
Astragaloside IV as a Memory-Enhancing Agent: In Silico Studies with In Vivo Analysis and Post Mortem ADME-Tox Profiling in Mice



Abstract
1. Introduction
2. Results
2.1. The ADME-Tox Studies
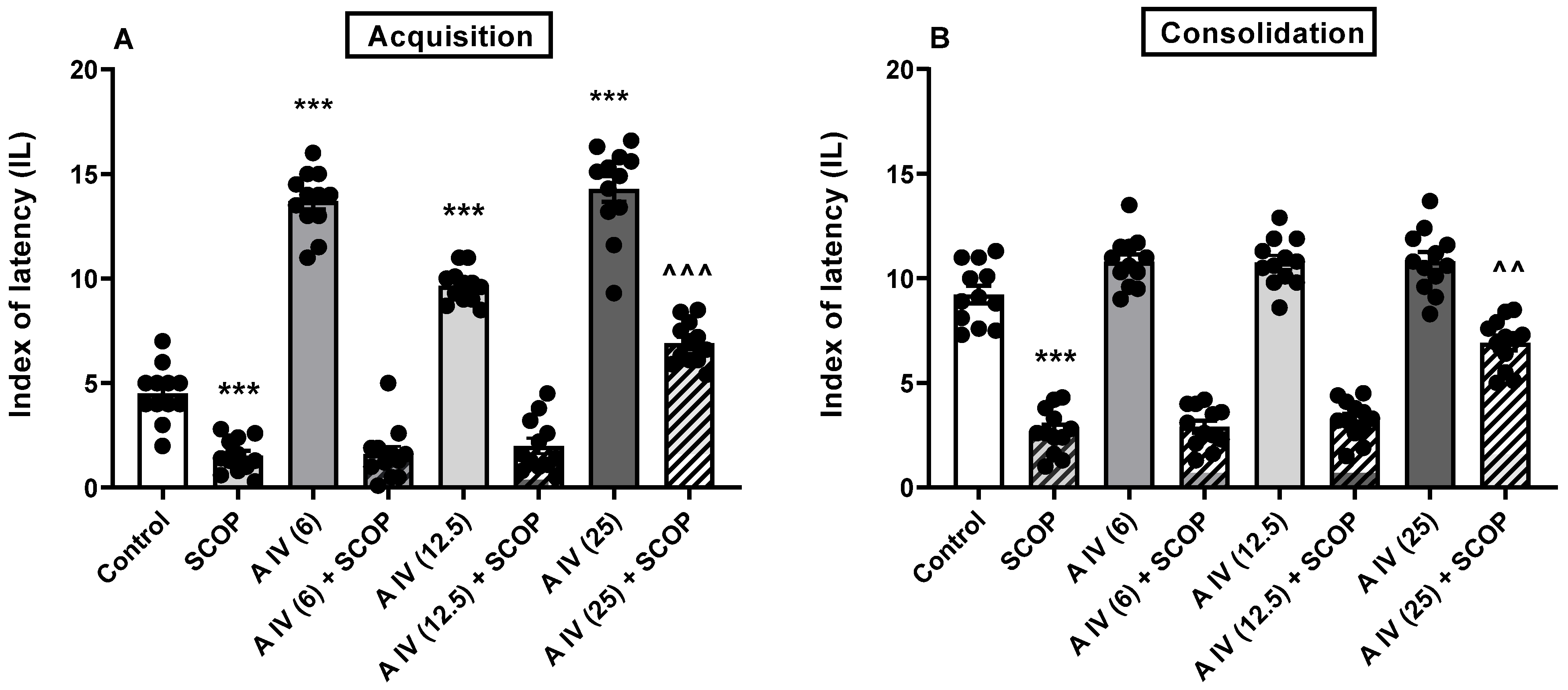
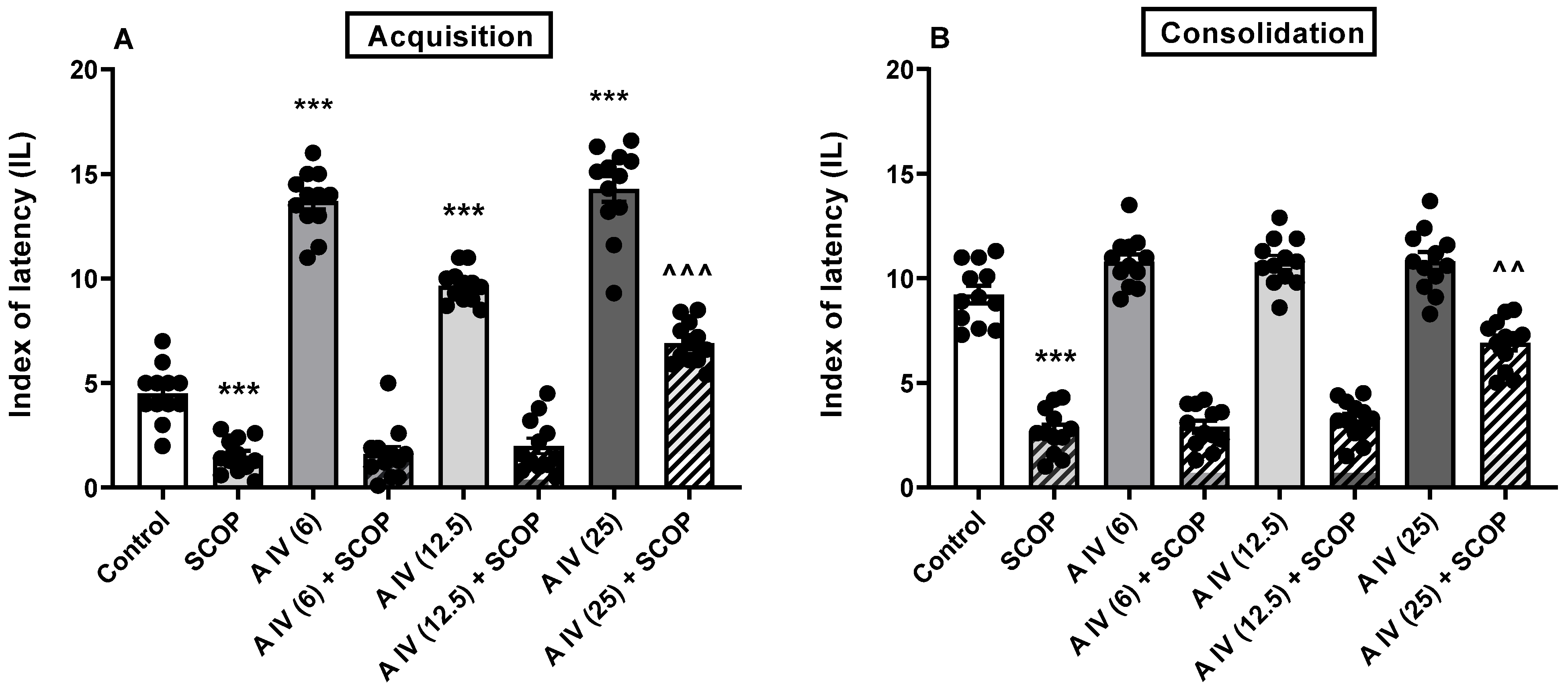
2.2. The Influence of Astragaloside IV on the Acquisition and Consolidation of Long-Term Memory Impairment Induced by SCOP Administration in Mice
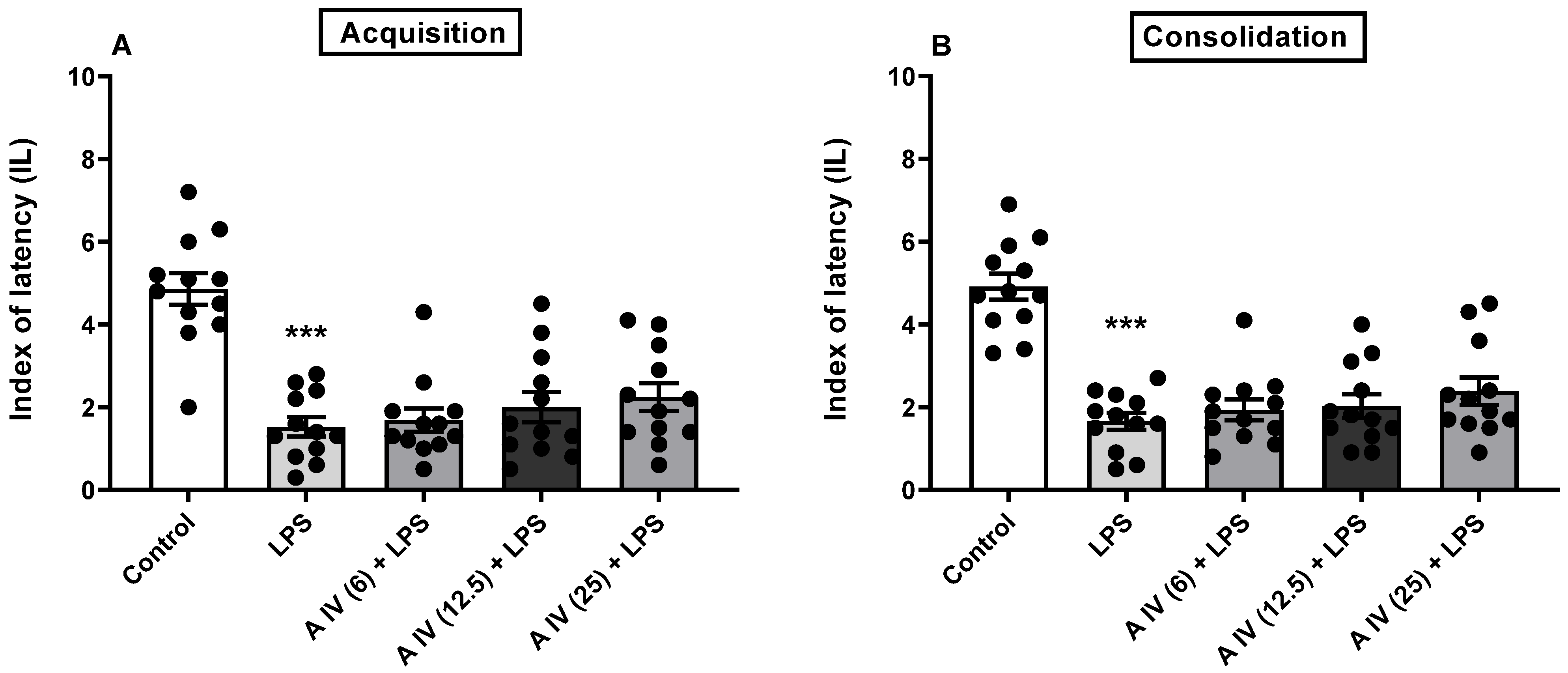
2.3. Influence of Astragaloside IV on the Acquisition and Consolidation of Long-Term Memory Impairments Induced by LPS Administration in Mice
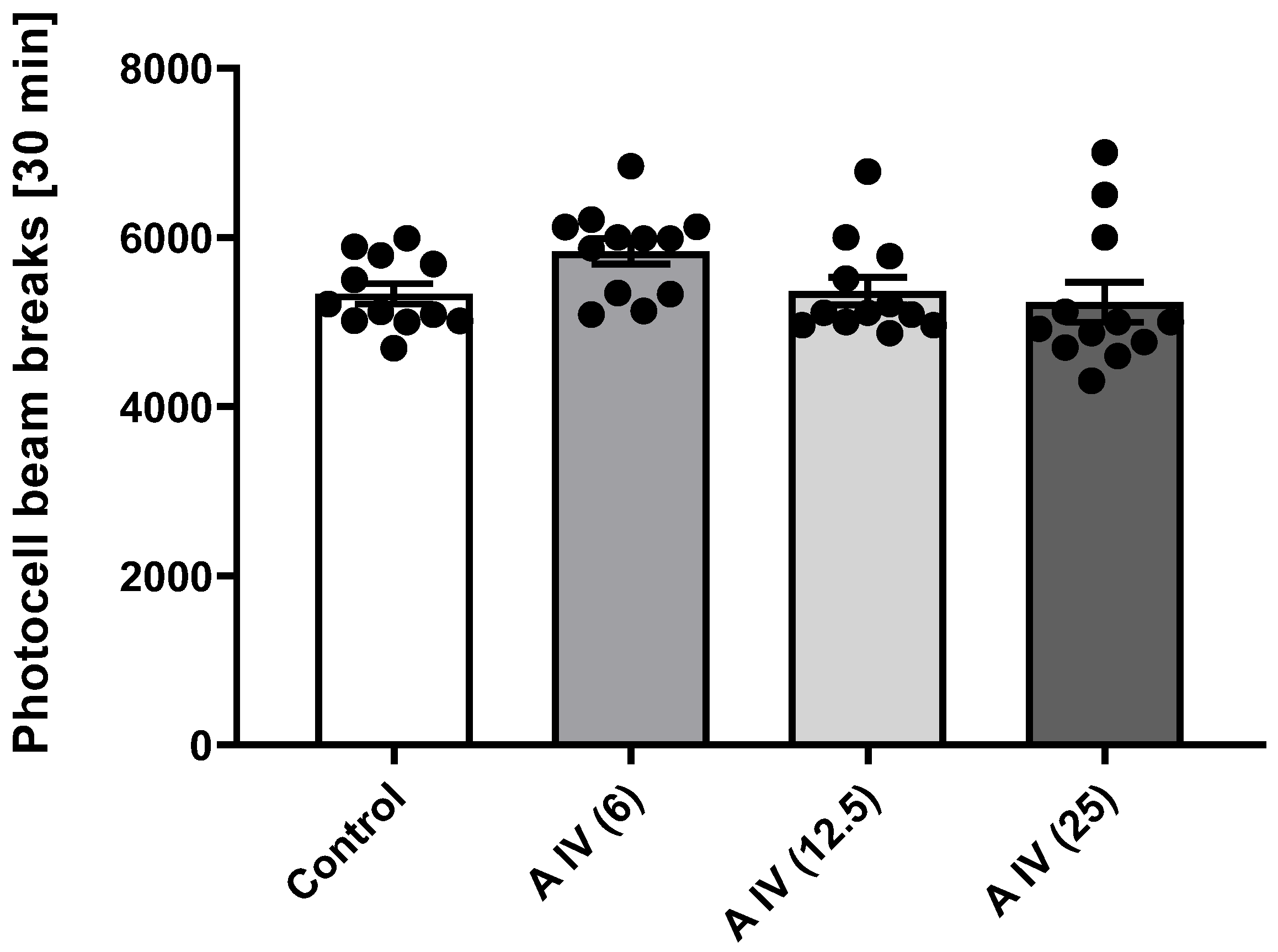
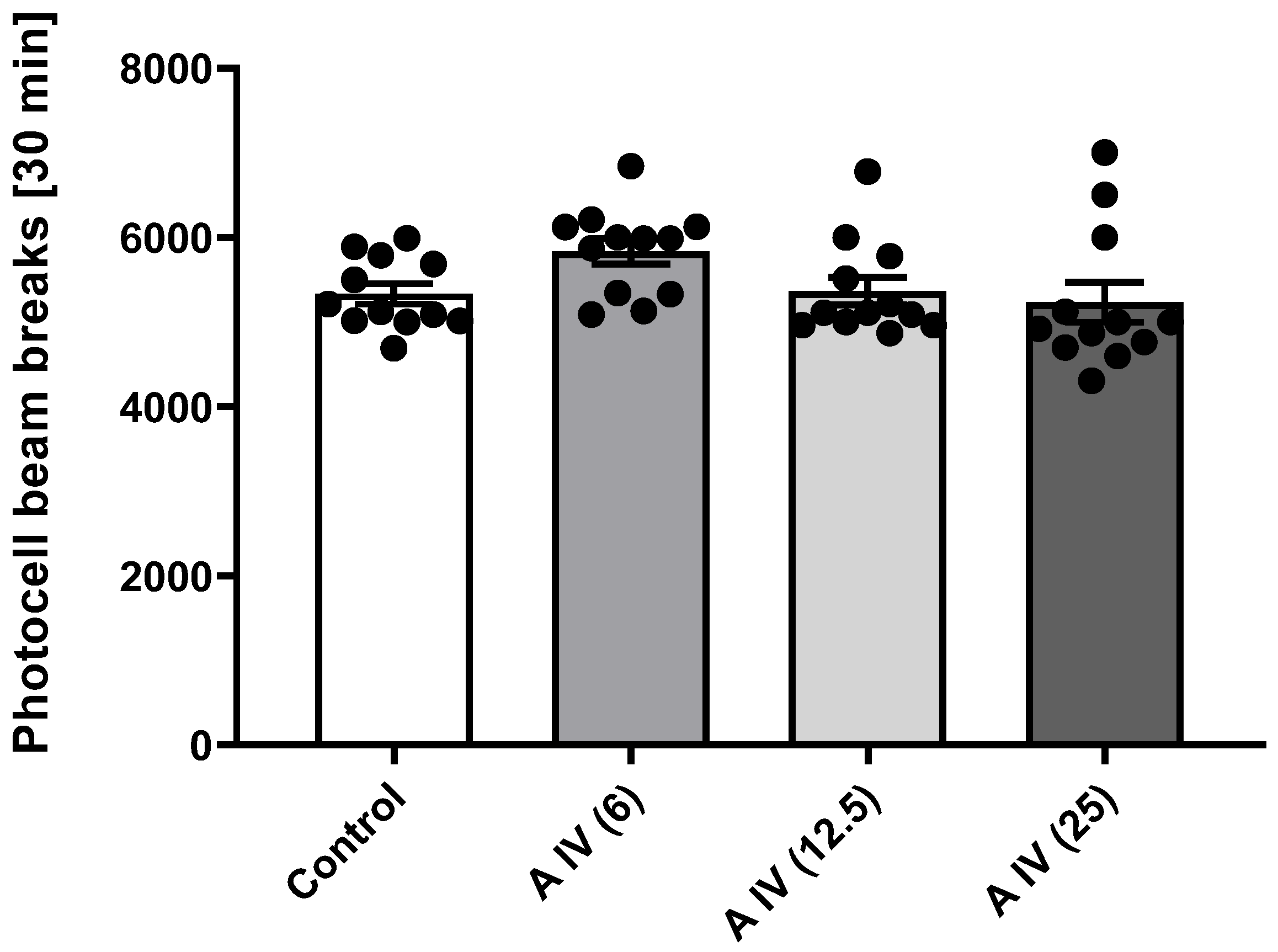
2.4. Locomotor Activity
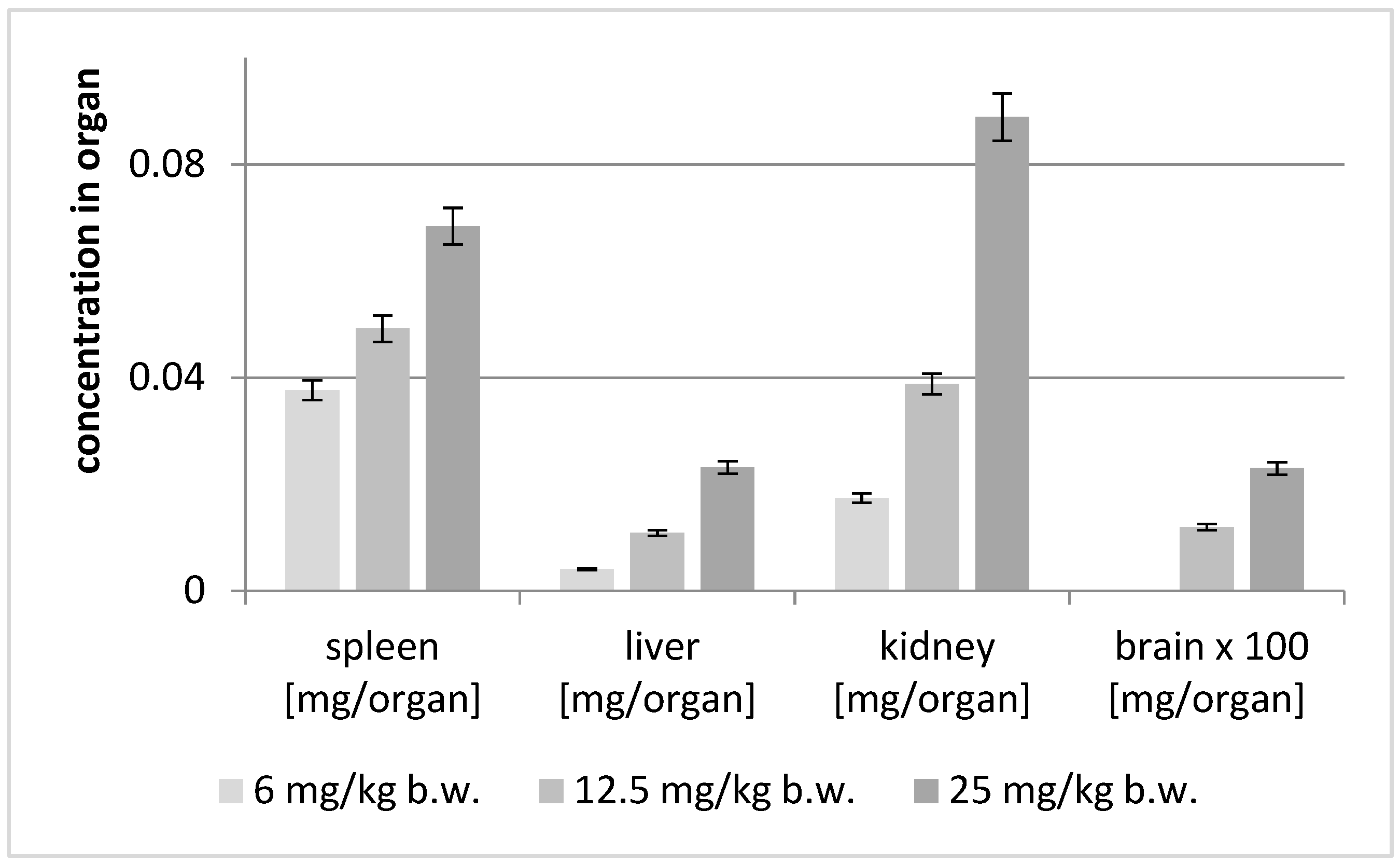
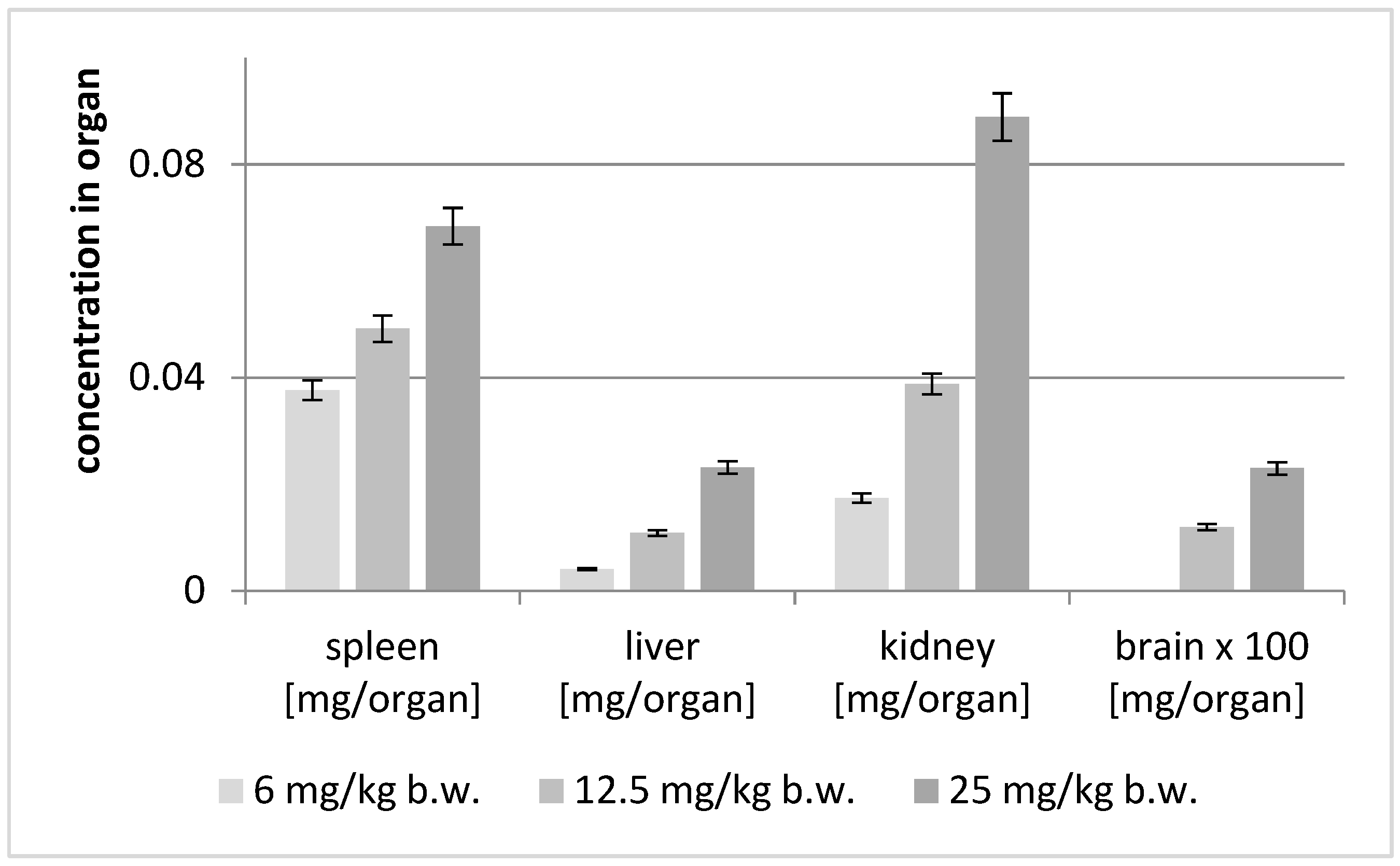
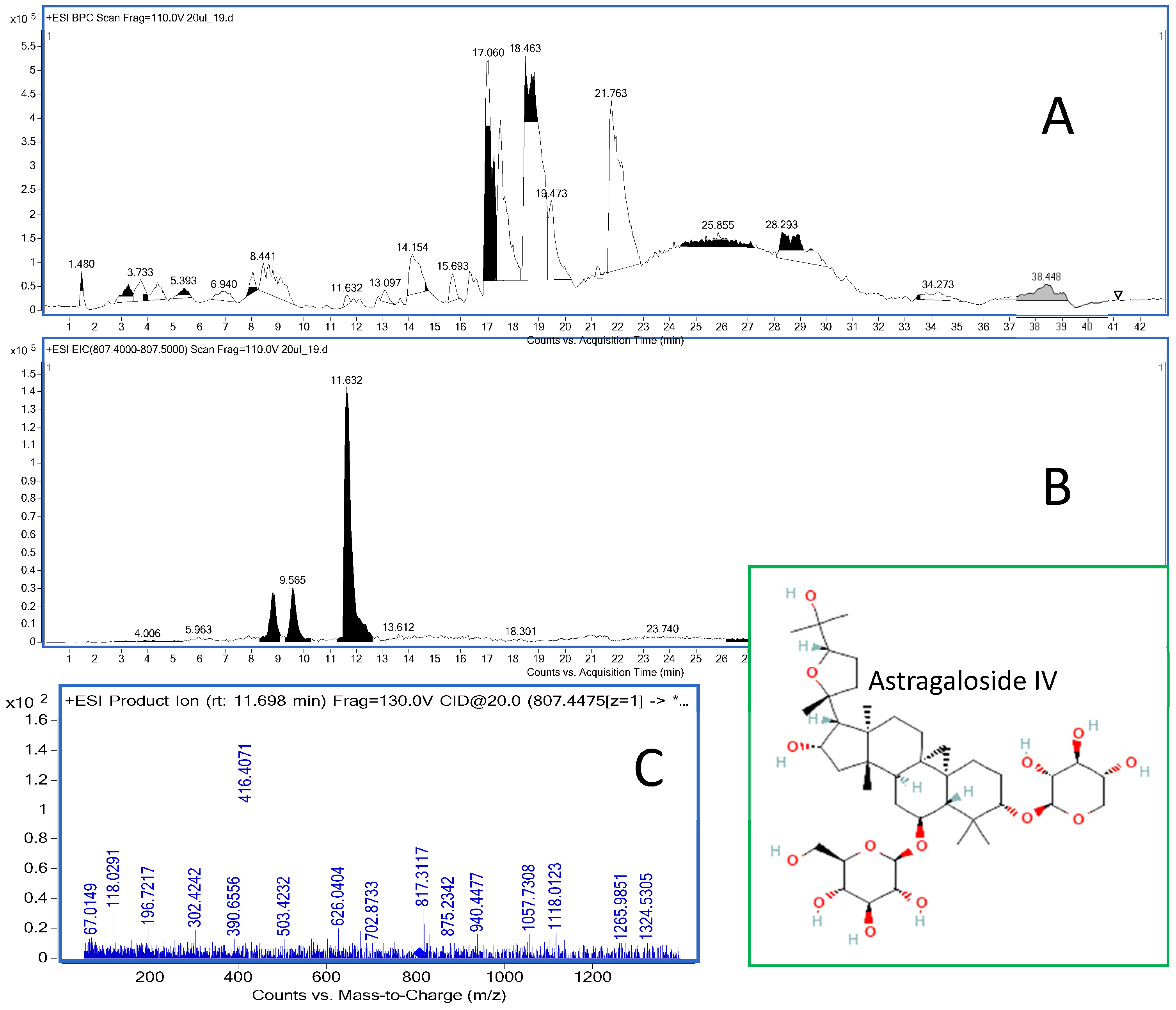
2.5. Post Mortem Analysis of the Distribution of Astragaloside IV in the Soft Organs
3. Discussion
4. Materials and Methods
4.1. Isolation of Astragaloside IV and ADME-Toxicity Profiling
4.2. Animal Studies
4.2.1. Mice
4.2.2. Medication
4.2.3. The Passive Avoidance Test
- Acquisition of long-term memory
- 2.
- Consolidation of long-term memory
4.2.4. The Locomotor Activity Test
4.2.5. Statistical Analysis
4.3. HPLC-ESI-QTOF-MS/MS Determination of Astragaloside IV in the Tissues of the Animals
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- World Health Organization. Global Action Plan on the Public Health Response to Dementia 2017–2025. 2017. Available online: https://www.who.int/publications/i/item/global-action-plan-on-the-public-health-response-to-dementia-2017---2025 (accessed on 10 April 2023).
- Hullinger, R.; Puglielli, L. Molecular and cellular aspects of age-related cognitive decline and Alzheimer’s disease. Behav. Brain Res. 2017, 322, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Kluever, V.; Fornasiero, E.F. Principles of brain aging: Status and challenges of modeling human molecular changes in mice. Ageing Res. Rev. 2021, 72, 101465. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.; Phan, C.W.; Lim, S.J.; Babji, A.S. Insights on the molecular mechanism of neuroprotection exerted by edible bird’s nest and its bioactive constituents. Food Sci. Hum. Wellness 2023, 12, 1008–1019. [Google Scholar] [CrossRef]
- Farooqui, A.A. Contribution of neuroinflammation, resolution, and neuroprotection in neurotraumatic diseases. In Neuroinflammation, Resolution, and Neuroprotection in the Brain; Farooqui, A.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 83–119. [Google Scholar]
- Rapposelli, S.; Digiacomo, M.; Balsamo, A. P-gp Transporter and its Role in Neurodegenerative Diseases. Curr. Top. Med. Chem. 2009, 9, 209–217. [Google Scholar] [CrossRef]
- Elufioye, T.O.; Berida, T.I.; Habtemariam, S. Plants-Derived Neuroprotective Agents: Cutting the Cycle of Cell Death through Multiple Mechanisms. Evid. Based Complement. Alternat. Med. 2017, 2017, 3574012. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. Alzheimer’s Disease: Genes, proteins, and therapy. Physiol. Rev. 2001, 81, 741–766. [Google Scholar] [CrossRef]
- Arriagada, P.V.; Marzloff, K.; Hyman, B.T. Distribution of Alzheimer-type pathologic changes in nondemented elderly individuals matches the pattern in Alzheimer’s disease. Neurology 1992, 42, 1681–1688. [Google Scholar] [CrossRef]
- Mrak, R.E.; Griffin, W.S.T. Interleukin-1, neuroinflammation, and Alzheimer’s disease. Neurobiol. Aging 2001, 22, 903–908. [Google Scholar] [CrossRef] [PubMed]
- Cole, G.; Teter, B.; Frautschy, S. Neuroprotective effects of curcumin. Adv. Exp. Med. Biol. 2007, 595, 197–212. [Google Scholar] [CrossRef]
- Garcia-Alloza, M.; Dodwell, S.A.; Borrelli, L.A.; Raju, S.; Bacskai, B. In vivo reduction of plaque size in APPswe/PS1D9 mice treated with curcumin. Alzheimer’s Dement. 2006, 2, S617. [Google Scholar] [CrossRef]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef] [PubMed]
- Houser, C.R.; Crawford, G.D.; Barber, R.P.; Salvaterra, P.M.; Vaughn, J.E. Organization and morphological characteristics of cholinergic neurons: An immunocytochemical study with a monoclonal antibody to choline acetyltransferase. Brain Res. 1983, 266, 97–119. [Google Scholar] [CrossRef] [PubMed]
- Woolf, N.J.; Butcher, L.L. Cholinergic systems mediate action from movement to higher consciousness. Behav. Brain Res. 2011, 221, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Woolf, N.J. Cholinergic systems in mammalian brain and spinal cord. Prog. Neurobiol. 1991, 37, 475–524. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Vieira, T.H.; Guimaraes, I.M.; Silva, F.R.; Ribeiro, F.M. Alzheimer’s disease: Targeting the Cholinergic System. Curr. Neuropharmacol. 2016, 14, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Pepeu, G.; Giovannini, M.G. The fate of the brain cholinergic neurons in neurodegenerative diseases. Brain Res. 2017, 1670, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Schliebs, R.; Arendt, T. The cholinergic system in aging and neuronal degeneration. Behav. Brain Res. 2011, 221, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Dunnett, S.B.; Everitt, B.J.; Robbins, T.W. The basal forebraincortical cholinergic system: Interpreting the functional consequences of excitotoxic lesions. Trends Neurosci. 1991, 14, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Hasselmo, M.E.; Anderson, B.P.; Bower, J.M. Cholinergic modulation of cortical associative memory function. J. Neurophysiol. 1992, 67, 1230–1246. [Google Scholar] [CrossRef]
- Lucas-Meunier, E.; Fossier, P.; Baux, G.; Amar, M. Cholinergic modulation of the cortical neuronal network. Pflugers Arch. 2003, 446, 17–29. [Google Scholar] [CrossRef]
- Sarter, M.; Bruno, J.P. Cognitive functions of cortical acetylcholine: Toward a unifying hypothesis. Brain Res. Rev. 1997, 23, 28–46. [Google Scholar] [CrossRef] [PubMed]
- Bucci, D.J.; Holland, P.C.; Gallagher, M. Removal of cholinergic input to rat posterior parietal cortex disrupts incremental processing of conditioned stimuli. J. Neurosci. 1998, 18, 8038–8046. [Google Scholar] [CrossRef] [PubMed]
- Voytko, M.L.; Olton, D.S.; Richardson, R.T.; Gorman, L.K.; Tobin, J.R.; Price, D.L. Basal forebrain lesions in monkeys disrupt attention but not learning and memory. J. Neurosci. 1994, 14, 167–186. [Google Scholar] [CrossRef] [PubMed]
- Pepeu, G.; Giovannini, M.G.; Bracco, L. Effect of cholinesterase inhibitors on attention. Chem. Biol. Interact. 2013, 203, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Bracco, L.; Bessi, V.; Padiglioni, S.; Marini, S.; Pepeu, G. Do cholinesterase inhibitors act primarily on attention deficit? A naturalistic study in Alzheimer’s disease patients. J. Alzheimers Dis. 2014, 40, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Jeon, S.J.; Jung, J.W.; Lee, S.; Yoon, B.H.; Shin, B.Y.; Son, K.H.; Cheong, J.H.; Kim, Y.S.; Kang, S.S.; et al. Tanshinone congeners improve memory impairments induced by scopolamine on passive avoidance tasks in mice. Eur. J. Pharmacol. 2007, 574, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Stępnik, K.; Kukula-Koch, W.; Plazinski, W.; Gawel, K.; Gaweł-Bęben, K.; Khurelbat, D.; Boguszewska-Czubara, A. Significance of Astragaloside IV from the Roots of Astragalus mongholicus as an Acetylcholinesterase Inhibitor-From the Computational and Biomimetic Analyses to the In Vitro and In Vivo Studies of Safety. Int. J. Mol. Sci. 2023, 24, 9152. [Google Scholar] [CrossRef] [PubMed]
- Stępnik, K.; Kukula-Koch, W. In Silico Studies on Triterpenoid Saponins Permeation through the Blood-Brain Barrier Combined with Postmortem Research on the Brain Tissues of Mice Affected by Astragaloside IV Administration. Int. J. Mol. Sci. 2020, 21, 2534. [Google Scholar] [CrossRef]
- van Breemen, R.B.; Li, Y. Caco-2 cell permeability assays to measure drug absorption. Expert Opin. Drug Metab. Toxicol. 2005, 1, 175–185. [Google Scholar] [CrossRef]
- DiMarco, R.L.; Hunt, D.R.; Dewi, R.E.; Heilshorn, S.C. Improvement of paracellular transport in the Caco-2 drug screening model using protein-engineered substrates. Biomaterials 2017, 129, 152–162. [Google Scholar] [CrossRef]
- Anastas, P.; Eghbali, N. Green chemistry: Principles and practice. Chem. Soc. Rev. 2010, 39, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Ekins, S.; Waller, C.L.; Swaan, P.W.; Cruciani, G.; Wrighton, S.A.; Wikel, J.H. Progress in predicting human ADME parameters in silico. J. Pharmacol. Toxicol. Methods 2000, 44, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Wilson, F.A.; Dietschy, J.M. The intestinal unstirred layer: Its surface area and effect on active transport kinetics. Biochim. Biophys. Acta 1974, 363, 112–126. [Google Scholar] [CrossRef] [PubMed]
- Wils, P.; Warnery, A.; Phung-Ba, V.; Legrain, S.; Scherman, D. High lipophilicity decreases drug transport across intestinal epithelial cells. J. Pharmacol. Exp. Ther. 1994, 269, 654–658. [Google Scholar] [PubMed]
- Artursson, P.; Palm, K.; Luthman, K. Caco-2 monolayers in experimental and theoretical predictions of drug transport. Adv. Drug Deliv. Rev. 1996, 22, 67–84. [Google Scholar] [CrossRef]
- New, R. Oral Delivery of Biologics via the Intestine. Pharmaceutics 2020, 13, 18. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Weickert, C.S.; Garner, B. Role of ATP-binding cassette transporters in brain lipid transport and neurological disease. J. Neurochem. 2008, 104, 1145–1166. [Google Scholar] [CrossRef] [PubMed]
- Tatsuta, T.; Naito, M.; Oh-hara, T.; Sugawara, I.; Tsuruo, T. Functional involvement of P-glycoprotein in blood-brain barrier. J. Biol. Chem. 1992, 267, 20383–20391. [Google Scholar] [CrossRef] [PubMed]
- Didziapetris, R.; Japertas, P.; Avdeef, A.; Petrauskas, A. Classification Analysis of P-Glycoprotein Substrate Specificity. J. Drug Target. 2003, 11, 391–406. [Google Scholar] [CrossRef]
- Lochner, M.; Thompson, A.J. The muscarinic antagonists scopolamine and atropine are competitive antagonists at 5-HT3 receptors. Neuropharmacology 2016, 108, 220–228. [Google Scholar] [CrossRef]
- Misane, I.; Ögren, S. Selective 5-HT1A Antagonists WAY 100635 and NAD-299 Attenuate the Impairment of Passive Avoidance Caused by Scopolamine in the Rat. Neuropsychopharmacology 2003, 28, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Araujo, D.M.; Lapchak, P.A.; Robitaille, Y.; Gauthier, S.; Quirion, R. Differential alteration of various cholinergic markers in cortical and subcortical regions of human brain in Alzheimer’s disease. J. Neurochem. 1988, 50, 1914–1923. [Google Scholar] [CrossRef] [PubMed]
- DeKosky, S.T.; Scheff, S.W.; Styren, S.D. Structural correlates of cognition in dementia: Quantification and assessment of synapse change. Neurodegeneration 1996, 5, 417–421. [Google Scholar] [CrossRef]
- Kuhl, D.E.; Koeppe, R.A.; Minoshima, S.; Snyder, S.E.; Ficaro, E.P.; Foster, N.L. In vivo mapping of cerebral acetylcholinesterase activity in aging and Alzheimer’s disease. Neurology 1999, 52, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Shinotoh, H.; Namba, H.; Fukushi, K.; Nagatsuka, S.; Tanaka, N.; Aotsuka, A. Progressive loss of cortical acetylcholinesterase activity in association with cognitive decline in Alzheimer’s disease: A positron emission tomography study. Ann. Neurol. 2000, 48, 194–200. [Google Scholar] [CrossRef]
- Bartus, R.T. On neurodegenerative diseases, models, and treatment strategies: Lessons learned and lessons forgotten a generation following the cholinergic hypothesis. Exp. Neurol. 2000, 163, 495–529. [Google Scholar] [CrossRef]
- Bartolomeo, A.C.; Morris, H.; Buccafusco, J.J.; Kille, N.; Rosenzweig-Lipson, S.; Husbands, M.G.; Sabb, A.L.; Abou-Gharbia, M.; Moyer, J.A.; Boast, C.A. The preclinical pharmacological profile of WAY-132983, a potent M1 preferring agonist. J. Pharmacol. Exp. Ther. 2000, 292, 584–596. [Google Scholar]
- Elrod, K.; Buccafusco, J.J. An evaluation of the mechanism of scopolamine-induced impairment in two passive avoidance protocols. Pharmacol. Biochem. Behav. 1988, 29, 15–21. [Google Scholar] [CrossRef]
- Blin, O.; Audebert, C.; Pitel, S.; Kaladjian, A.; Casse-Perrot, C.; Zaim, M.; Micallef, J.; Tisne-Versailles, J.; Sokoloff, P.; Chopin, P.; et al. Effects of dimethylaminoethanol pyroglutamate (DMAE p-Glu) against memory deficits induced by scopolamine: Evidence from preclinical and clinical studies. Psychopharmacol. Berl. 2009, 207, 201–212. [Google Scholar] [CrossRef]
- Blotnick-Rubin, E.; Anglister, L. Fine Localization of Acetylcholinesterase in the Synaptic Cleft of the Vertebrate Neuromuscular Junction. Front. Mol. Neurosci. 2018, 11, 123. [Google Scholar] [CrossRef]
- Gawel, K.; Labuz, K.; Gibula-Bruzda, E.; Jenda, M.; Marszalek-Grabska, M.; Silberring, J.; Kotlinska, J.H. Acquisition and reinstatement of ethanol-induced conditioned place preference in rats: Effects of the cholinesterase inhibitors donepezil and rivastigmine. J Psychopharmacol. 2016, 30, 676–687. [Google Scholar] [CrossRef]
- Gawel, K.; Gibula-Bruzda, E.; Dziedzic, M.; Jenda-Wojtanowska, M.; Marszalek-Grabska, M.; Silberring, J.; Kotlinska, J.H. Cholinergic activation affects the acute and chronic antinociceptive effects of morphine. Physiol. Behav. 2017, 169, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Batista, C.R.A.; Gomes, G.F.; Candelario-Jalil, E.; Fiebich, B.L.; de Oliveira, A.C.P. Lipopolysaccharide-Induced Neuroinflammation as a Bridge to Understand Neurodegeneration. Int. J. Mol. Sci. 2019, 20, 2293. [Google Scholar] [CrossRef] [PubMed]
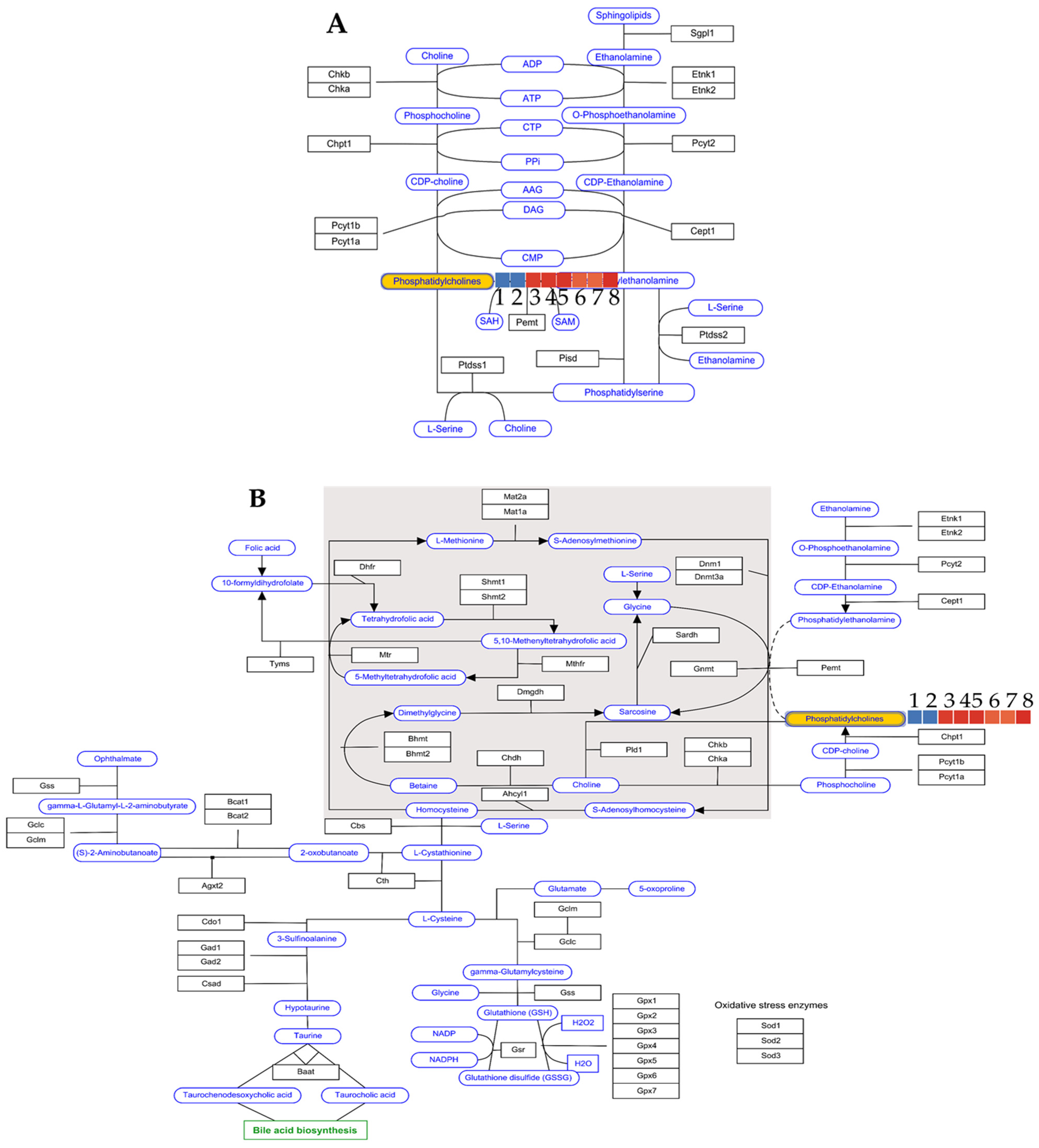
- Marcucci, H.; Paoletti, L.; Jackowski, S.; Banchio, C. Phosphatidylcholine biosynthesis during neuronal differentiation and its role in cell fate determination. J. Biol. Chem. 2010, 285, 25382–25393. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.M.; Williams, A.; Eisenbarth, S.C. Structure and function of the immune system in the spleen. Sci. Immunol. 2019, 4, eaau6085. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Liu, J.; He, Y.; Liu, L.; Xu, Z.; Lin, X.; Liu, N.; Kai, G. Systems pharmacology reveals the mechanism of Astragaloside IV in improving immune activity on cyclophosphamide-induced immunosuppressed mice. J. Ethnopharmacol. 2023, 313, 116533. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Jiang, H.; Tian, Y.; Zhao, W.; Wu, X. Astragaloside IV protects against polymicrobial sepsis through inhibiting inflammatory response and apoptosis of lymphocytes. J. Surg. Res. 2016, 200, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-D.; Zhang, C.; Liu, R.-H.; Li, H.-L.; Zhang, J.-T.; Mao, C.; Moran, S.; Chen, C.-L. Preclinical pharmacokinetics and tissue distribution of a natural cardioprotective agent astragaloside IV in rats and dogs. Life Sci. 2006, 79, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Feher, M.; Sourial, E.; Schmidt, J.M. A simple model for the prediction of blood-brain partitioning. Int. J. Pharm. 2000, 201, 239–247. [Google Scholar] [CrossRef]
- Liu, R.; Sun, H.; So, S.-S. Development of quantitative structure−property relationship models for early ADME evaluation in drug discovery. 2. Blood-brain barrier penetration. J Chem. Inf. Comput. Sci. 2001, 41, 1623–1632. [Google Scholar] [CrossRef]
- Van de Waterbeemd, H. In Silico Models to Predict Oral Absorption. In Comprehensive Medicinal Chemistry, II; Taylor, J.B., Triggle, D.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 669–697. [Google Scholar]
- Fischer, H.; Gottschlich, R.; Seelig, A. Blood-brain barrier permeation: Molecular parameters governing passive diffusion. J. Membr. Biol. 1998, 165, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Muehlbacher, M.; Spitzer, G.M.; Liedl, K.R.; Kornhuber, J. Qualitative prediction of blood-brain barrier permeability on a large and refined dataset. J. Comput. Aided Mol. Des. 2011, 25, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Hutter, M.C. Prediction of blood-brain barrier permeation using quantum chemically derived information. J. Comput. Aided Mol. Des. 2003, 17, 415–433. [Google Scholar] [CrossRef] [PubMed]
- Kotlinska, J.H.; Lopatynska-Mazurek, M.; Gawel, K.; Gabka, P.; Jenda-Wojtanowska, M.; Kruk-Slomka, M.; Marszalek-Grabska, M.; Danilczuk, Z.; Kedzierska, E.; Talarek, S.; et al. Impact of the metabotropic glutamate receptor7 (mGlu7) allosteric agonist, AMN082, on fear learning and memory and anxiety-like behavior. Eur. J. Pharmacol. 2019, 858, 172512. [Google Scholar] [CrossRef]
- Marszalek-Grabska, M.; Zakrocka, I.; Budzynska, B.; Marciniak, S.; Kaszubska, K.; Lemieszek, M.K.; Winiarczyk, S.; Kotlinska, J.H.; Rzeski, W.; Turski, W.A. Binge-like mephedrone treatment induces memory impairment concomitant with brain kynurenic acid reduction in mice. Toxicol. Appl. Pharmacol. 2022, 454, 116216. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | logPow | logBB | Pe,jejunum [10−4 cm/s] | Pe,Caco-2 [10−6 cm/s] | Number of H-donors | Number of H-acceptors | P-gp Substrate |
|---|---|---|---|---|---|---|---|
| AI | 5.020 | 0.46 | 1.05 | 2.0 | 7 | 16 | + |
| AII | 4.459 | 0.11 | 0.25 | 0.5 | 8 | 15 | + |
| AIII | 3.767 | 0.15 | 0.04 | 0.1 | 9 | 14 | - |
| AIV | 3.757 | 0.49 * | 0.06 | 0.1 | 9 | 14 | - |
| Brain [mg/Organ] | Spleen [mg/Organ] | Liver [mg/Organ] | Kidney [mg/Organ] | |||||
|---|---|---|---|---|---|---|---|---|
| Animal Group | Av | SD | Av | SD | Av | SD | Av | SD |
| 6 mg/kg | ND | ND | 0.03766 | 0.00186 | 0.00411 | 0.00011 | 0.01740 | 0.00209 |
| 12.5 mg/kg | 0.00012 | 0.00001 | 0.04919 | 0.00093 | 0.01084 | 0.00023 | 0.03882 | 0.00092 |
| 25 mg/kg | 0.00023 | 0.00002 | 0.06844 | 0.00133 | 0.02316 | 0.00016 | 0.08892 | 0.00613 |
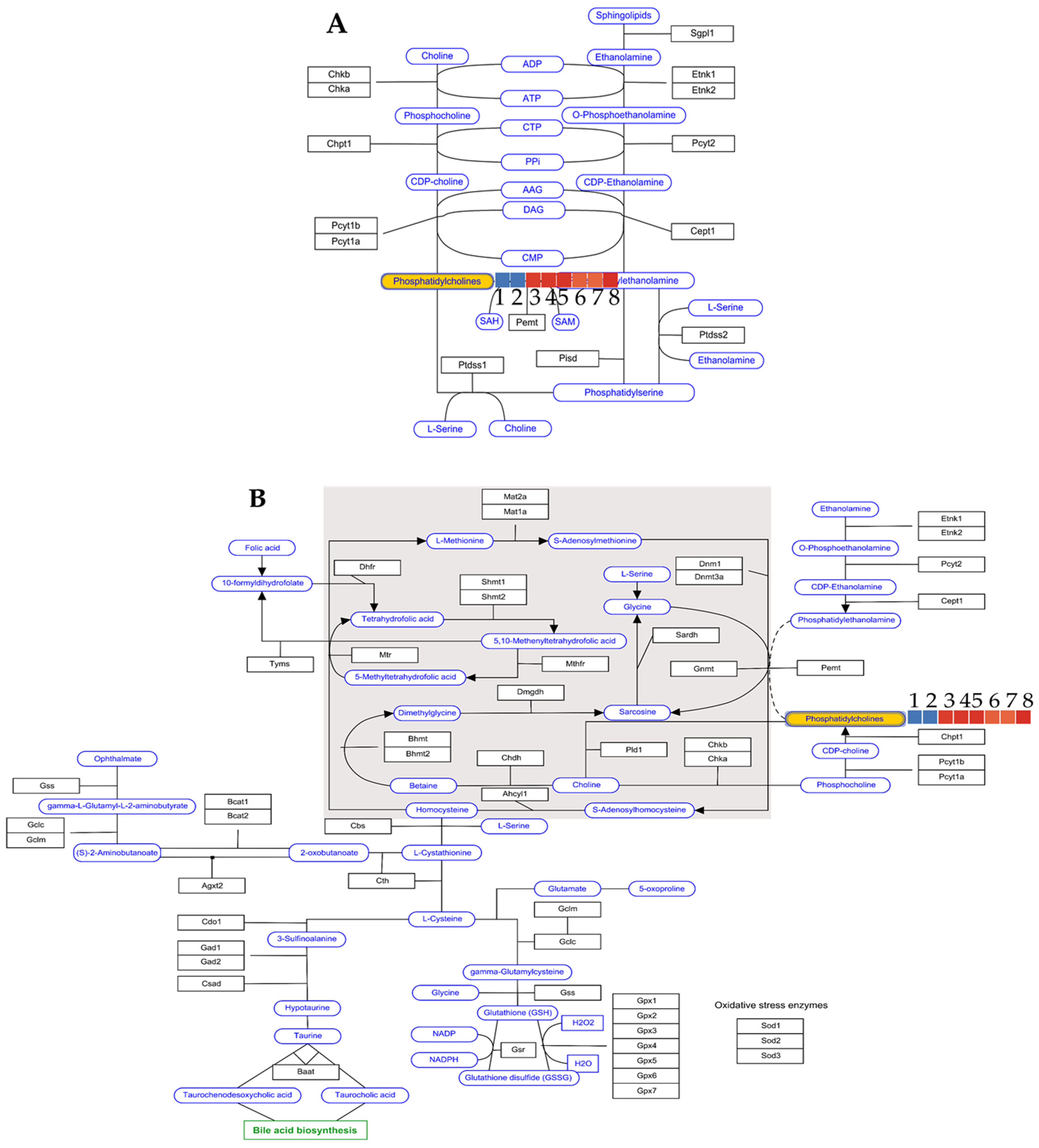
| Group Number | Group Type | Kennedy Pathway log2 | One-Carbon Metabolism log2 | Number of Brains Tested |
|---|---|---|---|---|
| 1 | LPS brain | 0 | 0 | 2 |
| 2 | LPS brain | 0 | 0 | 2 |
| 3 | Astragaloside IV 25 mg/kg | 20.425 | 20.425 | 3 |
| 4 | Astragaloside IV 25 mg/kg | 20.611 | 20.611 | 3 |
| 5 | Astragaloside IV 25 mg/kg | 20.763 | 20.763 | 3 |
| 6 | Saline group | 18.991 | 18.991 | 3 |
| 7 | Saline group | 18.766 | 18.766 | 3 |
| 8 | Saline group | 20.984 | 20.984 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stępnik, K.; Kukula-Koch, W.; Boguszewska-Czubara, A.; Gawel, K. Astragaloside IV as a Memory-Enhancing Agent: In Silico Studies with In Vivo Analysis and Post Mortem ADME-Tox Profiling in Mice. Int. J. Mol. Sci. 2024, 25, 4021. https://doi.org/10.3390/ijms25074021
Stępnik K, Kukula-Koch W, Boguszewska-Czubara A, Gawel K. Astragaloside IV as a Memory-Enhancing Agent: In Silico Studies with In Vivo Analysis and Post Mortem ADME-Tox Profiling in Mice. International Journal of Molecular Sciences. 2024; 25(7):4021. https://doi.org/10.3390/ijms25074021
Chicago/Turabian StyleStępnik, Katarzyna, Wirginia Kukula-Koch, Anna Boguszewska-Czubara, and Kinga Gawel. 2024. "Astragaloside IV as a Memory-Enhancing Agent: In Silico Studies with In Vivo Analysis and Post Mortem ADME-Tox Profiling in Mice" International Journal of Molecular Sciences 25, no. 7: 4021. https://doi.org/10.3390/ijms25074021
APA StyleStępnik, K., Kukula-Koch, W., Boguszewska-Czubara, A., & Gawel, K. (2024). Astragaloside IV as a Memory-Enhancing Agent: In Silico Studies with In Vivo Analysis and Post Mortem ADME-Tox Profiling in Mice. International Journal of Molecular Sciences, 25(7), 4021. https://doi.org/10.3390/ijms25074021