
Neutrophils and Anesthetic Drugs: Implications in Onco-Anesthesia

1
Department of Anesthesia and Intensive Care I, “Iuliu Haţieganu” University of Medicine and Pharmacy, 400012 Cluj-Napoca, Romania
2
Association for Research in Anesthesia and Intensive Care (ACATI), 400162 Cluj-Napoca, Romania
3
Onco-Anaesthesia Research Group, ESAIC, 1000 Brussels, Belgium
4
Outcome Research Consortium, Cleveland, OH 44195, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2024, 25(7), 4033; https://doi.org/10.3390/ijms25074033
Submission received: 12 February 2024
/
Revised: 24 March 2024
/
Accepted: 3 April 2024
/
Published: 4 April 2024
(This article belongs to the Special Issue The Role and Characterization of Neutrophils in Human Diseases)
{kind=link}
{kind=link}
Abstract
:Apart from being a significant line of defense in the host defense system, neutrophils have many immunological functions. Although there are not many publications that accurately present the functions of neutrophils in relation to oncological pathology, their activity and implications have been studied a lot recently. This review aims to extensively describe neutrophils functions’; their clinical implications, especially in tumor pathology; the value of clinical markers related to neutrophils; and the implications of neutrophils in onco-anesthesia. This review also aims to describe current evidence on the influence of anesthetic drugs on neutrophils’ functions and their potential influence on perioperative outcomes.
1. Introduction
Neutrophils (also known as polymorphonuclear leukocytes—PMNs) represent 50–70% of circulating leukocytes [1,2]. They are produced in bone marrow from where they enter the circulation and, subsequently, migrate to the tissues where they complete their functions [3]. PMNs can rapidly move to the tissues at the site of infection and inflammation via the leukocyte adhesion cascade, a complex cascade of adhesive steps between leukocytes and the endothelium [4,5].
Neutrophils are the first line of defense of the innate immune system against microorganisms [2,6], and this is accomplished by carrying out multiple antimicrobial functions [3,7,8].
The general functions of neutrophils include phagocytosis, degranulation, the production of reactive oxygen species (ROS) and the formation of neutrophil extracellular traps (NETs) [2,3,9,10,11,12]. More recently, numerous studies have reported additional specialized functions of neutrophils, namely producing cytokines and other inflammatory markers that contribute to the regulation of immune system functions [2], mediation/resolution of inflammation, and cytokine/chemokine modulation of immunity [13].
In the last few years, it has been shown that anesthetic agents may influence both the number and the subpopulations of neutrophils and their functions, so it is important to briefly describe these functions before describing the implications of anesthetic agents on neutrophils’ functions, leading to potential impacts on a patient’s outcome.
2. Neutrophils’ Functions
2.1. Phagocytosis
Phagocytosis is the process by which a particle is internalized by neutrophils into a phagosome where most of the pathogens are killed [2,14,15]. This process is receptor-mediated, during which pathogen-associated molecular patterns (PAMPs) are implicated in the recognition of pathogens [2,14] due to specific receptors for PAMPs on neutrophils’ surface [2,15].
2.2. Degranulation
Degranulation is the process during which effector molecules stored in internal granules of neutrophils destroy bacteria/pathogens [2]. These internal granules are classified as primary, secondary, and tertiary and contain myeloperoxidase (MPO), azuracidin, and defensins (primary granules, the first to be produced during the neutrophil development–promyelocyte phase), metalloproteinase (MMP)-9, lactoferrin (included in the secondary granules), albumin, and cytokines [16], and they are highly cytotoxic (as secretory granules in PMN) [2].
Circulating neutrophils are, normally, in a resting state when they cannot degranulate, leading to tissue damage. Neutrophils’ degranulation requires priming with the help of bacterial pathogens or products, pro-inflammatory cytokines, and chemokines, leading to neutrophils being ready to degranulate when in contact with different stimuli like pathogen-associated molecular patterns (PAMPs) and damage associate patterns (DAMPs) [2,13,17,18].
2.3. ROS
The production of reactive oxygen species (ROS) by nicotinamide adenine dinucleotide phosphate oxidase (NADPH-oxidase) in neutrophils plays an important role in the antimicrobial activity of these cells against bacterial and fungal infections and in inflammation [19]; they also prevent the excessive degranulation of tertiary granules of neutrophils [2,20]. ROS deficiency leads to recurrent severe bacterial infections [19,21,22], while excessive ROS formation leads to excessive inflammation and SIRS (e.g., during COVID-19 [23]). ROS have a direct antimicrobial activity, killing bacteria and fungi and some types of viruses [24], but they can also cross the bacterial cell membrane, damaging nucleic acids, cell membrane and cell proteins [19,25].
Once produced, superoxide anions (O2−) undergo dismutation to hydrogen peroxide (H2O2) that, subsequently, oxidizes ferrous iron and generates highly reactive hydroxyl radical OH−. On the other hand, MPO released from PMN granules converts hydrogen peroxide into a hypochlorous acid (HOCl) that is highly bactericidal [19,26]. ROS have the capacity to decrease bacterial growth during infections due to their ability to diffuse through bacterial membranes and may also damage their DNA, protein, and lipid molecules; ROS may be also involved in inflammation due to infection [26].
It is interesting to note that bacteria have developed intrinsic and extrinsic resistance mechanisms to ROS by developing scavenger mechanisms [19,26] and through iron sequestration and DNA damage repair [19,26].
It is also of interest that some anesthetic drugs, like lidocaine, may interfere with ROS production or their tissue effects [27]. Similarly, vitamin C and vitamin E act as ROS eliminating substances.
2.4. Neutrophil Extracellular Traps (NETs)
2.4.1. Formation of NETs
NETs are web-like networks of chromatin decondensed fibers covered with histones, enzymes from the granules (e.g., myeloperoxidase (MPO) and neutrophil elastase (NE) that can be used, along with citrullinated histone 3 (CitH3), as indirect markers of NETosis) [28,29] and antimicrobial proteins formed after nuclear membrane rupture, followed by the release of nucleoplasm into the cell and, finally, cell membrane destruction [3,30]. This network has a role in bacterial and cancer cell trapping and, finally, killing these cells after their removal from circulation [30].
The process of NET formation—NETosis—is defined as ROS-dependent regulated cell death [31]. However, progress has been made in describing the NETosis process, so it is actually considered to be a process that may be NADPH-oxidase (NOX)-dependent (former suicidal NETosis) and NOX-independent (so-called vital NETosis) [30,32,33], subsequently divided into three types [30]. The NOX-dependent pathway is triggered by lipopolysaccharide (LPS), phorbol 12-myristate 13-acetate (PMA), and bacteria, resulting in nuclear breakdown via ROS generation and the activation of certain enzymes like ERK1, P38, and Src [30,33]. On the other hand, the NOX-independent pathway is activated by bacteria and cancer cells, leading to the formation of ROS that act on certain enzymes, leading, in the end, to chromatin decondensation [30,34].
2.4.2. Assessment of NET Formation
Several methods are described for the visualization and quantification of NETs. They are classified as direct and indirect methods.
Immunofluorescence microscopy is a direct qualitative method consisting of granular staining for neutrophil-associated enzymes, histones, and extracellular DNA, using specific dyes and immunostaining for the enzymatic components of NETs [35,36]. This method allows NET identification in cell cultures, tissue sections, peripheral blood, or bronchoalveolar lavage. It can differentiate between the different stages of NET formation and between NET apoptosis and necrosis [35]. However, it is operator-dependent, time-consuming, laborious, and does not allow for quantitative analysis [36]. Nowadays, a semi-automatic quantitative assessment of NETosis is possible by using specific antibodies against histones, chromatin, and the cleavage site of the histone H3 [29,35].
Electron microscopy and scanning electron microscopy (SEM). Electron microscopy and SEM consist of an electron beam passing through or reflecting from the surface of the sample [35], which is then projected onto a fluorescent surface is collected by a sensor that creates an image [37]. SEM is first choice for visualizing NET formation. It creates a better image compared with simple electron microscopy [29]. The disadvantage is determined by the possibility of the electron beam stimulating the formation of NETs [29].
In vivo and in situ methods use intravital two-photon microscopy and subsequent electron emission detection based on DNA components [35,36]. Recent data show that NETs are less expressed in vivo compared to in vitro [29].
Real-time NETosis assessment uses the IncuCyte ZOOM imaging platform (Wilmington, DE, USA). This platform uses cell membrane properties with the permeability of DNA dyes from NUCLEAR-ID (Enzo Life Sciences, Inc., Farmingdale, NY, USA) and Sytox Green (Thermo Fisher cat no S7020, Waltham, MA, USA). By detecting morphological changes in cells and nuclei, this method can identify NETs from other cell deaths but has the disadvantage of generating false-positive results if the cell membrane integrity is compromised [38].
Flow cytometry and multispectral image flow cytometry allows the measurement of scattered light or fluorescence signals emitted by properly irradiated cells, leading to the differentiation of distinct cell populations in whole blood. It uses specific dyes (Sytox Green) or conjugated antibodies, which bind to cytokines, receptors, cellular components, and DNA [35]. Multispectral image flow cytometry can measure in whole blood both “suicidal” and “vital” NETs, and it can quantify NETs in the early stages that underestimate the formation of NETosis at a longer time interval [29,35].
Enzyme-linked immunosorbent assay (ELISA) quantifies some products of neutrophil degradation characteristic of NETs without direct visualization [29,35]. However, there is controversy regarding the best marker for detecting NET formation [35]. The CitH3-DNA ELISA is validated for reliable NET quantification in human plasma. While MPO-DNA complexes may also be identified via ELISA, discrepancies between in vivo and in vitro values have been identified [35].
Western blot is another indirect method, using specific antibodies to quantify a protein involved in the formation of NETs. It is a very sensitive and specific method with some disadvantages including costs and the looseness of specificity in a mix of protein antibodies [35].
2.4.3. Clinical Implications of NETs
NETs in Critical Patients
Although, as shown before, NETs contribute to bacterial clearance, excessive NETs promote inflammation and tissue injury in sepsis and other inflammatory diseases [39]. Thus, high levels of NET formation have been described for the bacterial origin (pneumonia) of ARDS or ventilation pneumonia [40], while there is a correlation between the severity of ARDS and NET serum level [41], as well as between the NET (evaluated by MPO-DNS complex) level and the severity of organ dysfunction and 28-day mortality in sepsis and septic shock [39,40]. Higher levels of NETs have been described during COVID-19, where NETs act as triggers of a cascade of inflammatory reactions, leading to a cytokine storm that is a characteristic of this viral infection [42,43]. NETs are also increased in cardiovascular diseases especially in atherosclerosis, venous thrombosis, and inflammatory pathologies like pancreatitis [44].
NETs and Cancer and Cancer Associated Thrombosis
Tumor progression depends on the balance between the inflammatory response generated by tumor-infiltrating immune cells and tumor-derived factors in the tumor microenvironment (TME). Neutrophils are the first cells to reach the inflammation/injury site. Recent data show that inflammation is closely linked with cancer development and that NETs generated by neutrophils in TME play a critical role in tumor development, progression, metastasis, and resistance to therapy [45,46,47].
This is why, more recently, NETs (or their indirect markers) have been considered biomarkers for different types of cancers [46]; elevated serum levels of NETs may be associated with cancer diagnosis [30,32,33]. It was demonstrated that high levels of CitH3 are associated with a poor prognosis in cancer patients [46,48,49]. The pathophysiology of NET formation in TME has a starting point in tumor cell necrosis generated by the hypoxic TME repleted with cytokines. DAMPs are, thus, released, causing inflammation and triggering NET formation, along with the activation of ROS pathways [30,45,46,50]. Studies have also showed that NETs are key players in transformation from inflammation to carcinogenesis by awakening dormant tumor cells and that the accumulation of intertumoral NETs facilitates tumor growth [46,50].
NETs are also implicated in tumor metastasis and cancer-associated thrombosis. It has been shown that NET-bound cancer cells, thus, promote metastasis [3,44,51]. It has also been shown that in certain types of cancers (breast, lung), metastases are associated with high levels of NETs [44,52,53,54]. Other implicated mechanisms are NET-DNA receptors on the tumor cell membrane [54]; cathepsin C, a protease released by tumor cells that primes NET formation, and the release of G-CSF at sites of NET dissemination and interleukin disbalance [44,54,55].
3. Cytokine Release
In the last few years, it has been shown that neutrophils’ release stimulates pro- (IL-6, TNF-α, IL-1, IL-17, IFN-α and γ) and anti-inflammatory interleukins (IL-4, IL-10, etc.), as well as other chemokines (CXCL1, CXCL8, CXCL10, CCL2-4), angiogenic factors, and growth factors [9,56,57]. This is another argument for the involvement of neutrophils in different clinical conditions, like acute inflammatory diseases, wound healing, autoimmune diseases, and cancer [9,56,57]. Neutrophils produce cytokines via degranulation, protein synthesis, NET-associated cytokine release, and the expression of receptor-bound cytokines [58].
4. Neutrophil-to-Lymphocyte Ratio (NLR)
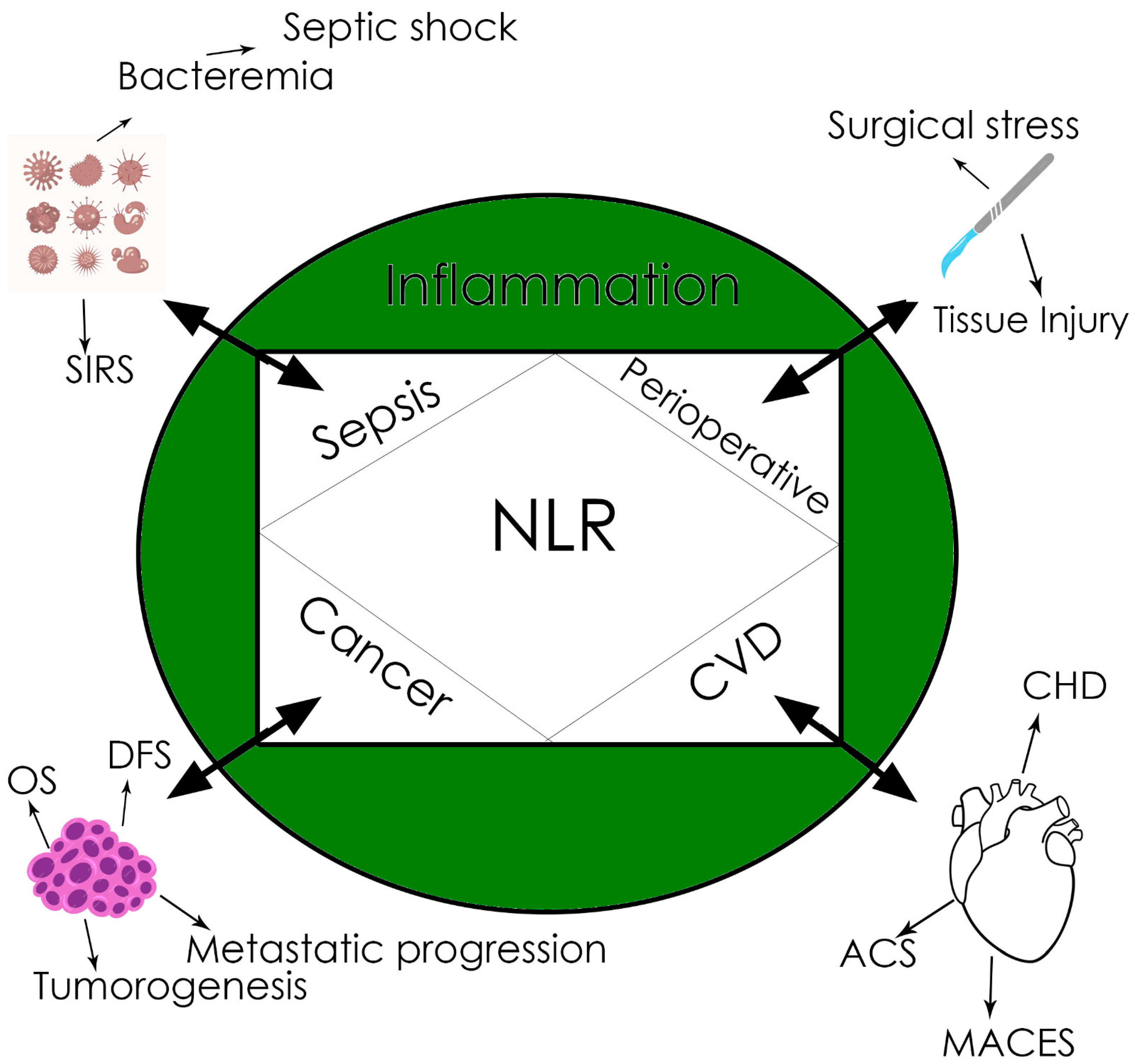
The NLR (ratio between the neutrophil and lymphocyte counts in peripheral blood) is a newly described biomarker, linking innate (the first-line neutrophils) and adaptive immunity (lymphocytes), through numerous relationships, with different diseases like sepsis, cancer, myocardial infarction, etc. [59,60,61], as well as with basically with any condition associated with tissue damage and inflammation that is ultimately followed by SIRS [59]. Even if a cut-off value is not definitely established and different values have been reported to be of reference (Figure 1), the significance of the NLR is high in many clinical conditions; most of the studies agree that an NLR value above three significantly increases mortality and worsens prognosis [59,61,62].
Of particular interest for anesthesia is the value of the NLR as a biomarker in sepsis and cancer and during the perioperative period. It has been demonstrated that the NLR is an early biomarker for sepsis and prognosis in sepsis, especially in those patients without leukocytosis [59,62,63].
In cancer biology, inflammation plays an important role in tumorigenesis, while in turn, cancer, once initiated, maintains inflammation due to tumor-released pro-inflammatory cytokines, subsequently recruiting immune cells [59,64]. This is why the NLR has been proved to be an early and reliable biomarker for cancer-related inflammation and prognosis, especially in solid tumors like hepatocarcinoma or colorectal cancer [59,65,66]. Moreover, it has been shown that the NLR may predict OS and DFS in surgical cancer patients.
In surgery, the preoperative value of the NLR was proved to be correlated with postoperative complications and mortality for different types of surgical interventions [59,67,68].
The Role of Neutrophils in Cancer
Despite many years of research, it is only in the last few years that the role of neutrophils in cancer has been extensively studied and begun to be understood.
It was shown in the previous sections that inflammation plays an important role in tumorigenesis [69]. Inflammation, especially chronic inflammation, acts as a promoter for transforming subthreshold neoplastic states into tumors [69]. Once developed, the tumor produces cytokines and chemokines that attract different leukocyte populations: neutrophils, macrophages, lymphocytes, and mast cells. These cells produce pro-inflammatory cytokines (of particular interest are TNF-α, IL-6, and interferon), proteases, MMPs, and NETs that support inflammation and favor tumor development and metastasis. Monocytes attracted to the tumor site turn into tumor-associated macrophages (TAMs) that play the role of a two-sided coin: on one side, they kill tumor cells, and on the other side, they produce a number of angiogenic (VEGF-C, D, F) and lymphangiogenic growth factors, cytokines, and proteases involved in tumor biology and progression [69,70].
The same double role in tumorigenesis is held by neutrophils. There is growing evidence that neutrophils play an important role in cancer biology and tumor progression [59,64]. It has been reported that large numbers of tumor-associated neutrophils (TANs), as well as an increased NLR, are associated with advanced stages of cancer, incomplete surgical resection, and poor short- and long-term postoperative prognosis [59,65,66,71]. The pro-tumor functions of neutrophils include ROS, MMPs, cytokines, and chemokines [71]. MMPs (especially MMP9) stimulate tumor cell proliferation and the production of VEGF and subsequent angiogenesis, while IL-6, IL-1β, and TNF-α, among pro-inflammatory cytokines, support chronic inflammation preceding or associated with tumor [72]. ROS are involved in the genotoxicity in tumor cells promoting carcinogenesis and neutrophil elastases promoting tumor cell proliferation [72,73]. Neutrophils also stimulate metastasis via a number of mechanisms including NETosis, promoting the retention of circulating tumor cells via ICAM clustering or direct adhesion [72]. Another mechanism implicated in the pro-tumor effects of neutrophils is represented by gMDSCs (also called PMN-MDSCs). These are low-density neutrophils with a suppressive and pro-tumoral phenotype that become detectable in advanced stages of cancer and are associated with T-cell impairment and immune suppression [74]. These cells play a role in tumor progression by promoting tumor vascularization, neoplastic cell local invasion, and distant metastasis and via the regulation of T and NK cells’ anti-tumor activity. In practice, MDSCs form a pre-metastatic niche, allowing the invasion of disseminated cancer cells [74,75]. These cells are targets for therapeutic modulation in cancer patients.
The other side of the coin represents the anti-tumor effects of neutrophils. There is clear evidence that neutrophils have a number of anti-tumor effects. Thus, neutrophils may directly kill cancer cells when activated by different mechanisms including ROS (inducing tumor cell lysis in early stages or rapidly growing tumors), MMPs (MMP8), direct lysis and apoptosis, and the regulation of T-cell functions [67,76].
The role of inflammation in cancer development and progression is also clinically supported by the anti-tumor effects of NSAIDs and aspirin that will be discussed later in this review.
5. The Influence of Anesthetic Drugs on Neutrophils and Their Functions
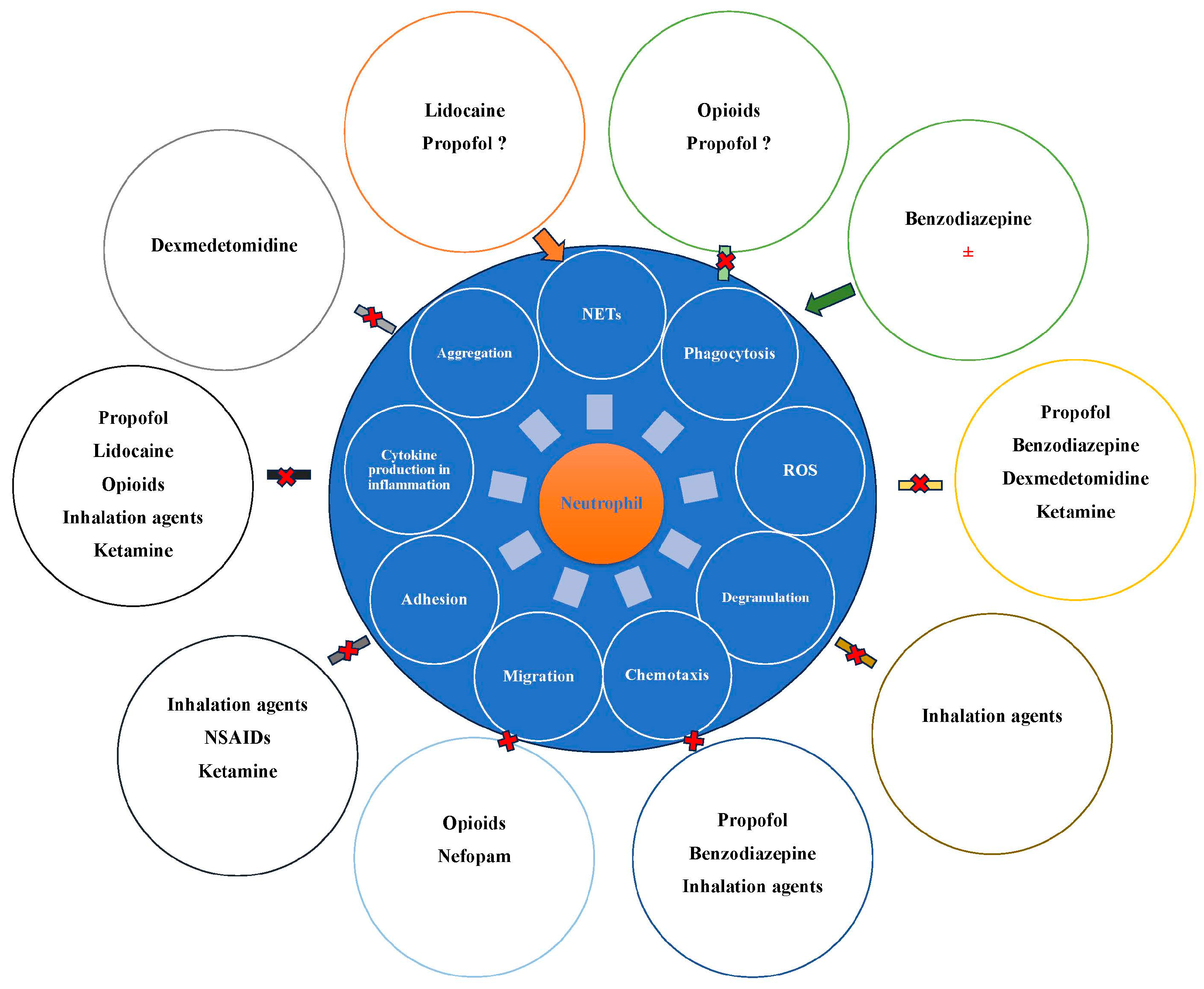
Over the years, a number of studies have reported that anesthetic agents may influence neutrophils and their functions (Figure 2).
5.1. Intravenous Anesthetic Agents and Neutrophils
5.1.1. Propofol
Propofol (2,6-diisopropylphenol) is, probably, the most widely used i.v. agent for both the induction and maintenance of anesthesia (total intravenous anesthesia—TIVA). TIVA has been extensively used in clinical practice in the last few years for different reasons: environmental protection, quality of recovery, risk of malignant hyperthermia, potential decreased risk of delirium, and surgical reasons (interference of anesthesia with the surgical field). In the last few years, after Wigmore’s study was published [77], a number of publications also reported that TIVA is followed by increased overall survival and disease-free survival after surgery in certain types of cancers, especially digestive cancers [78,79,80].
Propofol acts on GABA A receptors, allowing Cl− ions to enter the cells [81,82]. GABA is one of the most important neurotransmitters in the central nervous systems [83], and GABA A (and B) receptors are widely distributed in neutrophils, monocytes, and macrophages, probably mediating the effect of propofol in immune cells [84]. In higher concentrations, propofol directly opens chloride channels and blocks glutamate release via the inhibition of certain sodium channels [81,85]. Propofol also inhibits the N-methyl-d-aspartate (NMDA) receptors and modulates calcium influx, while muscarinic and dopaminergic receptors have also been involved in propofol’s mechanisms of action [81,86].
In clinically relevant concentrations, older studies reported that propofol inhibited phagocytosis and the killing of E.coli and Staphylococcus aureus [87], while others did not find this effect [88]. More recent publications highlighted propofol’s effects such as regulating macrophage and neutrophil phagocytosis, pyroptosis, the production of pro-inflammatory cytokines (especially IL-6), and the inhibition of macrophage migration and recruitment [83,89]. Taking into consideration the effect of propofol on bacterial killing, we may expect an increase in the incidence of infections in ICUs where propofol is used for sedating critical patients for long periods of time. Even if clinical studies are limited, however, the studies published so far have not reported an increase in the incidence of pneumonia or surgical site infections in patients with TIVA compared with inhalation anesthesia, but on the contrary, the incidence was lower in abdominal surgery cases [90,91]. The same lack of significant differences was reported between sedation with propofol and dexmedetomidine in ICUs [92]. What is interesting is the fact that these effects are not mediated in humans by GABA A receptors, which are not elicited on neutrophils’ surfaces but via other mechanisms, like the inhibition of ROS production, p44/42 mitogen-activated protein kinase phosphorylation, and mitochondrial membrane potential [81,83,93,94]. Since most of these studies have been performed in animals, there is a lack of human studies where confounding factors may interfere, especially studies in critically ill patients, so prospective RCTs in critically ill patients for the potential effects of propofol sedation are mandatory.
5.1.2. Ketamine
Ketamine is an i.v. anesthetic agent increasingly used in the last few years mainly due to it having analgesic and antidepressant properties without depressing respiration. Ketamine may also be used as an induction agent (alone or in combination with other agents or opioids) in unstable patients, taking into consideration its hemodynamic effects, especially for increasing blood pressure. Ketamine mainly acts as an antagonist of NMDA receptors, but other targets for its action have been described: the GABA A receptor, cyclic nucleotide-gated subtype 1 receptor (hyperpolarization-activated), and α- and β-adrenergic, -muscarinic, and -opioid receptors [81,95,96,97].The few studies of ketamine’s effects on neutrophils showed that ketamine attenuated the superoxide anion production of human neutrophils via PMA (phorbol myristate acetate) stimulation and decreased oxidative burst (via decreased phosphorylation of p47) and neutrophil adhesion in a concentration-dependent manner [81,98]. However, further studies are necessary in the context of the increased use of ketamine as an adjunct to anesthesia and as an antidepressant.
5.1.3. Benzodiazepines
Benzodiazepines act by potentiating the inhibitory effect of GABA at the postsynaptic site, resulting in the inhibition of neuronal membrane via hyperpolarization. Some benzodiazepines act on GABA A peripheral benzodiazepine receptors. Studies of the effect of benzodiazepines in human neutrophils are limited and show contradictory effects. Most of the studies showed the inhibition of fMLP-induced human neutrophil chemotaxis and superoxide production (for diazepam) [81], reduced oxidative burst, and reduced CD11b/CD18 expression and p38 mitogen-activated protein kinase phosphorylation in neutrophils (for midazolam) [81,99]. However, Marino et al. reported increased human neutrophil migration and phagocytosis via an interaction with the peripheral benzodiazepines’ receptors for diazepam [100]. Diazepam appeared to enhance phagocytosis by increasing intracellular calcium, while the stimulatory effect on migration was not calcium-dependent [100].
5.1.4. Alpha-2 Agonists
α2-adrenergic receptor agonists stimulate α2 adrenergic receptors, and in the case of clonidine, central imidazoline receptors are added as targets. More recently, dexmedetomidine has been commonly used for the sedation of critically ill patients in ICUs (in which case, it is administered for up to 72 days, or for even more days in some cases) or as an adjunct to anesthesia. Both dexmedetomidine and clonidine have analgesic effects and decrease opioid use. The treatment of delirium or alcohol withdrawal are other indications. Neutrophils express adrenoreceptors (including α2) on their surfaces, so an interaction with α2 agonists is expected at least due to this target [101]. A number of mostly in vitro studies in the last few years have shown that dexmedetomidine suppressed pro-inflammatory cytokines (IL-6, TNF-α, Necrosis factor) and antimicrobial effectors (ROS, RNS, NO, iNOS), as well as the local aggregation of neutrophils, leading to an in vitro anti-inflammatory effect that may be beneficial in critically ill patients in states with increased inflammatory response (SIRS) [102]. A number of pre- and mid-clinical studies suggest that dexmedetomidine may have anti-tumor effects by having indirect anti-inflammatory effects and directly inhibiting the survival, proliferation, and invasion of tumor cells [103].
Further clinical studies are necessary to quantify the impacts of these effects on long-term postoperative outcomes in cancer patients.
5.1.5. Inhalation Agents and Neutrophils
Inhalation agents act via numerous mechanisms, including increasing the sensitivity of GABA A receptors [81,104], potentiating the effects of glycine (an inhibitory transmitter) on glycine receptors, inhibiting α4β2 nicotinic acetylcholine receptors, and interacting with ion channels including the family of neurotransmitter receptors [105,106]. Volatile anesthetics impaired neutrophil functions by decreasing the number of reacting PMNs, chemotaxis and adhesion, and ROS production [107,108,109]. However, there are studies reporting contrary results, i.e., increased phagocytosis of apoptotic neutrophils by macrophages, thus reducing lung inflammatory injury after isoflurane exposure or having no impact on phagocytosis after isoflurane exposure [110,111]. A number of mostly animal model studies, subsequently focused on the effects of volatile agents on inflammation after ischemia/reperfusion. It has been reported that inhalation agents decreased neutrophil infiltration and subsequent organ injury and TNF-α and IL-6 levels [107,108,109,112]. It was also shown that inhalation agents suppress NK cells’ activity, with potential implications for immune surveillance and immune suppression [113,114,115] after cancer surgery, but data on these effects’ impacts on clinical long-term outcomes are still missing.
5.1.6. Local Anesthetics (LAs)
LAs are used mostly in the context of regional anesthesia. However, in the last few years, the i.v. infusion of lidocaine has gained popularity due to its multiple effects. It has been demonstrated that regional anesthesia/analgesia reduces stress response to surgery and the need for opioids while increasing NK cells’ activity and the number of T-helper cells.
LAs act by blocking voltage-gated sodium channels (VGSCs), thus blocking the transmission of the neural signal [116]. VGSCs are a family composed of nine members (nine subtypes defined by the nine types of α subunit) and are common not only in nervous systems but also in other types of cells, like heart muscles, pancreatic cells, and cancer cells. This is why VGSCs and LAs have many clinical implications in cardiac and respiratory disorders, diabetes, cognitive dysfunctions, and cancer [117].
Recent studies have shown that lidocaine, in vitro and in humans (as well as in Kulińska’s study of ropivacaine), enhanced NETosis in neutrophils at clinically relevant concentrations, thus having the possibility to change the course of inflammation and metastasis [118,119]. Kulinska et al. also showed that LAs upregulate nitric oxide synthase (NOS) production by neutrophils, thus increasing NO synthesis [120]. Moreover, a recent meta-analysis showed that perioperative i.v. lidocaine can be recommended as an anti-inflammatory and analgesic for elective surgery [121]; the same effects were reported in emergency departments for different painful inflammatory conditions [122]. It was shown that after elective surgery, lidocaine decreased IL-6, TNFα, and IL-1RA levels [121]. Further studies are also needed to better define the roles of LAs and, in particular, i.v. lidocaine as anti-inflammatory and analgesic drugs.
5.1.7. Opioids
Opioids are used perioperatively and in painful conditions for analgesia. These drugs exert their effect by acting on G-coupled protein µ-, δ-, and κ-receptors. Opioid receptors are widely expressed in the central and peripheral nervous systems and non-neuronal cells, like the gastrointestinal tract or immune cells [123]. µ-opioid receptors and nociceptin–orphanin receptors have been described in polymorphonuclear cells and NK, T, and B cells [123]. These receptors can be activated not only by opioids but also by cytokines (IL-1, IL-2, IL-7, IFN, TNF) and other inflammatory mediators. This is the pharmacological rationale for which, besides the well-known side effects of opioids mediated via central opioid receptors (e.g., respiratory depression), immune suppression is another effect, mediated by peripheral opioid receptors. In the last few years, it has been shown that morphine and other opioids are associated with an increased risk of infection and sepsis [124]. Morphine and other opioids inhibit neutrophil functions including phagocytosis, migration, antibody, and interleukin/chemokine production, thus influencing immune response during infections or cancer [125]. In cancer, on the other hand, certain neutrophils release endogenous opioids that diminish pain and inflammation, thus having potential negative effects on tumor progression [126]. Moreover, there are differences between opioids in immune effects and effects on cancer progression, with morphine probably being the most immune-suppressing opioid, while fentanyl has some immune stimulatory effects on regulatory T cells [127]. Further studies are very much needed to identify if these differences between opioids have clinical significance in cancer patients.
5.2. Minor Analgesics and Neutrophils
5.2.1. NSAIDs/Aspirin
NSAIDs are used during the perioperative period for analgesia. Their main mechanism of action is the inhibition of cyclooxygenase 1 and 2 (COX), enzymes involved in prostaglandin synthesis; thus, NSAIDs inhibit prostaglandin synthesis. Other mechanisms of action include interference with cell adhesion, involving cancer cells, platelets and leukocytes adhesion; the generation of superoxide anion in the plasma membranes of leukocytes, and the inhibition of neutrophil elastase [128]. On the other hand, NSAIDs have antineoplastic and neuroprotective effects [129]. Several types of cancers overexpress COX with subsequent implications for tumorigenesis [127,130]. Some of the NSAIDs inhibit transcription factors like NF-kappa B, influence neutrophil chemotaxis, or regulate signaling pathways [128], more recently described as other mechanisms for the anti-inflammatory effects of NSAIDs [127,128].
5.2.2. Nefopam
This newly introduced centrally acting non-opioid analgesic mainly acts by inhibiting the synaptic uptake of several transmitters like dopamine, noradrenaline/norepinephrine, and serotonin. Recently, it has been shown that nefopam inhibits neutrophil migration and wound-induced neutrophil recruitment, thus having anti-inflammatory effects, even if it is not an anti-inflammatory drug initially [131].
5.2.3. Impacts on Short- and Long-Term Outcomes
Taking into consideration the effects of anesthetic agents not only on neutrophils but also on immune response and cancer cell biology, the question is whether these effects impact the short- and long-term outcomes of surgical patients. At the moment, there are not many definitive conclusions on the impact of anesthetic agents on patients’ outcomes, and most of the studies are only under way. However, regarding regional anesthesia and postoperative long-term outcomes in cancer patients, most of the studies did not find a significant impact of regional anesthesia on cancer mortality and the incidence of recurrence [132,133].
The other question is whether TIVA may ameliorate long-term outcomes in cancer patients compared with inhalation anesthesia. While most of the studies in breast cancer patients did not find any difference [133], for other types of cancer (e.g., digestive cancers), there may be a difference in outcome, and some of the RCTs under way (NCT02786329, NCT04188158, NCT00684229, NCT01975064, NCT06017141) may bring light on this issue [133,134]
Preclinical and clinical studies published in the last few years on i.v. lidocaine are encouraging regarding short-term outcomes (pain, inflammation) in cancer patients, taking into consideration its anti-inflammatory and anti-tumor effects, so another question is whether i.v. lidocaine may impact disease-free survival and OS in cancer patients undergoing surgery.
At the moment, there are no data on this topic, but the above-mentioned RCTs under way may also generate more data on this issue.
Last but not least, another intriguing the question is whether anesthetics may, on the other hand, promote the pro-tumorigenic activity of neutrophils. To our knowledge, at this moment, there are no data on this aspect, so further studies will have to focus not only on the anti-tumor effects of certain anesthetic agents/techniques but also on the effects of promoting the pro-tumorigenic effects of certain neutrophil subpopulations.
5.3. Anesthetic Drugs and the NLR
As shown before, the NLR is a newly described biomarker closely related to patient outcomes. The NLR has many implications for anesthesia in sepsis and cancer and predicting postoperative complications [59,67,68]. As a consequence, a number of studies aimed to investigate whether anesthetic drugs may influence the NLR. In a recent study, Hasselager reported that the NLR was significantly higher during propofol anesthesia and on day 1 compared with sevoflurane in healthy volunteers who did not undergo surgical intervention [79]. However, Ní Eochagáin reported that a propofol–paravertebral block slightly attenuated the increase in the NLR in breast cancer surgery, with no benefit for long-term outcomes [135].
I.v. lidocaine postoperatively decreased the NLR after breast cancer surgery [136], while Alexa et al. did not find an influence of i.v. lidocaine at 24 h after colorectal surgery [137]. An erector spinae plane block (ESPB) significantly decreased the NLR after spine surgery [138]. The impact of the influence of anesthetic agents on the NLR for patient outcomes still needs to be determined via future RCTs studying long-term outcomes. Such studies should follow patients for a long time postoperatively to assess NLR variations’ relationship with clinical outcomes.
5.4. Anesthetic Agents and the Efficacy of Anti-Cancer Drugs Administered after Surgery
There is a paucity of data on the potential interactions between anesthetic agents and anti-cancer drugs, namely chemotherapeutical agents and immune therapy. The interaction between anesthetic agents and anti-cancer drugs may be produced via an indirect mechanism of modulation of the host immune response and direct interference with cancer cell biology. The few mostly in vitro and animal model studies published so far showed that propofol may increase chemotherapeutic activity via the inhibition of Slug expression (increasing paclitaxel-induced apoptosis), abrogation of gemcitabine-induced DNA-binding activity of NF-kB, and downregulation of NF-kB [139,140]. However, there are studies showing that propofol may decrease chemotherapeutic efficiency via the inhibition of gap junction function [139,140,141].
Lidocaine may attenuate chemotherapeutic drug resistance or increase chemotherapeutic drug efficiency by decreasing the expression of miR-21 or via the downregulation of ABC transporter protein expression [139]. Animal model studies also showed that lidocaine decreased tumor proliferation and increased chemotherapeutic drug efficiency via the activation of caspase 3, decreased Bcl-2 and Bax expression, and the inactivation of ERK1/2 and p38 pathways [142]. Future studies are needed to determine the clinical significance of these results in terms of ameliorating long-term outcomes.
6. Conclusions
Neutrophils are key factors in defense against infection and cancer and, in consequence, are important in onco-anesthesia and the perioperative period. Although many factors intervene during this period, there are strong arguments that anesthetic agents may influence neutrophils and their functions. Whether this influence has a meaningful impact on the short- and long-term outcomes of surgical patients remains to be determined in future large RCTS, which are urgently needed. At the moment, there are insufficient data to justify choosing certain anesthetic techniques/drugs to ameliorate postoperative outcomes in cancer patients.
Author Contributions
D.I. conducted the literature research; D.I. drafted the manuscript; D.I., A.L.A. and S.S. critically revised the manuscript; A.L.A. and S.S. conceived the figures. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mestas, J.; Hughes, C.C. Of mice and not men: Differences between mouse and human immunology. J. Immunol. 2004, 72, 2731–2738. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophils at the crossroads of innate and adaptive immunity. J. Leukoc. Biol. 2020, 108, 377–396. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophil: A cell with many roles in inflammation or several cell types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef] [PubMed]
- Chavakis, E.; Choi, E.Y.; Chavakis, T. Novel aspects in the regulation of the leukocyte adhesion cascade. Thromb. Haemost. 2009, 102, 191–197. [Google Scholar] [CrossRef]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef]
- Borregaard, N. Neutrophils, from marrow to microbes. Immunity 2010, 33, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Nauseef, W.M. How human neutrophils kill and degrade microbes: An integrated view. Immunol. Rev. 2007, 219, 88–102. [Google Scholar] [CrossRef] [PubMed]
- Burn, G.L.; Foti, A.; Marsman, G.; Patel, D.F.; Zychlinsky, A. The neutrophil. Immunity 2021, 54, 1377–1391. [Google Scholar] [CrossRef]
- Mayadas, T.N.; Cullere, X.; Lowell, C.A. The multifaceted functions of neutrophils. Annu. Rev. Pathol. 2014, 9, 181–218. [Google Scholar] [CrossRef]
- Rosales, C.; Uribe-Querol, E. Phagocytosis: A fundamental process in immunity. Biomed. Res. Int. 2017, 2017, 9042851. [Google Scholar] [CrossRef]
- Brinkmann, V. Neutrophil extracellular traps in the second decade. J. Innate Immun. 2018, 10, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Malech, H.L.; Deleo, F.R.; Quinn, M.T. The role of neutrophils in the immune system: An overview. Methods Mol. Biol. 2014, 1124, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Freeman, S.A.; Grinstein, S. Phagocytosis: Receptors, signal integration, and the cytoskeleton. Immunol. Rev. 2014, 262, 193–215. [Google Scholar] [CrossRef]
- Lim, J.J.; Grinstein, S.; Roth, Z. Diversity and versatility of phagocytosis: Roles in innate immunity, tissue remodeling, and homeostasis. Front. Cell Infect. Microbiol. 2017, 7, 191. [Google Scholar] [CrossRef] [PubMed]
- Ramadass, M.; Catz, S.D. Molecular mechanisms regulating secretory organelles and endosomes in neutrophils and their implications for inflammation. Immunol. Rev. 2016, 273, 249–265. [Google Scholar] [CrossRef] [PubMed]
- Mol, S.; Hafkamp, F.M.J.; Varela, L.; Simkhada, N.; Taanman-Kueter, E.W.; Tas, S.W.; Wauben, M.H.M.; Kormelink, T.G.; de Jong, E.C. Efficient neutrophil activation requires two simultaneous activating stimuli. Int. J. Mol. Sci. 2021, 22, 10106. [Google Scholar] [CrossRef] [PubMed]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.C.; Gougerot-Pocidalo, M.A.; Dang, P.M. Priming of the neutrophil respiratory burst: Role in host defense and inflammation. Immunol. Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH oxidase activation and bacterial resistance. Front. Cell. Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef]
- Potera, R.M.; Jensen, M.J.; Hilkin, B.M.; South, G.K.; Hook, J.S.; Gross, E.A.; Moreland, J.G. Neutrophil azurophilic granule exocytosis is primed by TNF-α and partially regulated by NADPH oxidase. Innate Immun. 2016, 22, 635–646. [Google Scholar] [CrossRef]
- Wolach, B.; Gavrieli, R.; de Boer, M.; van Leeuwen, K.; Berger-Achituv, S.; Stauber, T.; Ben Ari, J.; Rottem, M.; Schlesinger, Y.; Grisaru-Soen, G.; et al. Chronic granulomatous disease: Clinical, functional, molecular, and genetic studies. The Israeli experience with 84 patients. Am. J. Hematol. 2017, 92, 28–36. [Google Scholar] [CrossRef]
- Roos, D. Chronic granulomatous disease. Br. Med. Bull. 2016, 118, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Veenith, T.; Martin, H.; Le Breuilly, M.; Whitehouse, T.; Gao-Smith, F.; Duggal, N.; Lord, J.M.; Mian, R.; Sarphie, D.; Moss, P. High generation of reactive oxygen species from neutrophils in patients with severe COVID-19. Sci. Rep. 2022, 12, 10484. [Google Scholar] [CrossRef]
- Dupré-Crochet, S.; Erard, M.; Nüβe, O. ROS production in phagocytes: Why, when, and where? J. Leukoc. Biol. 2013, 94, 657–670. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Kettle, A.J.; Hampton, M.B. Reactive Oxygen species and neutrophil function. Annu. Rev. Biochem. 2016, 85, 765–792. [Google Scholar] [CrossRef]
- Herb, M.; Schramm, M. Functions of ROS in macrophages and antimicrobial immunity. Antioxidants 2021, 10, 313. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Suh, J.K.; Jeong, J.S.; Cho, S.Y.; Kim, D.W. Antioxidant effect of lidocaine and procaine on reactive oxygen species-induced endothelial dysfunction in the rabbit abdominal aorta. Korean J. Anesthesiol. 2010, 59, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil elastase and myeloperoxidase regulate the formation of neutrophil extracellular traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef]
- Kasprzycka, W.; Homa-Mlak, I.; Mlak, R.; Małecka-Massalska, T. Direct and indirect methods of evaluating the NETosis process. J. Pre-Clin. Clin. Res. 2019, 13, 50–56. [Google Scholar] [CrossRef]
- Efrimescu, C.I.; Buggy, P.M.; Buggy, D.J. Neutrophil extracellular trapping role in cancer, metastases, and cancer-related thrombosis: A narrative review of the current evidence base. Curr. Oncol. Rep. 2021, 23, 118. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P. Molecular mechanisms of cell death: Recommendations of the nomenclature committee on cell death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Yipp, B.G.; Kubes, P. NETosis: How vital is it? Blood 2013, 122, 2784–2794. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, M.; Khan, M.A.; Palaniyar, N. Neutrophil extracellular trap formation: Physiology, pathology, and pharmacology. Biomolecules 2019, 9, 365. [Google Scholar] [CrossRef]
- Khan, M.A.; Palaniyar, N. Transcriptional firing helps to drive NETosis. Sci. Rep. 2017, 7, 41749. [Google Scholar] [CrossRef] [PubMed]
- Stoimenou, M.; Tzoros, G.; Skendros, P.; Chrysanthopoulou, A. Methods for the assessment of NET formation: From neutrophil biology to translational research. Int. J. Mol. Sci. 2022, 23, 15823. [Google Scholar] [CrossRef]
- De Buhr, N.; Von Köckritz-Blickwede, M. How neutrophil extracellular traps become visible. J. Immunol. Res. 2016, 2016, 4604713. [Google Scholar] [CrossRef]
- Condello, M.; Caraglia, M.; Castellano, M.; Arancia, G.; Meschini, S. Structural and functional alterations of cellular components as revealed by electron microscopy. Microsc. Res. Tech. 2013, 76, 1057–1069. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Chan, D.W.; Zaal, K.J.; Kaplan, M.J. A high-throughput real-time imaging technique to quantify NETosis and distinguish mechanisms of cell death in human neutrophils. J. Immunol. 2018, 200, 869–879. [Google Scholar] [CrossRef]
- Denning, N.L.; Aziz, M.; Gurien, S.D.; Wang, P. DAMPs and NETs in sepsis. Front. Immunol. 2019, 10, 2536. [Google Scholar] [CrossRef]
- Mikacenic, C.; Moore, R.; Dmyterko, V.; West, T.E.; Altemeier, W.A.; Liles, W.C.; Lood, C. Neutrophil extracellular traps (NETs) are increased in the alveolar spaces of patients with ventilator-associated pneumonia. Crit. Care 2018, 22, 358. [Google Scholar] [CrossRef]
- Lefrançais, E.; Mallavia, B.; Zhuo, H.; Calfee, C.S.; Looney, M.R. Maladaptive role of neutrophil extracellular traps in pathogen-induced lung injury. JCI Insight 2018, 3, e98178. [Google Scholar] [CrossRef] [PubMed]
- Maruchi, Y.; Tsuda, M.; Mori, H.; Takenaka, N.; Gocho, T.; Huq, M.A.; Takeyama, N. Plasma myeloperoxidase-conjugated DNA level predicts outcomes and organ dysfunction in patients with septic shock. Crit. Care 2018, 22, 176. [Google Scholar] [CrossRef] [PubMed]
- Szturmowicz, M.; Demkow, U. Neutrophil extracellular traps (NETs) in severe SARS-CoV-2 lung disease. Int. J. Mol. Sci. 2021, 22, 8854. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Jin, J. Neutrophil extracellular traps: New players in cancer research. Front. Immunol. 2022, 13, 937565. [Google Scholar] [CrossRef] [PubMed]
- Hahn, J.; Knopf, J.; Maueröder, C.; Kienhöfer, D.; Leppkes, M.; Herrmann, M. Neutrophils and neutrophil extracellular traps orchestrate initiation and resolution of inflammation. Clin. Exp. Rheumatol. 2016, 34, 6–8. [Google Scholar]
- Kaltenmeier, C.; Yazdani, H.O.; Morder, K.; Geller, D.A.; Simmons, R.L.; Tohme, S. Neutrophil extracellular traps promote T cell exhaustion in the tumor microenvironment. Front. Immunol. 2021, 12, 785222. [Google Scholar] [CrossRef]
- Jiang, Z.Z.; Peng, Z.P.; Liu, X.C.; Guo, H.F.; Zhou, M.M.; Jiang, D.; Ning, W.R.; Huang, Y.F.; Zheng, L.; Wu, Y. Neutrophil extracellular traps induce tumor metastasis through dual effects on cancer and endothelial cells. Oncoimmunology 2022, 11, 2052418. [Google Scholar] [CrossRef]
- Thålin, C.; Lundström, S.; Seignez, C.; Daleskog, M.; Lundström, A.; Henriksson, P.; Helleday, T.; Phillipson, M.; Wallén, H.; Demers, M. Citrullinated histone H3 as a novel prognostic blood marker in patients with advanced cancer. PLoS ONE 2018, 13, e0191231. [Google Scholar] [CrossRef]
- Snoderly, H.T.; Boone, B.A.; Bennewitz, M.F. Neutrophil extracellular traps in breast cancer and beyond: Current perspectives on NET stimuli, thrombosis and metastasis, and clinical utility for diagnosis and treatment. Breast Cancer Res. 2019, 21, 145. [Google Scholar] [CrossRef]
- Albrengues, J.; Shields, M.A.; Ng, D.; Park, C.G.; Ambrico, A.; Poindexter, M.E.; Upadhyay, P.; Uyeminami, D.L.; Pommier, A.; Küttner, V.; et al. Neutrophil extracellular traps produced during inflammation awaken dormant cancer cells in mice. Science 2018, 361, eaao4227. [Google Scholar] [CrossRef]
- Lee, W.; Ko, S.Y.; Mohamed, M.S.; Kenny, H.A.; Lengyel, E.; Naora, H. Neutrophils facilitate ovarian cancer premetastatic niche formation in the omentum. J. Exp. Med. 2019, 216, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Cristinziano, L.; Modestino, L.; Antonelli, A.; Marone, G.; Simon, H.U.; Varricchi, G.; Galdiero, M.R. Neutrophil extracellular traps in cancer. Semin. Cancer Biol. 2022, 79, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Wysocki, R.W.; Amoozgar, Z.; Maiorino, L.; Fein, M.R.; Jorns, J.; Schott, A.F.; Kinugasa-Katayama, Y.; Lee, Y.; Won, N.H.; et al. Cancer cells induce metastasis-supporting neutrophil extracellular DNA traps. Sci. Transl. Med. 2016, 8, 361ra138. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Q.; Zhang, X.; Liu, X.; Zhou, B.; Chen, J.; Huang, D.; Li, J.; Li, H.; Chen, F.; et al. DNA of neutrophil extracellular traps promotes cancer metastasis via CCDC25. Nature 2020, 583, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Cong, M.; Li, J.; He, D.; Wu, Q.; Tian, P.; Wang, Y.; Yang, S.; Liang, C.; Liang, Y.; et al. Cathepsin C promotes breast cancer lung metastasis by modulating neutrophil infiltration and neutrophil extracellular trap formation. Cancer Cell 2021, 39, 423–437.e7. [Google Scholar] [CrossRef] [PubMed]
- Tamassia, N.; Bianchetto-Aguilera, F.; Arruda-Silva, F.; Gardiman, E.; Gasperini, S.; Calzetti, F.; Cassatella, M.A. Cytokine production by human neutrophils: Revisiting the “dark side of the moon”. Eur. J. Clin. Investig. 2018, 48, e12952. [Google Scholar] [CrossRef] [PubMed]
- Tecchio, C.; Scapini, P.; Pizzolo, G.; Cassatella, M.A. On the cytokines produced by human neutrophils in tumors. Semin. Cancer Biol. 2013, 23, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Gabrilovich, D. Cytokine production by neutrophils. In The Neutrophils, 3rd ed.; de Chaisemartin, L., Cassatella, M., Chollet-Martin, S., Eds.; Imperial College Press: London, UK, 2013; pp. 189–217. [Google Scholar]
- Buonacera, A.; Stancanelli, B.; Colaci, M.; Malatino, L. Neutrophil to lymphocyte ratio: An emerging marker of the relationships between the immune system and diseases. Int. J. Mol. Sci. 2022, 23, 3636. [Google Scholar] [CrossRef]
- Jiang, J.; Liu, R.; Yu, X.; Yang, R.; Xu, H.; Mao, Z.; Wang, Y. The neutrophil-lymphocyte count ratio as a diagnostic marker for bacteraemia: A systematic review and meta-analysis. Am. J. Emerg. Med. 2019, 37, 1482–1489. [Google Scholar] [CrossRef]
- Lee, M.J.; Park, S.D.; Kwon, S.W.; Woo, S.I.; Lee, M.D.; Shin, S.H.; Kim, D.H.; Kwan, J.; Park, K.S. Relation between neutrophil-to-lymphocyte ratio and index of microcirculatory resistance in patients with ST-segment elevation myocardial infarction undergoing primary percutaneous coronary intervention. Am. J. Cardiol. 2016, 118, 1323–1328. [Google Scholar] [CrossRef]
- Huang, Z.; Fu, Z.; Huang, W.; Huang, K. Prognostic value of neutrophil-to-lymphocyte ratio in sepsis: A meta-analysis. Am. J. Emerg. Med. 2020, 38, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Yoo, G.; Lee, T.; Uh, Y.; Kim, J. Identification of the robust predictor for sepsis based on clustering analysis. Sci. Rep. 2022, 12, 2336. [Google Scholar] [CrossRef] [PubMed]
- Hibino, S.; Kawazoe, T.; Kasahara, H.; Itoh, S.; Ishimoto, T.; Sakata-Yanagimoto, M.; Taniguchi, K. Inflammation-induced tumorigenesis and metastasis. Int. J. Mol. Sci. 2021, 22, 5421. [Google Scholar] [CrossRef] [PubMed]
- Templeton, A.J.; McNamara, M.G.; Šeruga, B.; Vera-Badillo, F.E.; Aneja, P.; Ocaña, A.; Leibowitz-Amit, R.; Sonpavde, G.; Knox, J.J.; Tran, B.; et al. Prognostic role of neutrophil-to-lymphocyte ratio in solid tumors: A systematic review and meta-analysis. J. Natl. Cancer Inst. 2014, 106, dju124. [Google Scholar] [CrossRef]
- Malietzis, G.; Giacometti, M.; Kennedy, R.H.; Athanasiou, T.; Aziz, O.; Jenkins, J.T. The emerging role of neutrophil to lymphocyte ratio in determining colorectal cancer treatment outcomes: A systematic review and meta-analysis. Ann. Surg. Oncol. 2014, 21, 3938–3946. [Google Scholar] [CrossRef] [PubMed]
- Silberman, S.; Abu-Yunis, U.; Tauber, R.; Shavit, L.; Grenader, T.; Fink, D.; Bitran, D.; Merin, O. Neutrophil-lymphocyte ratio: Prognostic impact in heart surgery. Early Outcomes Late Survival. Ann. Thorac. Surg. 2018, 105, 581–586. [Google Scholar] [CrossRef]
- Hajibandeh, S.; Hajibandeh, S.; Hobbs, N.; Mansour, M. Neutrophil-to-lymphocyte ratio predicts acute appendicitis and distinguishes between complicated and uncomplicated appendicitis: A systematic review and meta-analysis. Am. J. Surg. 2020, 219, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef]
- Schoppmann, S.F.; Birner, P.; Stöckl, J.; Kalt, R.; Ullrich, R.; Caucig, C.; Kriehuber, E.; Nagy, K.; Alitalo, K.; Kerjaschki, D. Tumor-associated macrophages express lymphatic endothelial growth factors and are related to peritumoral lymphangiogenesis. Am. J. Pathol. 2002, 161, 947–956. [Google Scholar] [CrossRef]
- Brandau, S.; Dumitru, C.A.; Lang, S. Protumor and antitumor functions of neutrophil granulocytes. Semin. Immunopathol. 2013, 35, 163–176. [Google Scholar] [CrossRef]
- Uribe-Querol, E.; Rosales, C. Neutrophils in Cancer: Two Sides of the Same Coin. J. Immunol. Res. 2015, 2015, 983698. [Google Scholar] [CrossRef] [PubMed]
- Houghton, A.M.; Rzymkiewicz, D.M.; Ji, H.; Gregory, A.D.; Egea, E.E.; Metz, H.E.; Stolz, D.B.; Land, S.R.; Marconcini, L.A.; Kliment, C.R.; et al. Neutrophil elastase-mediated degradation of IRS-1 accelerates lung tumor growth. Nat. Med. 2010, 16, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Zilio, S.; Serafini, P. Neutrophils and granulocytic MDSC: The janus God of cancer immunotherapy. Vaccines 2016, 4, 31. [Google Scholar] [CrossRef] [PubMed]
- Condamine, T.; Ramachandran, I.; Youn, J.I.; Gabrilovich, D.I. Regulation of tumor metastasis by myeloid-derived suppressor cells. Annu. Rev. Med. 2015, 66, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Tecchio, C.; Micheletti, A.; Cassatella, M.A. Neutrophil-derived cytokines: Facts beyond expression. Front. Immunol. 2014, 5, 508. [Google Scholar] [CrossRef] [PubMed]
- Wigmore, T.J.; Mohammed, K.; Jhanji, S. Long-term survival for patients undergoing volatile versus iv anesthesia for cancer surgery: A retrospective analysis. Anesthesiology 2016, 124, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Yap, A.; Lopez-Olivo, M.A.; Dubowitz, J.; Hiller, J.; Riedel, B. Global onco-anesthesia research collaboration group. In reply: Comment. on “Anesthetic technique and cancer outcomes: A meta-analysis of total intravenous versus volatile anesthesia”. Can. J. Anaesth. 2020, 67, 152–153. [Google Scholar] [CrossRef] [PubMed]
- Hasselager, R.P.; Hallas, J.; Gögenur, I. Inhalation or total intravenous anaesthesia and recurrence after colorectal cancer surgery: A propensity score matched Danish registry-based study. Br. J. Anaesth. 2021, 126, 921–930. [Google Scholar] [CrossRef]
- Che, X.; Li, T. Total versus inhaled intravenous anesthesia methods for prognosis of patients with lung, breast, or esophageal cancer: A cohort study. Front. Surg. 2023, 10, 1155351. [Google Scholar] [CrossRef]
- Meier, A.; Nizet, V. Impact of anesthetics on human neutrophil function. Anesth. Analg. 2019, 128, 569–574. [Google Scholar] [CrossRef]
- Lingamaneni, R.; Krasowski, M.D.; Jenkins, A.; Truong, T.; Giunta, A.L.; Blackbeer, J.; MacIver, M.B.; Harrison, N.L.; Hemmings, H.C., Jr. Anesthetic properties of 4-iodopropofol: Implications for mechanisms of anesthesia. Anesthesiology 2001, 94, 1050–1057. [Google Scholar] [CrossRef]
- Yi, S.; Tao, X.; Wang, Y.; Cao, Q.; Zhou, Z.; Wang, S. Effects of propofol on macrophage activation and function in diseases. Front. Pharmacol. 2022, 13, 964771. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.; Axtell, R.; Mitra, A.; Miranda, M.; Lock, C.; Tsien, R.W.; Steinman, L. Inhibitory role for GABA in autoimmune inflammation. Proc. Natl. Acad. Sci. USA 2010, 107, 2580–2585. [Google Scholar] [CrossRef]
- Haeseler, G.; Leuwer, M. High-affinity block of voltage-operated rat IIA neuronal sodium channels by 2,6 di-tert-butylphenol, a propofol analogue. Eur. J. Anaesthesiol. 2003, 20, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Kotani, Y.; Shimazawa, M.; Yoshimura, S.; Iwama, T.; Hara, H. The experimental and clinical pharmacology of propofol, an anesthetic agent with neuroprotective properties. CNS Neurosci. Ther. 2008, 14, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Krumholz, W.; Endrass, J.; Hempelmann, G. Propofol inhibits phagocytosis and killing of Staphylococcus aureus and Escherichia coli by polymorphonuclear leukocytes in vitro. Can. J. Anaesth. 1994, 41, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Heller, A.; Heller, S.; Blecken, S.; Urbaschek, R.; Koch, T. Effects of intravenous anesthetics on bacterial elimination in human blood in vitro. Acta Anaesthesiol. Scand. 1998, 42, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, D.C.; Margarit, S.C.; Hadade, A.N.; Mocan, T.N.; Miron, N.A.; Sessler, D.I. Choice of anesthetic technique on plasma concentrations of interleukins and cell adhesion molecules. Perioper. Med. 2013, 2, 8. [Google Scholar] [CrossRef] [PubMed]
- Koo, B.W.; Sim, J.B.; Shin, H.J.; Kim, D.W.; Kang, S.B.; Do, S.H.; Na, H.S. Surgical site infection after colorectal surgery according to the main anesthetic agent: A retrospective comparison between volatile anesthetics and propofol. Korean J. Anesthesiol. 2016, 69, 332–340. [Google Scholar] [CrossRef]
- Von Dossow, V.; Baur, S.; Sander, M.; Tønnesen, H.; Marks, C.; Paschen, C.; Berger, G.; Spies, C.D. Propofol increased the interleukin-6 to interleukin-10 ratio more than isoflurane after surgery in long-term alcoholic patients. J. Int. Med. Res. 2007, 35, 395–405. [Google Scholar] [CrossRef]
- Hughes, C.G.; Mailloux, P.T.; Devlin, J.W.; Swan, J.T.; Sanders, R.D.; Anzueto, A.; Jackson, J.C.; Hoskins, A.S.; Pun, B.T.; Orun, O.M.; et al. Dexmedetomidine or propofol for sedation in mechanically ventilated adults with sepsis. N. Engl. J. Med. 2021, 384, 1424–1436. [Google Scholar] [CrossRef]
- Sanders, R.D.; Grover, V.; Goulding, J.; Godlee, A.; Gurney, S.; Snelgrove, R.; Ma, D.; Singh, S.; Maze, M.; Hussell, T. Immune cell expression of GABAA receptors and the effects of diazepam on influenza infection. J. Neuroimmunol. 2015, 282, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.C.; Chung, P.J.; Ho, C.M.; Kuo, C.Y.; Hung, M.F.; Huang, Y.T.; Chang, W.Y.; Chang, Y.W.; Chan, K.H.; Hwang, T.L. Propofol inhibits superoxide production, elastase release, and chemotaxis in formyl peptide-activated human neutrophils by blocking formyl peptide receptor 1. J. Immunol. 2013, 190, 6511–5619. [Google Scholar] [CrossRef] [PubMed]
- Stahl, S.M. Mechanism of action of ketamine. CNS Spectr. 2013, 18, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.S.; Penna, A.; Orser, B.A. Ketamine increases the function of γ-aminobutyric acid type A receptors in hippocampal and cortical neurons. Anesthesiology 2017, 126, 666–677. [Google Scholar] [CrossRef]
- Zhou, C.; Douglas, J.E.; Kumar, N.N.; Shu, S.; Bayliss, D.A.; Chen, X. Forebrain HCN1 channels contribute to hypnotic actions of ketamine. Anesthesiology 2013, 118, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Zilberstein, G.; Levy, R.; Rachinsky, M.; Fisher, A.; Greemberg, L.; Shapira, Y.; Appelbaum, A.; Roytblat, L. Ketamine attenuates neutrophil activation after cardiopulmonary bypass. Anesth. Analg. 2002, 95, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Ghori, K.; O’Driscoll, J.; Shorten, G. The effect of midazolam on neutrophil mitogen-activated protein kinase. Eur. J. Anaesthesiol. 2010, 27, 562–565. [Google Scholar] [CrossRef]
- Marino, F.; Cattaneo, S.; Cosentino, M.; Rasini, E.; Di Grazia, L.; Fietta, A.M.; Lecchini, S.; Frigo, G. Diazepam stimulates migration and phagocytosis of human neutrophils: Possible contribution of peripheral-type benzodiazepine receptors and intracellular calcium. Pharmacology 2001, 63, 42–49. [Google Scholar] [CrossRef]
- Herrera-García, A.M.; Domínguez-Luis, M.J.; Arce-Franco, M.; Armas-González, E.; Álvarez de La Rosa, D.; Machado, J.D.; Pec, M.K.; Feria, M.; Barreiro, O.; Sánchez-Madrid, F.; et al. Prevention of neutrophil extravasation by α2-adrenoceptor-mediated endothelial stabilization. J. Immunol. 2014, 193, 3023–3035. [Google Scholar] [CrossRef]
- Chen, R.; Sun, Y.; Lv, J.; Dou, X.; Dai, M.; Sun, S.; Lin, Y. Effects of dexmedetomidine on immune cells: A narrative review. Front. Pharmacol. 2022, 13, 829951. [Google Scholar] [CrossRef] [PubMed]
- Carnet Le Provost, K.; Kepp, O.; Kroemer, G.; Bezu, L. Trial watch: Dexmedetomidine in cancer therapy. Oncoimmunology 2024, 13, 2327143. [Google Scholar] [CrossRef] [PubMed]
- Franks, N.P.; Lieb, W.R. Molecular and cellular mechanisms of general anaesthesia. Nature 1994, 367, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liang, P.; Liu, J.; Ke, B.; Wang, X.; Li, F.; Li, T.; Bayliss, D.A.; Chen, X. HCN1 channels contribute to the effects of amnesia and hypnosis but not immobility of volatile anesthetics. Anesth. Analg. 2015, 121, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Downie, D.L.; Hall, A.C.; Lieb, W.R.; Franks, N.P. Effects of inhalational general anaesthetics on native glycine receptors in rat medullary neurones and recombinant glycine receptors in Xenopus oocytes. Br. J. Pharmacol. 1996, 118, 493–502. [Google Scholar] [CrossRef]
- Stollings, L.M.; Jia, L.J.; Tang, P.; Dou, H.; Lu, B.; Xu, Y. Immune modulation by volatile anesthetics. Anesthesiology 2016, 125, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, D.; Rothe, G.; Schwall, B.; Schmid, P.; Schmitz, G.; Taeger, K.; Hobbhahn, J. Effects of volatile anaesthetics on human neutrophil oxidative response to the bacterial peptide FMLP. Br. J. Anaesth. 1997, 78, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Möbert, J.; Zahler, S.; Becker, B.F.; Conzen, P.F. Inhibition of neutrophil activation by volatile anesthetics decreases adhesion to cultured human endothelial cells. Anesthesiology 1999, 90, 1372–1381. [Google Scholar] [CrossRef] [PubMed]
- de Rossi, L.W.; Horn, N.A.; Hecker, K.E.; Robitzsch, T.; Hutschenreuter, G.; Rossaint, R. Effect of halothane and isoflurane on binding of ADP- and TRAP-6- activated platelets to leukocytes in Whole blood. Anesthesiology 2002, 96, 117–124. [Google Scholar] [CrossRef]
- Du, X.; Jiang, C.; Lv, Y.; Dull, R.O.; Zhao, Y.Y.; Schwartz, D.E.; Hu, G. Isoflurane promotes phagocytosis of apoptotic neutrophils through AMPK-mediated ADAM17/Mer signaling. PLoS ONE 2017, 12, e0180213. [Google Scholar] [CrossRef]
- Kong, H.Y.; Zhu, S.M.; Wang, L.Q.; He, Y.; Xie, H.Y.; Zheng, S.S. Sevoflurane protects against acute kidney injury in a small-size liver transplantation model. Am. J. Nephrol. 2010, 32, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Alam, A.; Rampes, S.; Patel, S.; Hana, Z.; Ma, D. Anesthetics or anesthetic techniques and cancer surgical outcomes: A possible link. Korean J. Anesthesiol. 2021, 74, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Buckley, A.; McQuaid, S.; Johnson, P.; Buggy, D.J. Effect of anaesthetic technique on the natural killer cell anti-tumour activity of serum from women undergoing breast cancer surgery: A pilot study. Br. J. Anaesth. 2014, 113, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Kim, R. Effects of surgery and anesthetic choice on immunosuppression and cancer recurrence. J. Transl. Med. 2018, 16, 8. [Google Scholar] [CrossRef] [PubMed]
- Tikhonov, D.B.; Zhorov, B.S. Mechanism of sodium channel block by local anesthetics, antiarrhythmics, and anticonvulsants. J. Gen. Physiol. 2017, 149, 465–481. [Google Scholar] [CrossRef] [PubMed]
- de Lera Ruiz, M.; Kraus, R.L. Voltage-gated sodium channels: Structure, function, pharmacology, and clinical indications. J. Med. Chem. 2015, 58, 7093–7118. [Google Scholar] [CrossRef]
- Kulińska, K.I.; Szałkowska, S.; Andrusiewicz, M.; Kotwicka, M.; Billert, H. The effect of local anaesthetics on apoptosis and NETosis of human neutrophils in vitro: Comparison between lidocaine and ropivacaine. Hum. Cell 2023, 36, 2027–2039. [Google Scholar] [CrossRef]
- Galoș, E.V.; Tat, T.F.; Popa, R.; Efrimescu, C.I.; Finnerty, D.; Buggy, D.J.; Ionescu, D.C.; Mihu, C.M. Neutrophil extracellular trapping and angiogenesis biomarkers after intravenous or inhalation anaesthesia with or without intravenous lidocaine for breast cancer surgery: A prospective, randomised trial. Br. J. Anaesth. 2020, 125, 712–721. [Google Scholar] [CrossRef]
- Kulinska, K.I.; Billert, M.; Sawinski, K.; Czerniak, K.; Gaca, M.; Kusza, K.; Nowak, K.W.; Siemionow, M.; Billert, H. Local anaesthetics upregulate nitric oxide generation in cord blood and adult human neutrophils. Sci. Rep. 2019, 9, 569. [Google Scholar] [CrossRef]
- Castro, I.; Carvalho, P.; Vale, N.; Monjardino, T.; Mourão, J. Systemic anti-inflammatory effects of intravenous lidocaine in surgical patients: A systematic review and meta-analysis. J. Clin. Med. 2023, 12, 3772. [Google Scholar] [CrossRef]
- Zhong, J.; Hu, J.; Mao, L.; Ye, G.; Qiu, K.; Zhao, Y.; Hu, S. Efficacy of intravenous lidocaine for pain relief in the emergency department: A systematic review and meta-analysis. Front. Med. 2022, 8, 706844. [Google Scholar] [CrossRef] [PubMed]
- Brejchova, J.; Holan, V.; Svoboda, P. Expression of opioid receptors in cells of the immune system. Int. J. Mol. Sci. 2020, 22, 315. [Google Scholar] [CrossRef] [PubMed]
- Boland, J.W.; Pockley, A.G. Influence of opioids on immune function in patients with cancer pain: From bench to bedside. Br. J. Pharmacol. 2018, 175, 2726–2736. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Ninkovic, J.; Banerjee, S.; Charboneau, R.G.; Das, S.; Dutta, R.; Kirchner, V.A.; Koodie, L.; Ma, J.; Meng, J.; et al. Opioid drug abuse and modulation of immune function: Consequences in the susceptibility to opportunistic infections. J. Neuroimmune Pharmacol. 2011, 6, 442–465. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.; Henderson, G.; Bailey, C.P. Emerging areas of opioid pharmacology. Br. J. Pharmacol. 2018, 175, 2715–2716. [Google Scholar] [CrossRef]
- Brogi, E.; Forfori, F. Anesthesia and cancer recurrence: An overview. J. Anesth. Analg. Crit. Care 2022, 2, 33. [Google Scholar] [CrossRef] [PubMed]
- Díaz-González, F.; Sánchez-Madrid, F. NSAIDs: Learning new tricks from old drugs. Eur. J. Immunol. 2015, 45, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Venuta, A.; Nasso, R.; Gisonna, A.; Iuliano, R.; Montesarchio, S.; Acampora, V.; Sepe, L.; Avagliano, A.; Arcone, R.; Arcucci, A.; et al. Celecoxib, a non-steroidal anti-inflammatory drug, exerts a toxic effect on human melanoma cells grown as 2D and 3D cell cultures. Life 2023, 13, 1067. [Google Scholar] [CrossRef] [PubMed]
- Tsoi, K.K.F.; Ho, J.M.W.; Chan, F.C.H.; Sung, J.J.Y. Long-term use of low-dose aspirin for cancer prevention: A 10-year population cohort study in Hong Kong. Int. J. Cancer 2019, 145, 267–273. [Google Scholar] [CrossRef]
- Hall, C.J.; Wicker, S.M.; Chien, A.T.; Tromp, A.; Lawrence, L.M.; Sun, X.; Krissansen, G.W.; Crosier, K.E.; Crosier, P.S. Repositioning drugs for inflammatory disease—Fishing for new anti-inflammatory agents. Dis. Model. Mech. 2014, 7, 1069–1081. [Google Scholar] [CrossRef]
- Sessler, D.; Pei, L.; Huang, Y.; Fleischmann, E.; Marhofer, P.; Kurz, A.; Mayers, D.B.; Meyer-Treschan, T.A.; Grady, M.; Tan, E.Y.; et al. Recurrence of breast cancer after regional or general anaesthesia: A randomised controlled trial. Lancet 2019, 394, 1807–1815. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.F.; Cata, J.P. Anesthesia techniques and long-term oncological outcomes. Front. Oncol. 2021, 11, 788918. [Google Scholar] [CrossRef] [PubMed]
- Murphy, O.; Forget, P.; Ma, D.; Buggy, D.J. Tumour excisional surgery, anaesthetic-analgesic techniques, and oncologic outcomes: A narrative review. Br. J. Anaesth. 2023, 131, 989–1001. [Google Scholar] [CrossRef] [PubMed]
- Ní Eochagáin, A.; Burns, D.; Riedel, B.; Sessler, D.I.; Buggy, D.J. The effect of anaesthetic technique during primary breast cancer surgery on neutrophil-lymphocyte ratio, platelet-lymphocyte ratio and return to intended oncological therapy. Anaesthesia 2018, 73, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Memary, E.; Mirkheshti, A.; Ghasemi, M.; Taheri, M.; Dolatabadi, A.; Kaboudvand, A. The effect of lidocaine infusion during general anesthesia on neutrophil-lymphocyte-ratio in breast cancer patient’s candidate for mastectomy; a clinical trial. J. Cell Mol. Anesth. 2016, 1, 146–153. [Google Scholar]
- Alexa, A.L.; Ciocan, A.; Zaharie, F.; Valean, D.; Sargarovschi, S.; Breazu, C.; Al Hajjar, N.; Ionescu, D. The influence of intravenous lidocaine infusion on postoperative outcome and neutrophil-to-lymphocyte ratio in colorectal cancer patients. A pilot study. J. Gastrointestin. Liver Dis. 2023, 32, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Domagalska, M.; Ciftsi, B.; Janusz, P.; Reysner, T.; Kolasinski, J.; Wieczorowska-Tobis, K.; Kowalski, G. The neutrophil-to-lymphocyte ratio (NLR) and platelet-to-lymphocyte ratio (PLR) levels following erector spinae plane block (ESPB) in posterior lumbar decompression: A randomized, controlled trial. Eur. Spine J. 2023, 32, 4192–4199. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.; Ninh, M.K.; Ashford, S.; Cornett, E.M.; Kaye, A.D.; Urits, I.; Viswanath, O. Anesthesia medications and interaction with chemotherapeutic agents. Oncol. Ther. 2021, 9, 121–138. [Google Scholar] [CrossRef]
- Jiang, S.; Liu, Y.; Huang, L.; Zhang, F.; Kang, R. Effects of propofol on cancer development and chemotherapy: Potential mechanisms. Eur. J. Pharmacol. 2018, 831, 46–51. [Google Scholar] [CrossRef]
- Wang, P.; Chen, J.; Mu, L.H.; Du, Q.H.; Niu, X.H.; Zhang, M.Y. Propofol inhibits invasion and enhances paclitaxel-induced apoptosis in ovarian cancer cells through the suppression of the transcription factor slug. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 1722–1729, Erratum in: Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 6446. [Google Scholar]
- Xing, W.; Chen, D.T.; Pan, J.H.; Chen, Y.H.; Yan, Y.; Li, Q.; Xue, R.F.; Yuan, Y.F.; Zeng, W.A. Lidocaine induces apoptosis and suppresses tumor growth in human hepatocellular carcinoma cells in vitro and in a xenograft model in vivo. Anesthesiology 2017, 126, 868–881. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
NLR’s clinical implications. CVD, cardiovascular disease. CHD, coronary heart disease. ACS, acute coronary syndrome. MACEs, major adverse cardiac events. DFS, disease-free survival. OS, overall survival. SIRS, systemic inflammatory response syndrome.
Figure 1.
NLR’s clinical implications. CVD, cardiovascular disease. CHD, coronary heart disease. ACS, acute coronary syndrome. MACEs, major adverse cardiac events. DFS, disease-free survival. OS, overall survival. SIRS, systemic inflammatory response syndrome.

Figure 2.
Influence of anesthetic drugs on neutrophils. ROS, reactive oxygen species. NETs, neutrophil extracellular traps. AINS, nonsteroidal anti-inflammatory drugs. Cross marks suggests inhibition, while arrows suggests stimulation.
Figure 2.
Influence of anesthetic drugs on neutrophils. ROS, reactive oxygen species. NETs, neutrophil extracellular traps. AINS, nonsteroidal anti-inflammatory drugs. Cross marks suggests inhibition, while arrows suggests stimulation.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alexa, A.L.; Sargarovschi, S.; Ionescu, D. Neutrophils and Anesthetic Drugs: Implications in Onco-Anesthesia. Int. J. Mol. Sci. 2024, 25, 4033. https://doi.org/10.3390/ijms25074033
AMA Style
Alexa AL, Sargarovschi S, Ionescu D. Neutrophils and Anesthetic Drugs: Implications in Onco-Anesthesia. International Journal of Molecular Sciences. 2024; 25(7):4033. https://doi.org/10.3390/ijms25074033
Chicago/Turabian StyleAlexa, Alexandru Leonard, Sergiu Sargarovschi, and Daniela Ionescu. 2024. "Neutrophils and Anesthetic Drugs: Implications in Onco-Anesthesia" International Journal of Molecular Sciences 25, no. 7: 4033. https://doi.org/10.3390/ijms25074033
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.