
| p | 0.522 | 0.003 | 0.000 | 0.000 | 0.000 | . | ||
| df | 758 | 758 | 758 | 758 | 758 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, S.; Lee, P.A.H.; Wang, Z.; Zhao, B. The Impact of 90 Parkinson’s Disease-Risk Single Nucleotide Polymorphisms on Urinary Bis(monoacylglycerol)phosphate Levels in the Prodromal and PD Cohorts. Int. J. Mol. Sci. 2024, 25, 2286. https://doi.org/10.3390/ijms25042286
Fang S, Lee PAH, Wang Z, Zhao B. The Impact of 90 Parkinson’s Disease-Risk Single Nucleotide Polymorphisms on Urinary Bis(monoacylglycerol)phosphate Levels in the Prodromal and PD Cohorts. International Journal of Molecular Sciences. 2024; 25(4):2286. https://doi.org/10.3390/ijms25042286
Chicago/Turabian StyleFang, Shuai, Priscilla Ann Hweek Lee, Zejian Wang, and Bo Zhao. 2024. "The Impact of 90 Parkinson’s Disease-Risk Single Nucleotide Polymorphisms on Urinary Bis(monoacylglycerol)phosphate Levels in the Prodromal and PD Cohorts" International Journal of Molecular Sciences 25, no. 4: 2286. https://doi.org/10.3390/ijms25042286