Abstract
As the principal ligand for NKG2D, MICA elicits the recruitment of subsets of T cells and NK cells in innate immunity. MICA gene variants greatly impact the functionality and expression of MICA in humans. The current study evaluated whether MICA polymorphisms distinctively influence the pathogenesis of psoriasis (PSO), rheumatoid arthritis (RA), and systemic lupus erythematosus (SLE) in Taiwanese subjects. The distributions of MICA alleles and levels of serum soluble NKG2D were compared between healthy controls and patients with PSO, RA, and SLE, respectively. The binding capacities and cell surface densities of MICA alleles were assessed by utilizing stable cell lines expressing four prominent Taiwanese MICA alleles. Our data revealed that MICA*010 was significantly associated with risks for PSO and RA (PFDR = 1.93 × 10−15 and 0.00112, respectively), while MICA*045 was significantly associated with predisposition to SLE (PFDR = 0.0002). On the other hand, MICA*002 was associated with protection against RA development (PFDR = 4.16 × 10−6), while MICA*009 was associated with a low risk for PSO (PFDR = 0.0058). MICA*002 exhibited the highest binding affinity for NKG2D compared to the other MICA alleles. Serum concentrations of soluble MICA were significantly elevated in SLE patients compared to healthy controls (p = 0.01). The lack of cell surface expression of the MICA*010 was caused by its entrapment in the endoplasmic reticulum. As a prevalent risk factor for PSO and RA, MICA*010 is deficient in cell surface expression and is unable to interact with NKG2D. Our study suggests that MICA alleles distinctively contribute to the pathogenesis of PSO, RA, and SLE in Taiwanese people.
1. Introduction
Natural killer (NK) cells possess an extensive repertoire of activating and inhibiting receptors, comprising natural killer group 2 member D (NKG2D), NKp30, NKp44, NKp46, killer-cell immunoglobulin-like receptors (KIR), and CD94/NKG2A receptors. NK cells employ inhibitory and activating receptors to monitor alterations in the expression of human leukocyte antigen (HLA) class I molecules and stress-induced ligands. This surveillance enables NK cells to achieve functional proficiency while gaining authorization for the elimination of target cells [1]. NK cells exert their impact across the innate and adaptive immune systems via their cytotoxicity and the secretion of cytokines. Throughout the process of immune surveillance, cytotoxic NK cells are tasked with promptly eradicating virus-infected or altered tumor cells. NK cells possess the ability to regulate immune responses through interaction with several cell types, including dendritic cells, macrophages, T cells, and endothelial cells. These interactions can either amplify or suppress immune responses [2]. There is increasing evidence indicating that NK cells, when acting as effector cells, can either promote the development of inflammation and autoimmunity or serve as a protective mechanism against their own detrimental effects [3,4]. The dysregulation of NK cells has the potential to result in an overactive inflammatory response and the emergence of autoimmune diseases [4,5]. The major histocompatibility complex class I chain-related gene A (MICA) exerts a pivotal role in orchestrating the immune response [6]. MICA serves as a significant ligand for NKG2D, facilitating the activation of NK cells and promoting reciprocal stimulation with specific subsets of T cells [7]. The production of the MICA protein is induced by DNA damage and physicochemical stressors, leading to the activation and surveillance of lymphocytes with specialized immune-related tasks. MICA enhances the capacity of the immune system to recognize and eliminate infections through the activation of NK and CD8+ T cells [6,8,9]. The emergence of graft-versus-host disease and systemic lupus erythematosus (SLE) is attributed to the processes of NK cell-mediated cytotoxicity and the production of immunosuppressive soluble MICA (sMICA) particles [10,11,12]. Autoimmune inflammatory illnesses emerge as a result of an imbalance in the regulation of immunological activation and tolerance. A recent study found that specific variants of MICA SNP haplotypes were strongly linked to a higher vulnerability to ankylosing spondylitis [13]. This finding implies MICA plays an essential part in the onset of autoimmune inflammatory illnesses. Additionally, a growing list of evidence indicates that NKG2D-positive CD4+ T cells and NK cells contribute to the pathogenesis of SLE and RA through interactions with MICA and sMICA/B [14]. Accordingly, NKG2D and NKG2D-L are important therapeutic targets in the treatment of autoimmune disorders by blocking the interaction between NKG2D of immune cells and MICA on non-immune cells. The current study sought to further evaluate the impact of MICA variants on the development of psoriasis (PSO), RA, and SLE. The data gathered in our study show that MICA*010 is a prevalent risk factor for both PSO and RA, while MICA*045 is a significant risk factor for SLE specifically in the Taiwanese population. The implications of our findings suggest that distinct MICA variants play differential roles in the development of immune-mediated inflammatory disorders (IMIDs).
2. Results
2.1. MICA*010 Is a Common Risk Factor for PSO and RA, Whereas MICA*045 Is Associated with SLE Susceptibility
In healthy Taiwanese controls, the allele frequencies of seven MICA alleles (MICA*010.01, *008.01, *002.01, *019.01, *012.01, *045, and *009.01) were greater than 0.05 (or 5%) and could be regarded as population-specific common alleles (Table 1). In the present genetic association investigation, we focused on those 7 common MICA alleles formed by 17 MICA non-synonymous coding SNPs (Supplementary Table S1). We found that MICA*010.01 was significantly associated with the risk for PSO (PFDR = 1.93 × 10−15; OR = 2.18; 95% CI = 1.82–2.62) (Table 1) and RA (PFDR = 0.0011; OR = 1.51; 95% CI = 1.18–1.93) (Table 2), suggesting that MICA*010.01 is the common risk factor for PSO and RA. In contrast, MICA*009.01 was significantly associated with protection against the development of PSO (PFDR = 0.0058; OR = 0.53; 95% CI = 0.38–0.75) (Table 1), and MICA*002.01 was significantly associated with protection against RA development (PFDR = 4.16 × 10−6; OR = 0.53; 95% CI = 0.41–0.70) (Table 2), respectively. In addition to MICA*010.01, MICA*012.01 appears to confer RA susceptibility (PFDR = 0.0330; OR = 1.41; 95% CI = 1.03–1.93) (Table 2). Notably, we identified that MICA*045 was the only allele substantially associated with SLE susceptibility (PFDR = 0.0002; OR = 2.24; 95% CI = 1.47–3.43) (Table 3). Taken together, our data indicate that the various MICA alleles have distinct roles in the development of diverse autoimmune diseases in Taiwanese people.

Table 1.
Association of MICA alleles with PSO in Taiwanese subjects.
Table 2.
Association of MICA alleles with RA in Taiwanese subjects.
Table 3.
Association of MICA alleles with SLE in Taiwanese subjects.
2.2. MICA*010 Is Deficient in Cell Surface Expression
MICA is a type I cell surface membrane protein with three extracellular domains. Two monoclonal antibodies (mAb 159,227 and 6D4) react with cell surface MICA extracellular domains. mAb 159,227 recognizes an epitope specific for MICA, while mAb 6D4 binds to a common epitope presented on both MICA and MICB. As shown in Figure 1A, MICA*008 and MICA*019 can be detected by both mAbs. MICA*008 was expressed at the highest level, while the MICA*019 expression level was intermediate among four alleles (MICA*002, *008, *010, and *019). MICA*002 was only detectable by mAb 6D4 but not by mAb 159227, suggesting that MICA*002 may function similarly to MICB. Moreover, the expression level of MICA*002 is lower than that of MICA*008 and MICA*019. Most importantly, both mAbs (159,227 and 6D4) were unable to detect MICA*010 on the cell surface as fluorescent intensities of staining were equivalent to those of parental cells and vector control cells (Figure 1A).
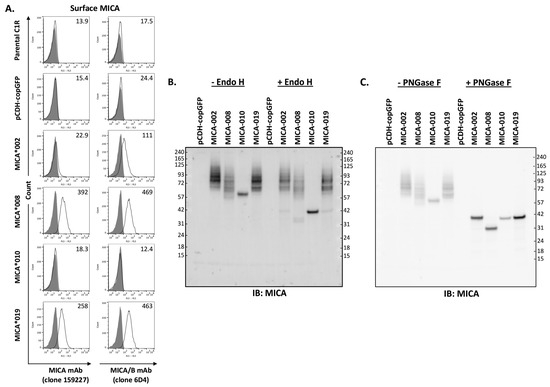
Figure 1.
Analysis of MICA allele expression. (A). Three independent FACS analyses of surface expression levels of MICA alleles with APC-conjugated anti-MICA (clone 159227, left row panels) or anti-MICA/B (clone 6D4, right row panels) monoclonal antibodies (mAbs). Anti-MICA mAb (clone 159227, R&D, FAB1300A-100) detected a low level of cell surface MICA*002 in small percentage of cells, while anti-MICA/B mAb (clone 6D4, BioLegend, 320907) detected MICA*002 in majority of cells. Both mAbs detected MICA*008 and MICA*019 with similar capacity, while they failed to detect the expression of MICA*010 on cell surface. (B). Western blot analysis of MICA allele proteins treated by Endo H. The total lysates from C1R cells expressing vector pCDH-copGFP, MICA*002, MICA*008, MICA*010, or MICA*019 were untreated (−Endo H) or treated with Endo H (+Endo H) before immunoblot with anti-MICA antibodies, as described in “Materials and Methods”. Endo H treatment led to a significant decrease in MICA*010’s molecular weight, while the treatment did not cause molecular weight changes in most MICA*002, MICA*008, and MICA*019 proteins. (C). Western blot analysis of MICA alleles treated with PNGase F. PNGase F treatment led to similar molecular weights for MICA*002, MICA*010, and MICA*019. MICA*008 without transmembrane segment and cytoplasmic domain had a smaller molecular weight than other MICA alleles after PNGase F treatment. Both Western blots (B,C) are representatives of two independent experiments.
Endo H removes high-mannose glycans from resident proteins in the endoplasmic reticulum (ER). It has been speculated that the absence of MICA*010 on the cell surface may be caused by ER retention of peptides. To examine the trafficking of MICA variants, we performed cellular MICA digestion analyses with Endo H and PNGase F. As shown in Figure 1B, MICA*010 protein’s molecular weight changed from 60 kDa to 42 kDa after Endo H and PNGase F treatment, and the MICA*010 protein was sensitive to the treatment of both Endo H and PNGase F. The sensitivity of MICA*010 to Endo H indicates that the MICA*010 peptide is trapped in ER. On the other hand, Endo H treatment did not alter the electrophoretic mobility patterns of MICA*002, *019, and *008. As shown in Figure 1C, after PNGase F treatment, the molecule weights of MICA*008, MICA*002, and MICA*019 were reduced to 36, 42, and 42 kDa, respectively, indicating that MICA*008, MICA*002, and MICA*019 are effectively processed into highly glycosylated mature MICA for cell surface expression.
2.3. MICA Alleles Have Different Binding Capacity for NKG2D
MICA is a major NKG2D ligand. Interaction between MICA on target cell surface and NKG2D on NK cells leads to NK cell activation and subsequent cytotoxicity. We used a flow cytometry-based binding assay to determine the effects of MICA alleles on their ability to interact with NKG2D. Surface staining of MICA on the transfected LCL cells was also performed and used as a normalization control. As shown in Figure 2, the binding of soluble NKG2D-Fc to MICA*002 was the highest as compared to all the other MICA alleles. The NKG2D binding capacity was similar between MICA*008 and MICA*019. MICA*010 cells failed to bind NKG2D-Fc, which could be explained by the undetectable level of cell surface MICA*010.
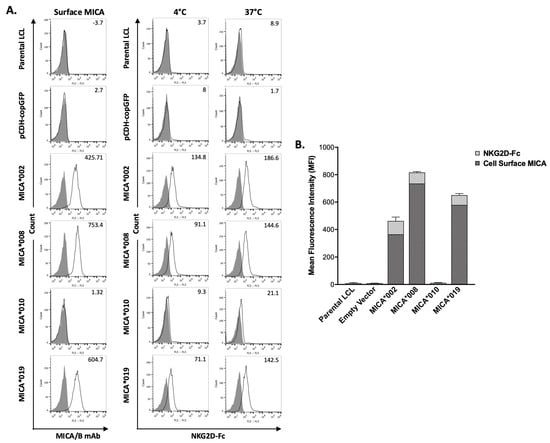
Figure 2.
Binding capacity of MICA alleles to recombinant human NKG2D-Fc fusion protein (rhNKG2D-Fc). (A). FACS analysis of the binding of MICA alleles to rhNKG2D-Fc. MICA on LCL stable cell lines expressing vector control, MICA*002, MICA*008, MICA*010, and MICA*019 were detected by anti-MICA/B (6D4) (left column panels). Binding capacity of cell lines expressing MICA alleles at 4 °C (middle column panels) and 37 °C (right column panels) was determined by flow cytometry using rhNKG2D-Fc and APC-conjugated anti-human IgG Fc, as described in “Materials and Methods”. Representative histograms of the cell surface staining of isotype controls (filled) and MICA (black lines) with the MFI values of MICA are shown (left column panels). The staining of the cells with APC-conjugated anti-human IgG Fc was used as negative control (filled). The binding of human rhNKG2D-Fc fusion protein to the cells at 4 °C (middle column panels) and 37 °C (right column panels) was detected by APC-conjugated anti-human IgG Fc (black lines). The MFI (mean fluorescent intensity) values of rhNKG2D-Fc surface staining are indicated. (B). Bar graph showing the average MFI of three independent experiments. The MFI values of MICA staining are shown with dark grey bars indicating the expression levels of different surface MICA alleles; MFI values of rhNKG2D-Fc staining are shown with light grey bars representing the binding ability of NKG2D to the respective surface MICA. MICA*008 expression level on cell surface was highest among MICA alleles. MICA*019 expression level was higher than MICA*002. MICA*002 had highest binding capacity among MICA variants. Cells expressing MICA*010 did not express surface MICA and had background staining by rhNKG2D-Fc. MICA*008 and MICA*019 had similar binding capacity at 4 °C and 37 °C.
2.4. Soluble MICA (sMICA) Levels Were Significantly Increased in Serum Samples of SLE Patients
Next, we examined the levels of sMICA in patients with SLE, RA, and psoriasis, as well as healthy controls. As depicted in Figure 3, serum sMICA concentrations were significantly higher in SLE patients than in healthy controls (p = 0.01). On the other hand, serum sMICA levels were comparable in healthy controls and in RA and psoriasis patients. Of note, we were unable to detect sMICA in RA and psoriasis patients carrying the homozygous MICA*010 allele, confirming that MICA*010 is a deficient allele for mature MICA expression.
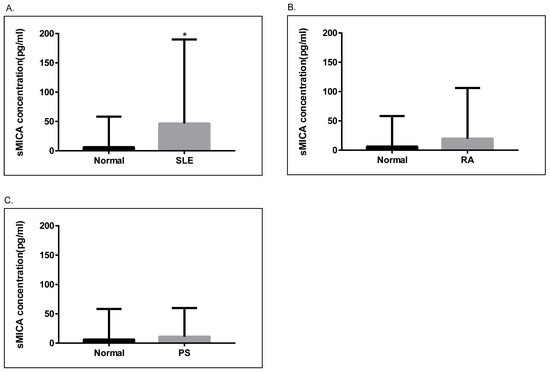
Figure 3.
Analyses of soluble MICA (sMICA) in psoriasis, RA, and SLE patients. (A). The concentrations of sMICA were significantly higher (p = 0.01) in SLE patients (N = 139, 46.66 ± 12.15 pg/mL) as compared to healthy controls (N = 92, 6.224 ± 5.417 pg/mL). (B). The concentrations of sMICA were not significantly different between healthy controls and RA patients (N = 80, 18.42 ± 10.55 pg/mL). (C). The concentrations of sMICA were not significantly different between healthy controls and psoriasis patients (N = 63, 11.08 ± 5.461 pg/mL). *, p < 0.05.
3. Discussion
MICA molecule engages C-type lectin activator receptor NKG2D on NK, γδ, and CD8+ αβ T cells to counteract the inhibiting response mediated by Killer Inhibitory Receptors (KIR) and/or CD94/NKG2A/B molecules [15]. The humoral response induced by MICA may contribute to graft rejection of solid organ transplantation [6,10,16,17,18]. MICA belongs to the non-classical class I family with highly polymorphic human stress antigens associated with tumor and inflammation surveillance [8]. A multitude of MICA functional variations have been discovered in global populations. The presence of transmembrane-encoded microsatellite triplet repeat polymorphism and coding sequence polymorphisms in MICA had a substantial impact on the expression of RNA and protein [8]. The MICA-A5.1 polymorphism results in the creation of a truncated protein that leads to greater amounts of sMICA in circulation. A substitution of valine with methionine at position 129 (MICA-129) of the MICA protein has been found to impact its capacity to bind to NKG2D, its cytotoxicity, interferon-γ release, and the density of its expression on the plasma membrane [8,19]. The MICA 129 methionine (Met) variation exhibited enhanced NKG2D signaling, eliciting greater NK-cell killing and interferon-γ release. Additionally, MICA-129Met induced rapid co-stimulation of CD8(+) T cells, resulting in a more speedy and pronounced reduction in NKG2D expression on both NK and CD8(+) T cells relative to the MICA-129 valine (Val) version [20].
Although over one hundred MICA alleles have been identified, their precise functional implications remain unknown. It is crucial to provide clarification concerning the unique distribution patterns of MICA alleles and the wide array of ethnic variations. We examined the impact of MICA alleles, which are formed by all coding SNPs, on the susceptibility to SLE, RA, and psoriasis in Taiwanese subjects. Specifically, MICA*010 was identified as a common risk factor for psoriasis and RA in Taiwanese subjects. In contrast, MICA*002, MICA*009, and MICA *012 played a protective role against the development of psoriasis. We confirmed that MICA*10 is incapable of producing a functional MICA in either soluble or membrane-bound form [13]. Furthermore, we demonstrate that the MICA*10 peptide becomes entangled in the ER, preventing its expression on the cell surface and producing sMICA in the current study. We also found that cell surface MICA*002 and MICA*010 bound to NKG2D with significantly different degrees of avidness. Our data suggest that the deficiency of MICA, a critical immune-stimulatory ligand for NKG2D, is an underlying mechanism for the susceptibility to psoriasis and RA. Functional MICA alleles are distinctively associated with the pathogenesis of psoriasis and RA in Taiwanese subjects.
Psoriasis represents an intricate autoimmune disease wherein cellular and molecular inputs to an excess immune reaction are currently identified [21]. The major histocompatibility complex (MHC) encompasses the susceptible locus for psoriasis with the greatest impact magnitude, which is influenced to some extent by variations of the HLA-Cw*0602 allele [22]. MICA is thought to be primarily stress-induced in psoriasis. Independent of HLA-B and HLA-C, the MICA-129Met allele, especially Met/Met homozygosity, had a strong correlation with the two types of cutaneous psoriasis (PsC) and psoriatic arthritis (PsA) [23]. The MICA-A9 variant, in accordance with the MICA*002 allele, is a potential genetic marker for the occurrence of PsA, irrespective of the HLA-C association [24,25]. Psoriasis was associated with SNPs with potent immunological function, namely, rs2507971, rs9260313, rs66609536, and rs380924, located in the class I region of the MHC [22]. SNP rs13437088, situated at the 16 kb telomeric region of MICA and 30 kb centromeric region of HLA-B, exhibited a robust association with Han Chinese psoriasis after adjusting the effect of the imputed HLA-Cw*0602 [26]. Furthermore, the prevalence of the MICA-A5.1 allele was substantially greater among Chinese subjects [27]. Risk alleles for HLA-B and HLA-C determine susceptibility to autoimmunity and inflammation. The MICA*010-HLA-B*4601-Cw*01 haplotype displayed a greater incidence of Type I and II psoriasis. Significantly, MICA*010 exhibited a robust correlation with HLA-Cw*01 [28], which correspond to two prominent risk loci in the Asian population, including Taiwanese subjects [29,30].
RA was associated with MICA-129Val, and RA patients had elevated sMICA levels [31]. In addition, MICA polymorphisms are associated with HLA-DRB1 shared epitope (SE) presence and autoantibody production in RA patients [32]. Support for the correlation of MICA-250 (rs1051794) with RA occurred independently of the recognized HLA-DRB1 risk alleles, suggesting that MICA is a susceptible gene [33,34]. The MICA rs1051792 polymorphism modifies the therapeutic response of RA patients to TNF-blocking therapy [35]. In the Tunisian population, the MICA-A9 allele was substantially associated with RF-negative RA patients, while the severity of RA may be influenced by the MICA-TM and MICA 129met/val genotypes [33]. The correlation observed in Koreans between the MICA-A4 allele and RA was found to be associated with HLA-DRB1*0405, whereas the MICA-A9 allele could possess a mild protective role toward RA susceptibility [36]. The MICA-A6 allele was an independent predictor of RA protection [37] in the SE+ subset of RA patients [38]. The functional confirmation of MICA*010 as a risk factor was achieved through deficient MICA expression, whereas MICA*002.01, characterized by distinct expression levels, exhibited a significant association with protection against the development of RA. Genetic variations within the MICA collectively contribute to the pathogenesis of RA.
Membrane MICA and sMICA are crucial for regulating the immune response to stimuli. Membrane-expressed MICA protein promotes the activation of NK and T cells. NK cell-mediated cytotoxicity and immunosuppressive sMICA could down-regulate NKG2D expression and encourage immunosuppressive CD4+ T-cell proliferation [39]. NKG2D-MICA engagement in SLE patients may initiate the mutual growth of MICA+ monocytes and NKG2D+CD4+ T cells [40]. However, sMICA promotes the proliferation of immunosuppressive NKG2D+CD4+ T cells, which correlates inversely with SLE disease activity [41]. A particular category of lupus individuals who had low levels of vitamin D, innate T-cell activation, and nephropathy had elevated levels of circulating sMICA [42], which were also substantially elevated among those with anti-SSB and anti-RNP autoantibodies [12]. Contact across MICA and NKG2D initiates NK cell activation and upregulation of CD69 and CD107 on NK cells, which contributes to NK cell exhaustion in SLE patients [12]. MICA and NKG2D have been identified as cytotoxic factors in cutaneous SLE [43]. The MICA-129Met/Val dimorphism influences MICA expression and the release of proteolytic sMICA [19]. Speedy NKG2D reduction on alloreactive CD8+T cells suggested that activation of MICA-129Met diminished the severity of acute graft-versus-host disease [20]. The combination of two of the three main markers (DR3-DQ2, MICA-5, and MICA-5.1) was related to an upsurge in genetic susceptibility and a reduced risk between MICA-9 and SLE [44], whereas the association between MICA-5.1 and SLE is owing to its disequilibrium of linkage with HLA B*08 [45]. HLA DRB1 *03, in conjunction with MICA-A5.1 and the absence of the two MICA-A6 and HLA DRB*11 alleles, is strongly associated with SLE [46]. The MICA 129Met allele, MICA-A9 allele, and 129Met/Met genotype were all associated with SLE susceptibility, and the combination of 129Met/A-9 variants displayed reduced expression of NKG2D and NK cell cytotoxicity but increased IFN-γ production from NK92MI cells [47]. Han Chinese with MICA*010 alleles had a reduced incidence of SLE [48]. The present study identified the MICA*045 allele as the sole risk factor for SLE, while no association was observed between the MICA*010 allele and SLE in Taiwanese subjects. The presence of 129Met in MICA*045 (Supplementary Table S1), which mediates stronger NKG2D signals than 129Val, may partially explain the association between MICA*045 and SLE. Nonetheless, the intricate MICA-NKG2D signaling pathways in various autoimmune inflammatory diseases have yet to be uncovered.
Our study has limitations. The genetic interactions between MICA and HLA in the pathogenesis of autoimmune inflammatory diseases, particularly HLA-C in PSO and HLA-DRB1 in RA, are an essential consideration [7]. Further, the effects of MICA alleles on modifications in serology tests, such as anti-cyclic citrullinated peptide (anti-CCP) positivity and clinical manifestations of autoimmune diseases, were not analyzed. Finally, we did not determine whether MICA alleles could affect outcomes of function-based therapies (especially biologic pharmaceuticals) in the treatment of PSO and RA [35,49].
4. Materials and Methods
4.1. Study Subjects
Patients with psoriasis (N = 579), RA (N = 604), and SLE (N = 682) were recruited at the Chang Gung Memorial Hospital Rheumatology Clinic. Patients with psoriasis, RA, and SLE were diagnosed according to the respective diagnostic criteria for psoriasis, RA, and SLE. In the same geographical region, 928 healthy blood donors of similar age and gender were recruited as controls. The Chang Gung Memorial Hospital Institutional Review Board (Protocol No. 20200043B0) approved the human study, and all participants provided written consent for the study.
4.2. Determination of MICA Alleles by DNA Sequence Analysis
The sense primer (5′-CAA GAC CTT CCT TCC ACC ACC T-3′) and antisense primer (5′-CCT TGT CAC CAA CAT GCC TAT CTT T-3′) were utilized to amplify the MICA gene-specific DNA fragment (2352 base pairs) containing exons 2, 3, 4, and 5. Three sequencing primers (5′-CAG CAG ACC TGT GTG TTA A-3′, 5′-GGT GAT GGG TTC GGG AA-3′, and 5′-TTC CTC TCC CCT CCT TAG A-3′) and BigDye v3.1 Sequencing kit (Applied Biosystems, Foster City, CA, USA) were utilized in DNA sequence analyses of MICA exon 2, 3, and 4 on an ABI 3730xl DNA Analyzer. MICA DNA sequences were determined and MICA haplotypes or alleles were assigned according to IMGT/HLA database (ftp://ftp.ebi.ac.uk/pub/databases/ipd/imgt/hla/fasta/MICA_nuc.fasta, release date: 2022, accessed on 1 February 2023).
4.3. Construction of MICA Gene Expression Constructs
MICA cDNAs of peripheral blood mononuclear cells from the carriers of MICA alleles (MICA*002, MICA*008, MICA*010, or MICA*019) were amplified with RT-PCR and cloned into the lentiviral vector pCDH-CMV-EF1-copGFP (Systems Biosciences, Mountain View, CA, USA), as previously described [13].
4.4. Generation of Stable Cell Lines Expressing MICA Alleles
The C1R (ATCC#CRL-1573, Manassas, VA, USA) and LCL-721.221 (ATCC#CRL-1855) cell lines expressing empty vector, MICA*002, MICA*008, MICA*010, or MICA*019 alleles were gated as copGFP-positive and sorted on vFACSAria III cytometer (BD Biosciences, Mountain View, CA, USA), as previously described [13]. Stable cell lines were maintained in DMEM medium supplemented with 10% fetal calf serum and 1% GlutaMax (Invitrogen, Carlsbad, CA, USA) and incubated at 37 °C and 5% CO2. Prior to use, copGFP expressions were monitored by FACS.
4.5. Analysis of Membrane-Bound MICA by Flow Cytometry
To determine surface expression of MICA alleles, the parental C1R and LCL-721.221 cells along with their respective stable clones expressing empty vector, MICA*002, MICA*008, MICA*010, and MICA*019 were stained separately with APC-conjugated anti-human MICA (clone 159227, R&D, Minneapolis, MN, USA) and MICA/MICB (clone 6D4, Biolegend, San Diego, CA, USA) for 30 min at 4 °C, followed by FACS analysis on an EC800 Flow Cytometry Analyzer (Sony Biotechnology, San Jose, CA USA). Transduced cells positive for GFP were gated for the determination of membrane-bound MICA.
4.6. Determination of MICA Trafficking
Total lysates of cells expressing MICA alleles were solubilized for 30 min on ice in lysis buffer (1% Triton X-100, 50 mM of Tris-Cl, pH 7.4, 300 mM of NaCl, 5 mM of EDTA, 0.02% NaN3) containing complete protease inhibitors (Roche Applied Science, Upper Bavaria, Germany). To investigate the trafficking of MICA alleles, the total cell lysates were deglycosylated with Endo H and PNGase F according to the manufacturer’s instructions (New England Biolabs, Ipswich, MA, USA). After the treatment, the samples were separated on 4–12% gradient SDS-PAGE, transferred to PVDF membranes (GE Healthcare, Uppsala, Sweden), and detected by rabbit anti-human MICA antibody (Abcam, Cambridge, UK) according to Methods section, as previously described [13].
4.7. NKG2D Binding Assay
LCL stable cell lines expressing vector control, MICA*002, MICA*008, MICA*010, and MICA*019 were incubated with recombinant human NKG2D-Fc fusion protein (R&D) at 4 °C or 37 °C, followed by staining with APC-conjugated anti-human IgG Fc. MICA surface expression was monitored by staining with anti-MICA/B (6D4, Biolegend) at 4 °C for 30 min, as described above. The binding capacity of rhNKG2D-Fc to MICA alleles was analyzed by EC800 Flow Cytometry Analyzer (Sony Biotechnology).
4.8. Determination of Soluble MICA (sMICA) Concentrations in Human Serum Samples
Concentrations of sMICA in serum samples from patients (psoriasis, RA, and SLE) and normal healthy controls were determined using a sandwich MICA DuoSet ELISA kit (R&D Systems, Minneapolis, MN, USA), according to the manufacturer’s instructions. The sMICA concentrations were calculated based on the standards from the same ELISA plate. Results were reported as the concentration of sMICA (pg/mL) in the serum sample.
4.9. Statistical Analysis
Logistic regression models were used to analyze relationship between the estimated haplotypes (alleles) and disease susceptibility. The 1% level of significance for p-values (p < 0.01) was used in genetic association analyses. Unpaired t-tests of GraphPad Prism 6.0 (GraphPad, La Jolla, CA, USA) were used to compare serum sMICA concentrations between normal healthy controls and PSA, RA, and SLE patients, respectively. The 5% level of significance (p < 0.05) was used for sMICA analysis.
5. Conclusions
Important functions are played by MICA variants in autoimmune inflammatory diseases. The MICA*002 allele with high avidity protects against psoriasis and RA, whereas the deficient MICA*010 allele increases the risk for psoriasis and RA development. The MICA*045 containing high activity 129Met is associated with SLE susceptibility. The mechanisms underlying the effects of MICA alleles on the pathogenesis of autoimmune diseases may involve differences in MICA expression levels and avidity for NKG2D among MICA alleles. SLE disease activity might be affected by the elevated sMICA levels, which are influenced by the MICA alleles. Our data demonstrate that MICA variants impact the pathogenesis of common autoimmune inflammatory diseases, including PSO, RA, and SLE. The determination of MICA alleles could facilitate the diagnosis of autoimmune diathesis.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms25053036/s1.
Author Contributions
Conceptualization, resources, funding acquisition: C.-M.W., J.W. and J.-Y.C. Methodology, data curation, investigation: Y.-J.J.W., K.-P.T. and J.-W.Z. Original draft preparation: C.-M.W., K.-P.T., J.W., J.-Y.C. and Y.-J.J.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Chang Gung Memorial Hospital (grant numbers: CMRPG 5K0131, CMRPG 5H0022, CMRPG 5K0042, and CMRPG 3J1422) and the Ministry of Science and Technology, Taiwan (grant numbers: MOST 109-2314-B-182-068-MY3 and 112-2314-B-182-057).
Institutional Review Board Statement
This study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board of Chang Gung Memorial Hospital (protocol code No. 20200043B0 approved on 18 April 2020).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We greatly appreciate the Shin Chu Blood Donor Center for the collection of samples.
Conflicts of Interest
The authors declare no conflicts of interest. This study was carried out in accordance with the relevant guidelines and regulations. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; and in the decision to publish the results.
References
- Paul, S.; Lal, G. The Molecular Mechanism of Natural Killer Cells Function and Its Importance in Cancer Immunotherapy. Front. Immunol. 2017, 8, 1124. [Google Scholar] [CrossRef]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Velardi, A. Natural killer cell alloreactivity 10 years later. Curr. Opin. Hematol. 2012, 19, 421–426. [Google Scholar] [CrossRef]
- Zitti, B.; Bryceson, Y.T. Natural killer cells in inflammation and autoimmunity. Cytokine Growth Factor Rev. 2018, 42, 37–46. [Google Scholar] [CrossRef]
- Kucuksezer, U.C.; Aktas Cetin, E.; Esen, F.; Tahrali, I.; Akdeniz, N.; Gelmez, M.Y.; Deniz, G. The Role of Natural Killer Cells in Autoimmune Diseases. Front. Immunol. 2021, 12, 622306. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Stastny, P. Role of MICA in the immune response to transplants. Tissue Antigens 2010, 76, 171–176. [Google Scholar] [CrossRef]
- Tchacrome, I.; Zhu, Q.; Saleh, M.A.; Zou, Y. Diseases association with the polymorphic major histocompatibility complex class I related chain a: MICA gene. Transpl. Immunol. 2022, 75, 101665. [Google Scholar] [CrossRef]
- Choy, M.K.; Phipps, M.E. MICA polymorphism: Biology and importance in immunity and disease. Trends Mol. Med. 2010, 16, 97–106. [Google Scholar] [CrossRef]
- Duan, S.; Guo, W.; Xu, Z.; He, Y.; Liang, C.; Mo, Y.; Wang, Y.; Xiong, F.; Guo, C.; Li, Y.; et al. Natural killer group 2D receptor and its ligands in cancer immune escape. Mol. Cancer 2019, 18, 29. [Google Scholar] [CrossRef]
- Boukouaci, W.; Busson, M.; Peffault de Latour, R.; Rocha, V.; Suberbielle, C.; Bengoufa, D.; Dulphy, N.; Haas, P.; Scieux, C.; Amroun, H.; et al. MICA-129 genotype, soluble MICA, and anti-MICA antibodies as biomarkers of chronic graft-versus-host disease. Blood 2009, 114, 5216–5224. [Google Scholar] [CrossRef]
- Boukouaci, W.; Al-Daccak, R.; Dulphy, N.; Lauden, L.; Amokrane, K.; Fortier, C.; Marzais, F.; Bennabi, M.; Peffault de Latour, R.; Socie, G.; et al. Soluble MICA-NKG2D interaction upregulates IFN-γ production by activated CD3-CD56+ NK cells: Potential impact on chronic graft versus host disease. Hum. Immunol. 2013, 74, 1536–1541. [Google Scholar] [CrossRef]
- Hervier, B.; Ribon, M.; Tarantino, N.; Mussard, J.; Breckler, M.; Vieillard, V.; Amoura, Z.; Steinle, A.; Klein, R.; Kötter, I.; et al. Increased Concentrations of Circulating Soluble MHC Class I-Related Chain A (sMICA) and sMICB and Modulation of Plasma Membrane MICA Expression: Potential Mechanisms and Correlation With Natural Killer Cell Activity in Systemic Lupus Erythematosus. Front. Immunol. 2021, 12, 633658. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.M.; Tan, K.P.; Jan Wu, Y.J.; Lin, J.C.; Zheng, J.W.; Yu, A.L.; Wu, J.M.; Chen, J.Y. MICA*019 Allele and Soluble MICA as Biomarkers for Ankylosing Spondylitis in Taiwanese. J. Pers. Med. 2021, 11, 564. [Google Scholar] [CrossRef]
- Wei, L.; Xiang, Z.; Zou, Y. The Role of NKG2D and Its Ligands in Autoimmune Diseases: New Targets for Immunotherapy. Int. J. Mol. Sci. 2023, 24, 17545. [Google Scholar] [CrossRef]
- Bauer, S.; Groh, V.; Wu, J.; Steinle, A.; Phillips, J.H.; Lanier, L.L.; Spies, T. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science 1999, 285, 727–729. [Google Scholar] [CrossRef] [PubMed]
- Pabon, M.A.; Navarro, C.E.; Martin, R.; Rodriguez, M.; Martin, I.; Gaitan, L.; Gomez, A.; Lozano, E. Minor histocompatibility antigens as risk factor for poor prognosis in kidney transplantation. Transplant. Proc. 2011, 43, 3319–3323. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Alvarez, B.; Alonso-Arias, R.; Bravo-Mendoza, C.; Lopez-Vazquez, A.; Ortega, T.; Baltar, J.M.; Coto, E.; Ortega, F.; Lopez-Larrea, C. Identification of epitopes and immunodominant regions on the MICA protein defined by alloantibodies from kidney transplant patients. Transplantation 2009, 88 (Suppl. S3), S68–S77. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.T.; Stephens, H.A.; Fernando, R.; Karasu, A.; Harber, M.; Howie, A.J.; Powis, S.; Zou, Y.; Stastny, P.; Madrigal, J.A.; et al. Major histocompatibility complex class I-related chain A allele mismatching, antibodies, and rejection in renal transplantation. Hum. Immunol. 2011, 72, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Isernhagen, A.; Schilling, D.; Monecke, S.; Shah, P.; Elsner, L.; Walter, L.; Multhoff, G.; Dressel, R. The MICA-129Met/Val dimorphism affects plasma membrane expression and shedding of the NKG2D ligand MICA. Immunogenetics 2016, 68, 109–123. [Google Scholar] [CrossRef]
- Isernhagen, A.; Malzahn, D.; Viktorova, E.; Elsner, L.; Monecke, S.; von Bonin, F.; Kilisch, M.; Wermuth, J.M.; Walther, N.; Balavarca, Y.; et al. The MICA-129 dimorphism affects NKG2D signaling and outcome of hematopoietic stem cell transplantation. EMBO Mol. Med. 2015, 7, 1480–1502. [Google Scholar] [CrossRef]
- Harden, J.L.; Krueger, J.G.; Bowcock, A.M. The immunogenetics of Psoriasis: A comprehensive review. J. Autoimmun. 2015, 64, 66–73. [Google Scholar] [CrossRef]
- Knight, J.; Spain, S.L.; Capon, F.; Hayday, A.; Nestle, F.O.; Clop, A.; Barker, J.N.; Weale, M.E.; Trembath, R.C. Conditional analysis identifies three novel major histocompatibility complex loci associated with psoriasis. Hum. Mol. Genet. 2012, 21, 5185–5192. [Google Scholar] [CrossRef]
- Pollock, R.A.; Chandran, V.; Pellett, F.J.; Thavaneswaran, A.; Eder, L.; Barrett, J.; Rahman, P.; Farewell, V.; Gladman, D.D. The functional MICA-129 polymorphism is associated with skin but not joint manifestations of psoriatic disease independently of HLA-B and HLA-C. Tissue Antigens 2013, 82, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, S.; Martinez-Borra, J.; Torre-Alonso, J.C.; Gonzalez-Roces, S.; Sanchez del Río, J.; Rodriguez Pérez, A.; Brautbar, C.; López-Larrea, C. The MICA-A9 triplet repeat polymorphism in the transmembrane region confers additional susceptibility to the development of psoriatic arthritis and is independent of the association of Cw*0602 in psoriasis. Arthritis Rheum. 1999, 42, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- González, S.; Martínez-Borra, J.; López-Vázquez, A.; García-Fernández, S.; Torre-Alonso, J.C.; López-Larrea, C. MICA rather than MICB, TNFA, or HLA-DRB1 is associated with susceptibility to psoriatic arthritis. J. Rheumatol. 2002, 29, 973–978. [Google Scholar] [PubMed]
- Feng, B.J.; Sun, L.D.; Soltani-Arabshahi, R.; Bowcock, A.M.; Nair, R.P.; Stuart, P.; Elder, J.T.; Schrodi, S.J.; Begovich, A.B.; Abecasis, G.R.; et al. Multiple Loci within the major histocompatibility complex confer risk of psoriasis. PLoS Genet. 2009, 5, e1000606. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Zhang, S.Z.; Xiao, C.Y.; Hou, Y.P.; Li, L.; Luo, H.C.; Jiang, H.Y.; Zuo, W.Q. The A5.1 allele of the major histocompatibility complex class I chain-related gene A is associated with psoriasis vulgaris in Chinese. Br. J. Dermatol. 2000, 143, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Romphruk, A.V.; Romphruk, A.; Choonhakarn, C.; Puapairoj, C.; Inoko, H.; Leelayuwat, C. Major histocompatibility complex class I chain-related gene A in Thai psoriasis patients: MICA association as a part of human leukocyte antigen-B-Cw haplotypes. Tissue Antigens 2004, 63, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.W.; Tsai, T.F. HLA-Cw1 and Psoriasis. Am. J. Clin. Dermatol. 2021, 22, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.S.; Tsai, T.F. Associations between HLA-Cw1 and Systemic Treatment Response of Asian Psoriasis Patients. Mol. Diagn. Ther. 2022, 26, 541–549. [Google Scholar] [CrossRef]
- Mariaselvam, C.M.; Boukouaci, W.; Charron, D.; Krishnamoorthy, R.; Tamouza, R.; Misra, D.P.; Negi, V.S. Association of MICA-129 polymorphism and circulating soluble MICA level with rheumatoid arthritis in a south Indian Tamil population. Int. J. Rheum. Dis. 2018, 21, 656–663. [Google Scholar] [CrossRef]
- Wielińska, J.; Tarassi, K.; Iwaszko, M.; Kościńska, K.; Wysoczańska, B.; Mole, E.; Kitsiou, V.; Świerkot, J.; Kolossa, K.; Kouniaki, D.; et al. Shared epitope and polymorphism of MICA and NKG2D encoding genes in Greek and Polish patients with rheumatoid arthritis. Cent.-Eur. J. Immunol. 2021, 46, 92–98. [Google Scholar] [CrossRef]
- Achour, Y.; Kammoun, A.; Ben Hamad, M.; Mahfoudh, N.; Chaabane, S.; Marzouk, S.; Keskes, L.; Gaddour, L.; Bahloul, Z.; Maalej, A. Association study of MICA gene polymorphisms with rheumatoid arthritis susceptibility in south Tunisian population. Int. J. Immunogenet. 2014, 41, 486–492. [Google Scholar] [CrossRef]
- Kirsten, H.; Petit-Teixeira, E.; Scholz, M.; Hasenclever, D.; Hantmann, H.; Heider, D.; Wagner, U.; Sack, U.; Hugo Teixeira, V.; Prum, B.; et al. Association of MICA with rheumatoid arthritis independent of known HLA-DRB1 risk alleles in a family-based and a case control study. Arthritis Res. Ther. 2009, 11, R60. [Google Scholar] [CrossRef]
- Iwaszko, M.; Świerkot, J.; Dratwa, M.; Wysoczańska, B.; Korman, L.; Bugaj, B.; Kolossa, K.; Jeka, S.; Wiland, P.; Bogunia-Kubik, K. Association of MICA-129Met/Val polymorphism with clinical outcome of anti-TNF therapy and MICA serum levels in patients with rheumatoid arthritis. Pharmacogenomics J. 2020, 20, 760–769. [Google Scholar] [CrossRef]
- Mok, J.W.; Lee, Y.J.; Kim, J.Y.; Lee, E.B.; Song, Y.W.; Park, M.H.; Park, K.S. Association of MICA polymorphism with rheumatoid arthritis patients in Koreans. Hum. Immunol. 2003, 64, 1190–1194. [Google Scholar] [CrossRef]
- Singal, D.P.; Li, J.; Zhang, G. Microsatellite polymorphism of the MICA gene and susceptibility to rheumatoid arthritis. Clin. Exp. Rheumatol. 2001, 19, 451–452. [Google Scholar]
- Martinez, A.; Fernandez-Arquero, M.; Balsa, A.; Rubio, A.; Alves, H.; Pascual-Salcedo, D.; Martin-Mola, E.; de la Concha, E.G. Primary association of a MICA allele with protection against rheumatoid arthritis. Arthritis Rheum. 2001, 44, 1261–1265. [Google Scholar] [CrossRef]
- Molfetta, R.; Quatrini, L.; Santoni, A.; Paolini, R. Regulation of NKG2D-Dependent NK Cell Functions: The Yin and the Yang of Receptor Endocytosis. Int. J. Mol. Sci. 2017, 18, 1677. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wang, H.; Ni, B.; He, Y.; Li, J.; Tang, Y.; Fu, X.; Wang, Q.; Xu, G.; Li, K.; et al. Mutual activation of CD4+ T cells and monocytes mediated by NKG2D-MIC interaction requires IFN-gamma production in systemic lupus erythematosus. Mol. Immunol. 2009, 46, 1432–1442. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Turtle, C.J.; Booth, G.C.; Riddell, S.R.; Gooley, T.A.; Stevens, A.M.; Spies, T.; Groh, V. Normally occurring NKG2D+CD4+ T cells are immunosuppressive and inversely correlated with disease activity in juvenile-onset lupus. J. Exp. Med. 2009, 206, 793–805. [Google Scholar] [CrossRef]
- Pérez-Ferro, M.; Romero-Bueno, F.I.; Serrano Del Castillo, C.; Mahillo, I.; Alvear, A.; Largo, R.; Herrero-Beaumont, G.; Sánchez-Pernaute, O. A subgroup of lupus patients with nephritis, innate T cell activation and low vitamin D is identified by the enhancement of circulating MHC class I-related chain A. Clin. Exp. Immunol. 2019, 196, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Vorwerk, G.; Zahn, S.; Bieber, T.; Wenzel, J. NKG2D and its ligands as cytotoxic factors in cutaneous lupus erythematosus. Exp. Dermatol. 2021, 30, 847–852. [Google Scholar] [CrossRef]
- Gambelunghe, G.; Gerli, R.; Bocci, E.B.; Del Sindaco, P.; Ghaderi, M.; Sanjeevi, C.B.; Bistoni, O.; Bini, V.; Falorni, A. Contribution of MHC class I chain-related A (MICA) gene polymorphism to genetic susceptibility for systemic lupus erythematosus. Rheumatology 2005, 44, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Torres, B.; Vilches, J.R.; López-Nevot, M.A.; Ortego-Centeno, N.; Jiménez-Alonso, J.; González-Gay, M.A.; de Ramón, E.; Sánchez-Román, J.; Núñez-Roldán, A.; et al. No primary association of MICA polymorphism with systemic lupus erythematosus. Rheumatology 2006, 45, 1096–1100. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fojtíková, M.; Novota, P.; Cejková, P.; Pešičková, S.; Tegzová, D.; Cerná, M. HLA class II, MICA and PRL gene polymorphisms: The common contribution to the systemic lupus erythematosus development in Czech population. Rheumatol. Int. 2011, 31, 1195–1201. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yoshida, K.; Komai, K.; Shiozawa, K.; Mashida, A.; Horiuchi, T.; Tanaka, Y.; Nose, M.; Hashiramoto, A.; Shiozawa, S. Role of the MICA polymorphism in systemic lupus erythematosus. Arthritis Rheum. 2011, 63, 3058–3066. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Zhu, Q.; Chen, C.; Fu, X.; Li, Y.; Liu, L.; Luo, Q.; Wang, F.; Wang, Y. Association Between Major Histocompatibility Complex Class I Chain-Related Gene Polymorphisms and Susceptibility of Systemic Lupus Erythematosus. Am. J. Med. Sci. 2017, 354, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Morelli, M.; Galluzzo, M.; Scarponi, C.; Madonna, S.; Scaglione, G.L.; Girolomoni, G.; Talamonti, M.; Bianchi, L.; Albanesi, C. Allelic Variants of HLA-C Upstream Region, PSORS1C3, MICA, TNFA and Genes Involved in Epidermal Homeostasis and Barrier Function Influence the Clinical Response to Anti-IL-12/IL-23 Treatment of Patients with Psoriasis. Vaccines 2022, 10, 1977. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).