
A Narrative Review on Adipose Tissue and Overtraining: Shedding Light on the Interplay among Adipokines, Exercise and Overtraining



Abstract
1. Introduction
2. Results
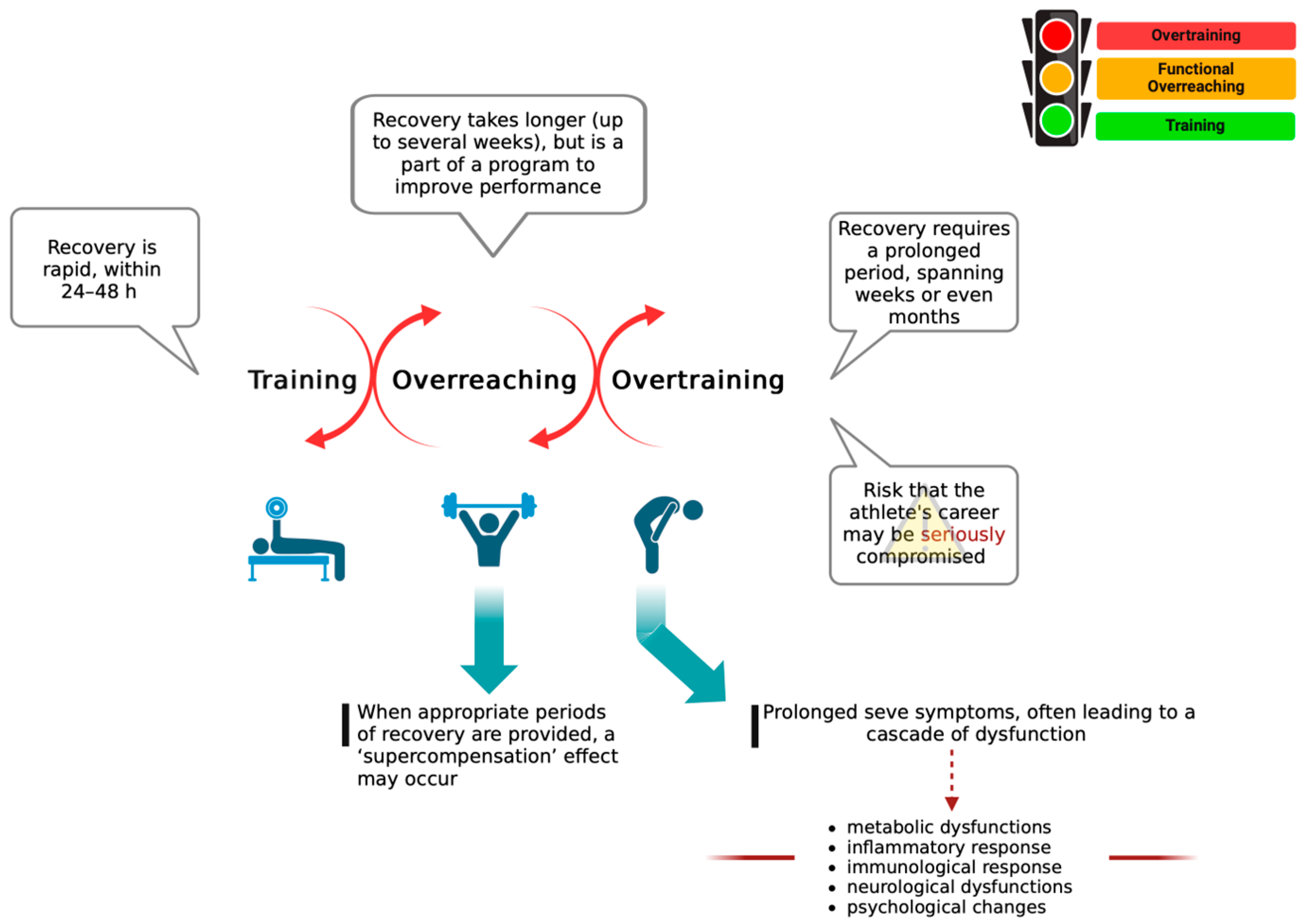
2.1. The Main Theories Underlying OTS
- -
- Glycogen Hypothesis: Insufficient muscle glycogen levels can significantly impair performance by depriving the muscles of sufficient fuel to sustain the workload. As glycogen serves as a primary energy source for muscles, its inadequate availability can hinder the ability to maintain intensity and prolong exercise duration, ultimately impacting overall performance and contributing to worsening fatigue [17,18]. Although low muscle glycogen levels may be linked to exercise-induced fatigue, the connection with OTS seems tenuous. Snyder et al. reported that even athletes with elevated carbohydrate consumption and normal glycogen levels can potentially develop OTS [18]. Furthermore, long-term carbohydrate supplementation in rats as an intervention to prevent or mitigate OTS attenuated OTS-induced performance decrements but did not reach statistical significance and was not able to protect against muscle damage [19].
- -
- Central Fatigue Theory: Fatigue is a complex phenomenon, with several interactions among central and peripheral factors. In the presence of prolonged and excessive training, concurrent with other stressors and insufficient recovery, performance decrements can result in chronic maladaptation that can lead to OTS [20]. OTS typically involves disturbances in mood, sleep, and behavior [2,21,22]. The main neurotransmitter implicated in the regulation of these functions is serotonin (5-HT) [23]. An increase in the synthesis of 5-hydroxytryptamine (5-HT) in the central nervous system (CNS) can give rise to OTS. Physical exercise diminishes the levels of BCAA by increasing their oxidation into glucose; this favors the entry of tryptophan into the brain, followed by its conversion into 5-HT [24]. Increased 5-HT synthesis in the brain has been positively correlated with fatigue [25,26]. Literature data showed how administering serotonin reuptake inhibitors to athletes increases 5-HT levels in the brain and diminishes performance [23]. In contrast, in marathon runners, supplementation with BCAA is related to improved physical and mental health; this is potentially attributed to a reduction in 5-HT synthesis [25].
- -
- Glutamine Hypothesis: There is substantial evidence demonstrating reductions in blood levels of the amino acid glutamine in OTS [27]. According to the glutamine theory, the frequently observed impaired immune response and associated increased rate of upper respiratory tract infections seen in OTS may be attributed to reduced blood glutamine levels; this is because glutamine is a primary fuel utilized by immune cells. Literature data have shown that extended periods of exercise can temporarily reduce plasma glutamine concentrations in athletes experiencing overtraining [28].
- -
- Oxidative Stress Hypothesis: Changes in redox homeostasis have been documented in athletes suffering from OTS [29]. During exercise, a certain level of oxidative stress is considered beneficial as reactive oxygen species (ROS) released from damaged muscles play a role in regulating cellular repair [30,31]. Additionally, exercise-induced ROS serves as a potent physiological stimulus to increase the expression of antioxidant enzymes, thereby enhancing antioxidant defense mechanisms [32]. However, excessive physical exercise can be detrimental for athletes, as it leads to a chronic and potentially dangerous elevation of ROS levels [33]. Athletes undergoing OTS seem to be more vulnerable to oxidative damage due to diminished responses to exercise-induced stress [33]. This increased vulnerability may result in oxidative stress becoming pathological, leading to inflammation and muscle fatigue, ultimately inhibiting athletic performance [33]. The relationship between the increased oxidative stress state and OTS remains still unclear, and there is limited clinically relevant research to elucidate whether it acts as a trigger or a consequence of OTS.
- -
- Autonomic Nervous System Hypothesis: An imbalance in the autonomic nervous system could explain some symptoms of OTS. Specifically, a reduction in sympathetic activation and an increase in parasympathetic dominance may contribute to performance inhibition, fatigue, and depression; reduced nocturnal urinary excretion of catecholamines is also observed [4]. Heart rate variability (HRV) is utilized as an indicator of autonomic function [4]. Studies employing HRV have suggested that the impact of intense training on automatic control can be reversible. Restoring a balance between sympathetic and parasympathetic forces may be attainable with a week of rest [7].
- -
- Hypothalamic Hypothesis: The hypothalamic hypothesis suggests that disruptions in the hypothalamic-pituitary-adrenal (HPA) and hypothalamic-pituitary-gonadal axes may contribute to OTS. Endurance athletes often exhibit changes in the function of the HPA axis, and overtrained athletes may experience variations in several hormones, such as cortisol, adrenocorticotropic hormone, and testosterone. However, the existing data about these hormonal changes are contradictory, and alterations in these axes are influenced by many factors, such as exercise capacity and other hormonal levels [34].
- -
- Muscle Damage Hypothesis: It is widely known that exhaustive exercise is linked with increased oxygen consumption in skeletal muscles, elevated lipid peroxidation, and inhibition of key mitochondrial enzymes [35]. Endurance training, when not properly balanced with adequate rest, typically does not lead to functional damage and promotes muscular maladaptation along with oxidative stress generation and a reduction in the muscle defense system [36]. Under normal training conditions, muscle fibers regenerate through the activation of satellite cells, resulting in the generation of new differentiated myofibers [37]. Conversely, continued intense training combined with inadequate rest can lead to a decrease in satellite cell numbers [38], reducing the formation of new fibers and the appropriate regeneration of damaged ones, thus contributing to the development of OTS.
- -
- Cytokine Hypothesis: The cytokine hypothesis states that the process of muscle contraction and repetitive joint action leads to tissue microtraumas, initiating physiological adaptation via the activation of local inflammatory response and recruitment of cytokines and interleukins such as IL-1 beta, IL-6, and TNF-α [39]. Continued rigorous training coupled with insufficient rest can exacerbate this inflammatory response, leading to a chronic and pathological condition [40]. This theory has been extensively supported by several literature studies. High levels of pro-inflammatory cytokines such as IL-1 beta, IL-6, and TNF-α are also linked to an increased restriction of food intake and alteration of cellular catabolism, leading to a reduction in glycogen stores [41]. In addition, some cytokines induce a reduction of GLUT-4 transporters in stressed muscles, interfering with glucose transport for glycogen synthesis. This process may contribute to excessive muscular fatigue in overtrained athletes [42].
2.2. Uncovering OTS Markers: Biochemical, Immunological, and Hormonal Responses
- -
- Biochemical markers, including creatine kinase (CK), urea, uric acid, ammonia, blood lactate, and plasma glutamine, have undergone extensive study across various athletes [57,58]. These markers are already used in many areas of sport to evaluate an athlete’s training response, helping coaches to identify workload and prevent injuries [59]. CK activity reflects both the intensity and volume of exercise. It is important to note that some athletes may show minimal increases in CK activity, making elevated CK activity alone insufficient to conclusively indicate OTS [60]. With regards to urea, increased concentrations may occur in response to elevated training loads, particularly in intensive endurance training, due to enhanced protein catabolism and gluconeogenesis [61]. Ammonia levels decrease at rest in athletes with OTS, although this finding is inconsistent, while uric acid levels do not show significant changes in overtrained athletes [62]. Marked responses were also observed in muscle damage markers such as blood lactate [63]. Lactate is typically released after exhaustive exercises and in smaller amounts during mild to moderate activities. Its levels are exponentially proportional to the level of activity. While several studies have reported reduced lactate levels in OTS along with impaired anaerobic lactate performance, Cadegiani et al. found increased lactate levels in OTS, likely due to impaired muscle recovery [34]. However, analyzing lactate levels post-exercise precludes the examination of lactate clearance and dynamic metabolism during rest periods. Nevertheless, numerous other metabolic variables also influence blood lactate, making it ineffective in identifying OTS on its own. Regarding glutamine levels, it has been reported that overtrained athletes exhibit low levels of plasma glutamine [6]. Changes in glutamine can also occur after physical trauma, inflammation, and infection [6]. Furthermore, plasma glutamine is also influenced by diet; in fact, it temporarily increases after the consumption of protein-rich foods [6]. Thus, the debate remains ongoing regarding whether glutamine can be reliably used as a marker of impending overtraining.
- -
- Oxygen free radicals generated during exercise contribute to muscular fatigue and are implicated in muscle damage [30]. Overtrained athletes presented higher indices of lipoperoxidation, such as thiobarbituric acid reactive substances (TBARS) and the blood ratio of reduced vs. oxidized glutathione (GSH/GSSG ratio) [57]. Literature data reported that continuous submaximal training led to a decrease in GSH and an increase in GSSG, followed by a decrease in the GSH:GSSG ratio [64]. Generally, athletes who experienced OTS exhibit higher exercise-induced variations also in TBARS levels compared to well-trained ones [4]. Further studies are needed to determine if these indices could serve as valuable markers of OTS.
- -
- Immunological markers are highly responsive to various forms of stress, including both physiological and psychological stressors. Athletes experiencing OTS are frequently reported to be immunosuppressed. As a result, immune parameters could potentially serve as indicators of stress levels in the context of exercise training. Recent investigations have focused on salivary immunoglobulin A (IgA) even if findings regarding salivary IgA levels in OTS athletes have been inconsistent: one study documented a significant reduction in salivary IgA levels among athletes experiencing overtraining symptoms [63], Halson SL et al. did not observe a statistically significant decrease following intensified training in a group of cyclists [65]. On the other hand, exercise and training induce changes in circulating numbers of lymphocyte subsets [66]. With heavy training, the T-lymphocyte CD4+/CD8+ ratio typically decreases [67]. However, this ratio seems to be similar between athletes diagnosed with OTS and well-trained athletes [68]. Literature data revealed that the expression of certain proteins on the surface of T-lymphocytes may be more sensitive to differentiate between the OT athletes and healthy ones [69]. Specifically, the expression of CD45RO+ on CD4+ cells was significantly higher in athletes experiencing OTS compared to healthy and well-trained controls [70]. Using this indicator, OTS could be identified with high specificity; however, further in-depth studies are needed.
- -
- Hormonal markers are often considered potential tools for identifying OTS, yet they are susceptible to numerous confounding variables, such as diurnal and seasonal timing. Among the hormones most investigated in OTS are cortisol, adrenocorticotropic hormone (ACTH), and human growth hormone (HGH) [34]. Regarding serum cortisol levels concerning OTS, literature data are inconsistent. While some studies report no change, others indicate increases, decreases, or variable responses. Evidence also suggests ACTH and GH deficiency in athletes with OTS. Specifically, ACTH and GH responses to insulin-induced were found to be lower in overtrained athletes compared to healthy, well-trained controls [56]. Unfortunately, current data are inconsistent in some cases, as they depend on various factors, including individual training capacity, intrinsic vulnerability to stressful factors, and other hormonal levels [71].
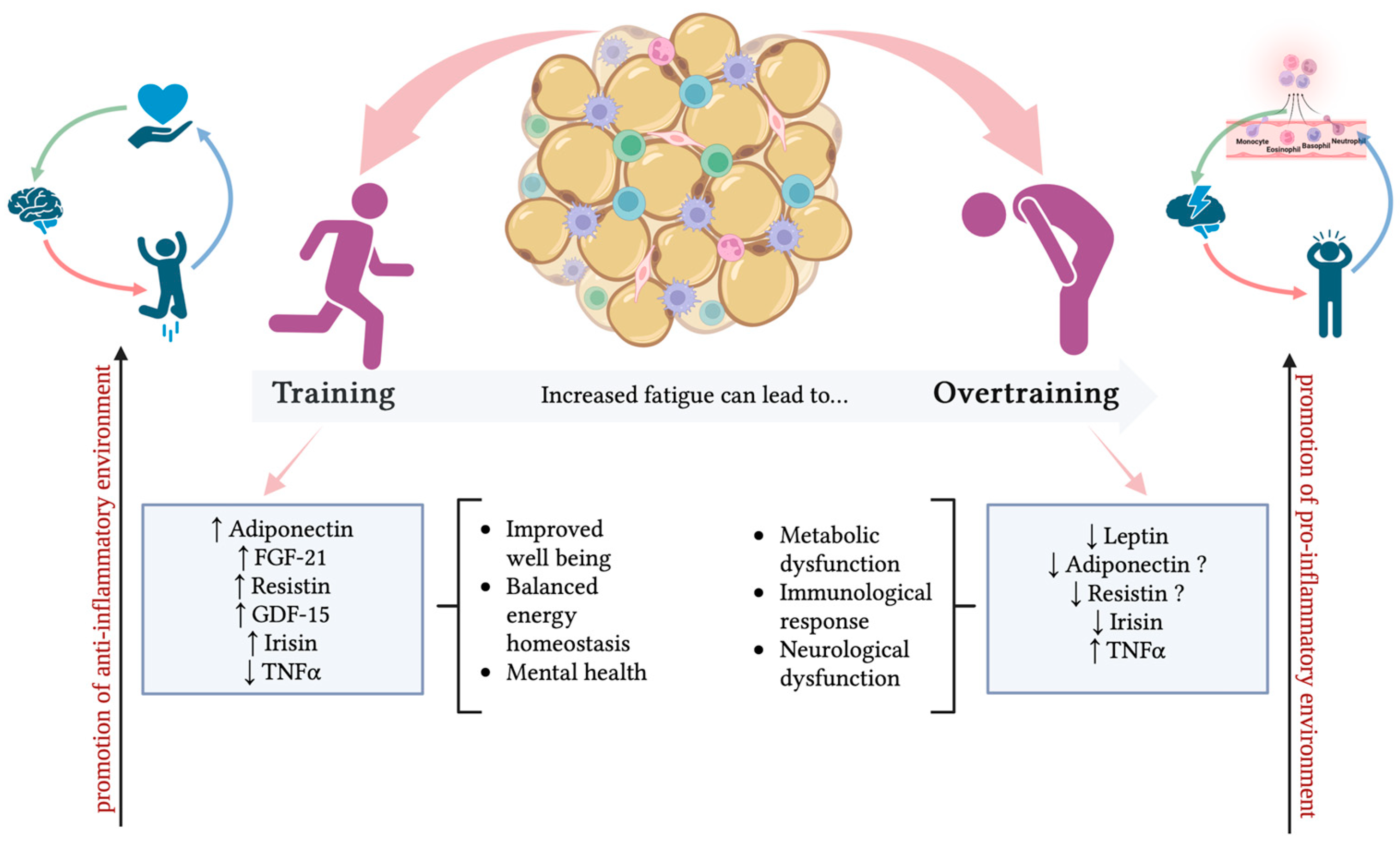
2.3. Adipose Tissue in Training, Reaching, and Overtraining
2.4. Regulation of the Secretion of Adipokines and Batokines
2.5. Adiponectin in Physical Activity and in OTS
2.6. Leptin in Physical Activity and OTS
2.7. Other Adipokines in Physical Activity and in OTS
- -
- TNF-α is a 26-kDa transmembrane multi-functional cytokine that, after a proteolytic cleavage, produces a 17-KDa soluble molecule. Biological functions include the regulation of many cellular and biological processes, such as immune function, cell differentiation, proliferation, apoptosis, and energy metabolism. Beyond macrophages as a well-known source of TNF-α, mature adipocytes and stromovascular fraction (SVF), which contains preadipocytes, endothelial cells, smooth muscle cells, fibroblasts, leukocytes, and macrophages in AT, also represent an important source of TNF-α production [138,139]. Generally, catabolic states are associated with lower levels of soluble circulating TNF-α compared to the obese state characterized by higher levels [140]. Secreted TNF-α then acts both on AT and peripheral tissues through the binding with two receptors, TNFR1 and 2 [138]. How exercise can modify the expression of AT-derived TNF-α is controversial. Regarding circulating levels, the response of TNFα to exercise largely depends on the intensity of training. Regular moderate exercise causes TNFα suppression through IL-6 [140,141], while results are different for acute exercise, such as marathon running, where TNF-α concentrations strongly increase [142]. Interestingly, the increase in TNF-α levels is higher with strenuous training caused by muscle damage during exercise [143]. Thus, in OTS, TNF-α is considered a key cytokine included in the “cytokine hypothesis” of OTS, and thus, these proinflammatory cytokines are significantly associated with measures of depressed mood, sleep disturbances, and stress in OTS athletes [144]. As said above, the cytokine hypothesis considers that an imbalance involving excessive exercise and inadequate recovery induces musculoskeletal trauma, increasing the production and release of proinflammatory cytokines, mainly IL-6, TNF-α, and IL-1beta, which interact with different organic systems, initiating most of the signs and symptoms linked to performance decrement [144].
- -
- Resistin is a cysteine-rich hormone secreted from white adipocytes and macrophages in humans. Resistin forms multimeric structures (hexamer and trimer) in circulation [146]. Resistin is involved in insulin resistance, but it is also involved in inflammatory responses in humans [147]. High resistin levels induce insulin resistance and exert proinflammatory effects [148].
- -
- Growth differentiation factor 15 (GDF-15) was first recognized in 1997 as a macrophage inhibitory cytokine-1 expressed in multiple tissues, including skeletal muscles [155]. Nowadays, it is known that it is also produced by adipose tissue under stress conditions to maintain cell and tissue homeostasis [156]. Recent studies also suggested that GDF-15 functions as an exerkine secreted by AT, exhibiting a possible protective role in exercise-induced muscle injury or inflammation. Normally, circulating blood levels of GDF-15 are quite low but it can be measured before and after exercise to assess the exercise-induced stress response intensity of each individual. In detail, high GDF-15 levels have been associated with lower muscle mass and strength, frailty, and declining physical function [157,158,159,160]. Tchou et al. found a strong increase in plasma GDF15 at the end of an ultramarathon foot race in male athletes [161]. In professional rugby players, circulating GDF15 was significantly increased after a session of intense training [162], and an increase in serum GDF15 was also observed in football players of the Spanish Football League 12 h after a match [163]. Recently, plasma GDF15 concentrations were found to gradually increase during and after a controlled vigorous submaximal exercise in young, healthy males [164].
- -
- Irisin is a cytokine produced by both muscle and AT during exercise [168]. Irisin is considered a thermogenic adipomyokine, able to improve glucose and lipid metabolism and ameliorate the effects of obesity-driven inflammation [169]. Irisin levels can be induced by exercise, both acute and resistance [170]. Irisin expression is regulated by the transcriptional coactivator peroxisome proliferator-activated receptor-gamma coactivator 1 alpha (PGC1-α) [171]. An increase in PGC1-α expression in the muscle of transgenic mice is associated with WAT browning in a process mediated by irisin [170]. Additionally, PGC1-α is associated with exercise-mediated benefits and energy metabolism, leading to the hypothesis that exercise may stimulate irisin release [170].
3. Discussion
4. Methods
Author Contributions
Funding
Conflicts of Interest
References
- Haugen, T.; Seiler, S.; Sandbakk, Ø.; Tønnessen, E. The Training and Development of Elite Sprint Performance: An Integration of Scientific and Best Practice Literature. Sports Med. Open 2019, 5, 44. [Google Scholar] [CrossRef] [PubMed]
- Kreher, J.B.; Schwartz, J.B. Overtraining Syndrome: A Practical Guide. Sports Health 2012, 4, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Meeusen, R.; Duclos, M.; Foster, C.; Fry, A.; Gleeson, M.; Nieman, D.; Raglin, J.; Rietjens, G.; Steinacker, J.; Urhausen, A.; et al. Prevention, diagnosis, and treatment of the overtraining syndrome: Joint consensus statement of the European College of Sport Science and the American College of Sports Medicine. Med. Sci. Sports Exerc. 2013, 45, 186–205. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, L.E.; VanHeest, J.L. The unknown mechanism of the overtraining syndrome: Clues from depression and psychoneuroimmunology. Sports Med. 2002, 32, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.G.; Potter, A.W.; Brooks, K.A. Overtraining Syndrome: Causes, Consequences, and Methods for Prevention. J. Sport Hum. Perform. 2014, 2, 1–14. [Google Scholar]
- Gleeson, M. Biochemical and immunological markers of over-training. J. Sports Sci. Med. 2002, 1, 31–41. [Google Scholar] [PubMed]
- Smith, L.L. Cytokine hypothesis of overtraining: A physiological adaptation to excessive stress? Med. Sci. Sports Exerc. 2000, 32, 317–331. [Google Scholar] [CrossRef] [PubMed]
- MacKinnon, L.T. Special feature for the Olympics: Effects of exercise on the immune system: Overtraining effects on immunity and performance in athletes. Immunol. Cell Biol. 2000, 78, 502–509. [Google Scholar] [CrossRef]
- Mika, A.; Macaluso, F.; Barone, R.; Di Felice, V.; Sledzinski, T. Effect of Exercise on Fatty Acid Metabolism and Adipokine Secretion in Adipose Tissue. Front. Physiol. 2019, 10, 26. [Google Scholar] [CrossRef]
- Becic, T.; Studenik, C.; Hoffmann, G. Exercise Increases Adiponectin and Reduces Leptin Levels in Prediabetic and Diabetic Individuals: Systematic Review and Meta-Analysis of Randomized Controlled Trials. Med. Sci. 2018, 6, 97. [Google Scholar] [CrossRef]
- Morais, G.P.; Chemerka, C.; Masson, A.; Seerattan, R.-A.; da Rocha, A.L.; Krawetz, R.; da Silva, A.S.R.; Herzog, W. Excessive downhill training leads to early onset of knee osteoarthritis. Osteoarthr. Cartil. 2021, 29, 870–881. [Google Scholar] [CrossRef] [PubMed]
- Docherty, S.; Harley, R.; McAuley, J.J.; Crowe, L.A.N.; Pedret, C.; Kirwan, P.D.; Siebert, S.; Millar, N.L. The effect of exercise on cytokines: Implications for musculoskeletal health: A narrative review. BMC Sports Sci. Med. Rehabil. 2022, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhou, H.; Zhao, C.; He, H. Effect of Exercise Training on Body Composition and Inflammatory Cytokine Levels in Overweight and Obese Individuals: A Systematic Review and Network Meta-Analysis. Front. Immunol. 2022, 13, 921085. [Google Scholar] [CrossRef] [PubMed]
- Berriel, G.P.; Costa, R.R.; da Silva, E.S.; Schons, P.; de Vargas, G.D.; Peyré-Tartaruga, L.A.; Kruel, L.F.M. Stress and recovery perception, creatine kinase levels, and performance parameters of male volleyball athletes in a preseason for a championship. Sports Med. Open 2020, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, L.E.; Bergeron, M.F.; Lee, E.C.; Mershon, J.E.; Armstrong, E.M. Overtraining Syndrome as a Complex Systems Phenomenon. Front. Netw. Physiol. 2022, 1, 794392. [Google Scholar] [CrossRef] [PubMed]
- Cadegiani, F.A.; Kater, C.E. Novel causes and consequences of overtraining syndrome: The EROS-DISRUPTORS study. BMC Sports Sci. Med. Rehabil. 2019, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Costill, D.L.; Flynn, M.G.; Kirwan, J.P.; Houmard, J.A.; Mitchell, J.B.; Thomas, R.; Park, S.H. Effects of repeated days of intensified training on muscle glycogen and swimming performance. Med. Sci. Sports Exerc. 1988, 20, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.J.; Jude, B.; Lanner, J.T. Intramuscular mechanisms of overtraining. Redox Biol. 2020, 35, 101480. [Google Scholar] [CrossRef] [PubMed]
- Snyder, A.C.; Kuipers, H.; Cheng, B.; Servais, R.; Fransen, E. Overtraining following intensified training with normal muscle glycogen. Med. Sci. Sports Exerc. 1995, 27, 1063–1070. [Google Scholar] [CrossRef]
- Meeusen, R.; Watson, P.; Hasegaw, H.; Roelands, B.; Piacentini, M.F. Brain neurotransmitters in fatigue and overtraining. Appl. Physiol. Nutr. Metab. 2007, 32, 857–864. [Google Scholar] [CrossRef]
- Campbell, E.H.; Poudevigne, M.; McFarlane, S.; Dilworth, L.; Irving, R. Evidence That Sleep Is an Indicator of Overtraining during the Competition Phase of Adolescent Sprinters. J. Sports Med. 2021, 2021, 6694547. [Google Scholar] [CrossRef] [PubMed]
- Kellmann, M. Preventing overtraining in athletes in high-intensity sports and stress/recovery monitoring. Scand. J. Med. Sci. Sports 2010, 20, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Heijnen, S.; Hommel, B.; Kibele, A.; Colzato, L.S. Neuromodulation of Aerobic Exercise—A Review. Front. Psychol. 2015, 6, 1890. [Google Scholar] [CrossRef] [PubMed]
- Budgett, R.; Hiscock, N.; Arida, R.M.; Castell, L.M. The effects of the 5-HT2C agonist m-chlorophenylpiperazine on elite athletes with unexplained underperformance syndrome (overtraining). Br. J. Sports Med. 2010, 44, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, M.; Foster, C.; Keul, J. Overtraining in endurance athletes: A brief review. Med. Sci. Sports Exerc. 1993, 25, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Castell, L.M.; Poortmans, J.R.; Leclercq, R.; Brasseur, M.; Duchateau, J.; Newsholme, E.A. Some aspects of the acute phase response after a marathon race, and the effects of glutamine supplementation. Eur. J. Appl. Physiol. Occup. Physiol. 1997, 75, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Coqueiro, A.Y.; Macedo Rogero, M.; Tirapegui, J. Glutamine as an Anti-Fatigue Amino Acid in Sports Nutrition. Nutrients 2019, 11, 863. [Google Scholar] [CrossRef] [PubMed]
- Halson, S.L.; Jeukendrup, A.E. Does overtraining exist? An analysis of overreaching and overtraining research. Sports Med. 2004, 34, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Tanskanen, M.; Atalay, M.; Uusitalo, A. Altered oxidative stress in overtrained athletes. J. Sports Sci. 2010, 28, 309–317. [Google Scholar] [CrossRef]
- Powers, S.K.; Ji, L.L.; Kavazis, A.N.; Jackson, M.J. Reactive oxygen species: Impact on skeletal muscle. Compr. Physiol. 2011, 1, 941–966. [Google Scholar]
- Bellafiore, M.; Bianco, A.; Battaglia, G.; Naccari, M.S.; Caramazza, G.; Padulo, J.; Chamari, K.; Paoli, A.; Palma, A. Training session intensity affects plasma redox status in amateur rhythmic gymnasts. J. Sport Health Sci. 2019, 8, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Viña, J.; Gimeno, A.; Sastre, J.; Desco, C.; Asensi, M.; Pallardó, F.V.; Cuesta, A.; Ferrero, J.A.; Terada, L.S.; Repine, J. Mechanism of Free Radical Production in Exhaustive Exercise in Humans and Rats; Role of Xanthine Oxidase and Protection by Allopurinol. IUBMB J. 2000, 49, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Margonis, K.; Fatouros, I.G.; Jamurtas, A.Z.; Nikolaidis, M.G.; Douroudos, I.; Chatzinikolaou, A.; Mitrakou, A.; Mastorakos, G.; Papassotiriou, I.; Taxildaris, K.; et al. Oxidative stress biomarkers responses to physical overtraining: Implications for diagnosis. Free Radic. Biol. Med. 2007, 43, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Cadegiani, F.A.; Kater, C.E. Hypothalamic-Pituitary-Adrenal (HPA) Axis Functioning in Overtraining Syndrome: Findings from Endocrine and Metabolic Responses on Overtraining Syndrome (EROS)—EROS-HPA Axis. Sports Med. Open 2017, 3, 45. [Google Scholar] [CrossRef] [PubMed]
- Radak, Z.; Zhao, Z.; Koltai, E.; Ohno, H.; Atalay, M. Oxygen Consumption and Usage during Physical Exercise: The Balance between Oxidative Stress and ROS-Dependent Adaptive Signaling. Antioxid. Redox Signal. 2013, 18, 1208–1246. [Google Scholar] [CrossRef] [PubMed]
- García-Pallarés, J.; Sánchez-Medina, L.; Carrasco, L.; Díaz, A.; Izquierdo, M. Endurance and neuromuscular changes in world-class level kayakers during a periodized training cycle. Eur. J. Appl. Physiol. 2009, 106, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Wagers, A.J.; Conboy, I.M. Cellular and molecular signatures of muscle regeneration: Current concepts and controversies in adult myogenesis. Cell 2005, 122, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Seene, T.; Kaasik, P. Role of exercise therapy in prevention of decline in aging muscle function: Glucocorticoid myopathy and unloading. J. Aging Res. 2012, 2012, 172492. [Google Scholar] [CrossRef][Green Version]
- Pichot, V.; Roche, F.; Gaspoz, J.M.; Enjolras, F.; Antoniadis, A.; Minini, P.; Costes, F.; Busso, T.; Lacour, J.R.; Barthélémy, J.C. Relation between heart rate variability and training load in middle-distance runners. Med. Sci. Sports Exerc. 2000, 32, 1729–1736. [Google Scholar] [CrossRef]
- Cerqueira, É.; Marinho, D.A.; Neiva, H.P.; Lourenço, O. Inflammatory Effects of High and Moderate Intensity Exercise—A Systematic Review. Front. Physiol. 2019, 10, 1550. [Google Scholar] [CrossRef]
- Snyder, A.C. Overtraining and glycogen depletion hypothesis. Med. Sci. Sports Exerc. 1998, 30, 1146–1150. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wang, J.; Hu, X.; Huang, X.J.; Chen, G. Current understanding of glucose transporter 4 expression and functional mechanisms. World J. Biol. Chem. 2020, 11, 76–98. [Google Scholar] [CrossRef] [PubMed]
- Rogeri, P.S.; Gasparini, S.O.; Martins, G.L.; Costa, L.K.F.; Araujo, C.C.; Lugaresi, R.; Kopfler, M.; Lancha, A.H. Crosstalk between Skeletal Muscle and Immune System: Which Roles Do IL-6 and Glutamine Play? Front. Physiol. 2020, 11, 582258. [Google Scholar] [CrossRef] [PubMed]
- Lotrich, F.E. Inflammatory cytokine-associated depression. Brain Res. 2015, 1617, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Greiwe, J.S.; Hickner, R.C.; Hansen, P.A.; Racette, S.B.; Chen, M.M.; Holloszy, J.O.; Goyaram, V.; Kohn, T.A.; Ojuka, E.O.; Schiavon, M.; et al. Effects of endurance exercise training on muscle glycogen accumulation in humans. J. Appl. Physiol. 1999, 87, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Hearris, M.A.; Hammond, K.M.; Fell, J.M.; Morton, J.P. Regulation of Muscle Glycogen Metabolism during Exercise: Implications for Endurance Performance and Training Adaptations. Nutrients 2018, 10, 298. [Google Scholar] [CrossRef] [PubMed]
- Blomstrand, E. A Role for Branched-Chain Amino Acids in Reducing Central Fatigue. J. Nutr. 2006, 136, 544S–547S. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, L.; Rabelo, P.; Moraes, M.; Teixeira-Coelho, F.; Coimbra, C.; Wanner, S.; Soares, D. Physical exercise-induced fatigue: The role of serotonergic and dopaminergic systems. Braz. J. Med. Biol. Res. 2017, 50, e6432. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M. Dosing and Efficacy of Glutamine Supplementation in Human Exercise and Sport Training. J. Nutr. 2008, 138, 2045S–2049S. [Google Scholar] [CrossRef]
- Reid, M.B.; Allen, D.G.; Whitehead, N.P.; Froehner, S.C.; Ji, L.L.; Zuo, L.; Pannell, B.K.; Re, A.T.; Best, T.M.; Wagner, P.D.; et al. Invited Review: Redox modulation of skeletal muscle contraction: What we know and what we don’t. J. Appl. Physiol. 2001, 90, 724–731. [Google Scholar] [CrossRef]
- Finaud, J.; Lac, G.; Filaire, E. Oxidative stress: Relationship with exercise and training. Sports Med. 2006, 36, 327–358. [Google Scholar] [CrossRef] [PubMed]
- Daniela, M.; Catalina, L.; Ilie, O.; Paula, M.; Daniel-Andrei, I.; Ioana, B. Effects of Exercise Training on the Autonomic Nervous System with a Focus on Anti-Inflammatory and Antioxidants Effects. Antioxidants 2022, 11, 350. [Google Scholar] [CrossRef] [PubMed]
- Bobbert, T.; Brechtel, L.; Mai, K.; Otto, B.; Maser-Gluth, C.; Pfeiffer, A.F.H.; Spranger, J.; Diederich, S. Adaptation of the hypothalamic-pituitary hormones during intensive endurance training. Clin. Endocrinol. 2005, 63, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Tominaga, T.; Ruhee, R.T.; Ma, S. Characterization and Modulation of Systemic Inflammatory Response to Exhaustive Exercise in Relation to Oxidative Stress. Antioxidants 2020, 9, 401. [Google Scholar] [CrossRef] [PubMed]
- Małkowska, P.; Sawczuk, M. Cytokines as Biomarkers for Evaluating Physical Exercise in Trained and Non-Trained Individuals: A Narrative Review. Int. J. Mol. Sci. 2023, 24, 11156. [Google Scholar] [CrossRef] [PubMed]
- La Torre, M.E.; Monda, A.; Messina, A.; de Stefano, M.I.; Monda, V.; Moscatelli, F.; Tafuri, F.; Saraiello, E.; Latino, F.; Monda, M.; et al. The Potential Role of Nutrition in Overtraining Syndrome: A Narrative Review. Nutrients 2023, 15, 4916. [Google Scholar] [CrossRef] [PubMed]
- Kreher, J.B. Diagnosis and prevention of overtraining syndrome: An opinion on education strategies. Open Access J. Sports Med. 2016, 7, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Coco, M.; Buscemi, A.; Ramaci, T.; Tusak, M.; Di Corrado, D.; Perciavalle, V.; Maugeri, G.; Perciavalle, V.; Musumeci, G. Influences of Blood Lactate Levels on Cognitive Domains and Physical Health during a Sports Stress. Brief Review. Int. J. Environ. Res. Public Health 2020, 17, 9043. [Google Scholar] [CrossRef] [PubMed]
- Haller, N.; Behringer, M.; Reichel, T.; Wahl, P.; Simon, P.; Krüger, K.; Zimmer, P.; Stöggl, T. Blood-Based Biomarkers for Managing Workload in Athletes: Considerations and Recommendations for Evidence-Based Use of Established Biomarkers. Sports Med. 2023, 53, 1315–1333. [Google Scholar] [CrossRef]
- Brancaccio, P.; Maffulli, N.; Limongelli, F.M. Creatine kinase monitoring in sport medicine. Br. Med. Bull. 2007, 81–82, 209–230. [Google Scholar] [CrossRef]
- Haller, N.; Reichel, T.; Zimmer, P.; Behringer, M.; Wahl, P.; Stöggl, T.; Krüger, K.; Simon, P. Blood-Based Biomarkers for Managing Workload in Athletes: Perspectives for Research on Emerging Biomarkers. Sports Med. 2023, 53, 2039–2053. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Hsiao, Y.T.; Huang, W.C. Physiological and Psychological Effects of Treadmill Overtraining Implementation. Biology 2021, 10, 515. [Google Scholar] [CrossRef] [PubMed]
- Trochimiak, T.; Hübner-Woźniak, E. Effect of exercise on the level of immunoglobulin a in saliva. Biol. Sport 2012, 29, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Petrov, L.; Alexandrova, A.; Kachaunov, M.; Penov, R.; Sheytanova, T.; Kolimechkov, S. Effect of glutathione supplementation on swimmers’ performance. Pedagog. Phys. Cult. Sports 2021, 25, 215–224. [Google Scholar] [CrossRef]
- Halson, S.L.; Bridge, M.W.; Meeusen, R.; Busschaert, B.; Gleeson, M.; Jones, D.A.; Jeukendrup, A.E. Time course of performance changes and fatigue markers during intensified training in trained cyclists. J. Appl. Physiol. 2002, 93, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, E.; Tagesen, E.C.; Hart, T.L.; Miller, B.A.; Jajtner, A.R. Comparison of the lymphocyte response to interval exercise versus continuous exercise in recreationally trained men. Brain Behav. Immun. Health 2022, 20, 100415. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.; Hunt, J.; Piasecki, J.; Hough, J. Lymphocyte and dendritic cell response to a period of intensified training in young healthy humans and rodents: A systematic review and meta-analysis. Front. Physiol. 2022, 13, 998925. [Google Scholar] [CrossRef]
- Baker, C.; Piasecki, J.; Hunt, J.A.; Hough, J. The reproducibility of dendritic cell and T cell counts to a 30-min high-intensity cycling protocol as a tool to highlight overtraining. Exp. Physiol. 2024, 109, 380–392. [Google Scholar] [CrossRef]
- Petibois, C.; Cazorla, G.; Poortmans, J.R.; Déléris, G. Biochemical aspects of overtraining in endurance sports: The metabolism alteration process syndrome. Sports Med. 2003, 33, 83–94. [Google Scholar] [CrossRef]
- Petibois, C.; Cazorla, G.; Poortmans, J.R.; Déléris, G. Biochemical aspects of overtraining in endurance sports: A review. Sports Med. 2002, 32, 867–878. [Google Scholar] [CrossRef]
- Urhausen, A.; Kindermann, W. Diagnosis of overtraining: What tools do we have? Sports Med. 2002, 32, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Cadegiani, F.A.; Kater, C.E. Hormonal aspects of overtraining syndrome: A systematic review. BMC Sports Sci. Med. Rehabil. 2017, 9, 14. [Google Scholar] [CrossRef]
- Graf, C.; Ferrari, N. Metabolic Health—The Role of Adipo-Myokines. Int. J. Mol. Sci. 2019, 20, 6159. [Google Scholar] [CrossRef]
- Coelho, M.; Oliveira, T.; Fernandes, R. Biochemistry of adipose tissue: An endocrine organ. Arch. Med. Sci. 2013, 9, 191–200. [Google Scholar] [CrossRef]
- Jung, S.M.; Sanchez-Gurmaches, J.; Guertin, D.A. Brown Adipose Tissue Development and Metabolism. In Brown Adipose Tissue; Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2019; Volume 251, pp. 3–36. [Google Scholar]
- Sakers, A.; De Siqueira, M.K.; Seale, P.; Villanueva, C.J. Adipose-tissue plasticity in health and disease. Cell 2022, 185, 419–446. [Google Scholar] [CrossRef]
- Colleluori, G.; Perugini, J.; Di Vincenzo, A.; Senzacqua, M.; Giordano, A.; Cinti, S. Brown Fat Anatomy in Humans and Rodents. Methods Mol. Biol. 2022, 2448, 19–42. [Google Scholar] [PubMed]
- Chait, A.; den Hartigh, L.J. Adipose Tissue Distribution Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Alfieri, A.; D’Angelo, S.; Mazzeo, F. Role of Nutritional Supplements in Sport, Exercise and Health. Nutrients 2023, 15, 4429. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, N.; Walden, T.B.; Shabalina, I.G.; Timmons, J.A.; Cannon, B.; Nedergaard, J. Chronic peroxisome proliferator-activated receptor gamma (PPARgamma) activation of epididymally derived white adipocyte cultures reveals a population of thermogenically competent, UCP1-containing adipocytes molecularly distinct from classic brown adipocytes. J. Biol. Chem. 2010, 285, 7153–7164. [Google Scholar]
- Seale, P.; Bjork, B.; Yang, W.; Kajimura, S.; Chin, S.; Kuang, S.; Scimè, A.; Devarakonda, S.; Conroe, H.M.; Erdjument-Bromage, H.; et al. PRDM16 controls a brown fat/skeletal muscle switch. Nature 2008, 454, 961–967. [Google Scholar] [CrossRef]
- Kirichenko, T.V.; Markina, Y.V.; Bogatyreva, A.I.; Tolstik, T.V.; Varaeva, Y.R.; Starodubova, A.V. The Role of Adipokines in Inflammatory Mechanisms of Obesity. Int. J. Mol. Sci. 2022, 23, 14982. [Google Scholar] [CrossRef] [PubMed]
- Neto, J.C.R.; Lira, F.S.; Oyama, L.M.; Zanchi, N.E.; Yamashita, A.S.; Batista, M.L., Jr.; Oller do Nascimento, C.M.; Seelaender, M. Exhaustive exercise causes an anti-inflammatory effect in skeletal muscle and a pro-inflammatory effect in adipose tissue in rats. Eur. J. Appl. Physiol. 2009, 106, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Qi, Z.; Ding, S. Exercise-Induced Adipose Tissue Thermogenesis and Browning: How to Explain the Conflicting Findings? Int. J. Mol. Sci. 2022, 23, 13142. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.; Karpe, F.; Lafontan, M.; Frayn, K. Physical activity and exercise in the regulation of human adipose tissue physiology. Physiol. Rev. 2012, 92, 157–191. [Google Scholar] [CrossRef] [PubMed]
- Anthony, S.R.; Guarnieri, A.; Lanzillotta, L.; Gozdiff, A.; Green, L.C.; O’Grady, K.; Helsley, R.N.; Owens Iii, A.P.; Tranter, M. HuR expression in adipose tissue mediates energy expenditure and acute thermogenesis independent of UCP1 expression. Adipocyte 2020, 9, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Bal, N.C.; Singh, S.; Reis, F.C.G.; Maurya, S.K.; Pani, S.; Rowland, L.A.; Periasamy, M. Both brown adipose tissue and skeletal muscle thermogenesis processes are activated during mild to severe cold adaptation in mice. J. Biol. Chem. 2017, 292, 16616–16625. [Google Scholar] [CrossRef] [PubMed]
- Grygiel-Górniak, B. Peroxisome proliferator-activated receptors and their ligands: Nutritional and clinical implications—A review. Nutr. J. 2014, 13, 17. [Google Scholar] [CrossRef]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1α-dependent myokine that drives browning of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Fisher, F.M.; Kleiner, S.; Douris, N.; Fox, E.C.; Mepani, R.J.; Verdeguer, F.; Wu, J.; Kharitonenkov, A.; Flier, J.S.; Maratos-Flier, E.; et al. FGF21 regulates PGC-1α and browning of white adipose tissues in adaptive thermogenesis. Genes Dev. 2012, 26, 271–281. [Google Scholar] [CrossRef]
- Lira, F.S.; Rosa, J.C.; Yamashita, A.S.; Koyama, C.H.; Batista, M.L., Jr.; Seelaender, M. Endurance training induces depot-specific changes in IL-10/TNF-alpha ratio in rat adipose tissue. Cytokine 2009, 45, 80–85. [Google Scholar] [CrossRef]
- Scheffer, D.D.L.; Latini, A. Exercise-induced immune system response: Anti-inflammatory status on peripheral and central organs. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165823. [Google Scholar] [CrossRef] [PubMed]
- Main, L.C.; Dawson, B.; Grove, J.R.; Landers, G.J.; Goodman, C. Impact of training on changes in perceived stress and cytokine production. Res. Sports Med. 2009, 17, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef] [PubMed]
- Marlatt, K.L.; Ravussin, E. Brown Adipose Tissue: An Update on Recent Findings. Curr. Obes. Rep. 2017, 6, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Stanford, K.I.; Lynes, M.D.; Takahashi, H.; Baer, L.A.; Arts, P.J.; May, F.J.; Lehnig, A.C.; Middelbeek, R.J.W.; Richard, J.J.; So, K.; et al. 12,13-diHOME: An Exercise-Induced Lipokine that Increases Skeletal Muscle Fatty Acid Uptake. Cell Metab. 2018, 27, 1111–1120.e3. [Google Scholar] [CrossRef] [PubMed]
- de Las Heras, N.; Klett-Mingo, M.; Ballesteros, S.; Martín-Fernández, B.; Escribano, Ó.; Blanco-Rivero, J.; Balfagón, G.; Hribal, M.L.; Benito, M.; Lahera, V.; et al. Chronic Exercise Improves Mitochondrial Function and Insulin Sensitivity in Brown Adipose Tissue. Front. Physiol. 2018, 9, 1122. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Delgado, G.; Martinez-Tellez, B.; Gil, A.; Ruiz, J.R. Is Brown Adipose Tissue-Mediated Adaptive Thermogenesis the Missing Component of the Constrained Total Energy Expenditure Model? Ann. Nutr. Metab. 2016, 69, 51–53. [Google Scholar] [CrossRef] [PubMed]
- Muise, E.S.; Azzolina, B.; Kuo, D.W.; El-Sherbeini, M.; Tan, Y.; Yuan, X.; Mu, J.; Thompson, J.R.; Berger, J.P.; Wong, K.K. Adipose fibroblast growth factor 21 is up-regulated by peroxisome proliferator-activated receptor gamma and altered metabolic states. Mol. Pharmacol. 2008, 74, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Hondares, E.; Iglesias, R.; Giralt, A.; Gonzalez, F.J.; Giralt, M.; Mampel, T.; Villarroya, F. Thermogenic activation induces FGF21 expression and release in brown adipose tissue. J. Biol. Chem. 2011, 286, 12983–12990. [Google Scholar] [CrossRef]
- Kim, K.H.; Kim, S.H.; Min, Y.K.; Yang, H.M.; Lee, J.B.; Lee, M.S. Acute exercise induces FGF21 expression in mice and in healthy humans. PLoS ONE 2013, 8, e63517. [Google Scholar] [CrossRef]
- Cuevas-Ramos, D.; Almeda-Valdés, P.; Meza-Arana, C.E.; Brito-Córdova, G.; Gómez-Pérez, F.J.; Mehta, R.; Oseguera-Moguel, J.; Aguilar-Salinas, C.A. Exercise increases serum fibroblast growth factor 21 (FGF21) levels. PLoS ONE 2012, 7, e38022. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Petrovic, N.; Cao, R.; Larsson, O.; Lim, S.; Chen, S.; Feldmann, H.M.; Liang, Z.; Zhu, Z.; Nedergaard, J.; et al. Hypoxia-independent angiogenesis in adipose tissues during cold acclimation. Cell Metab. 2009, 9, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Nurnazahiah, A.; Shahril, M.R.; Syamimi, Z.N.; Aryati, A.; Sulaiman, S.; Lua, P.L. Relationship of objectively measured physical activity and sedentary behaviour with health-related quality of life among breast cancer survivors. Health Qual. Life Outcomes 2020, 18, 222. [Google Scholar] [CrossRef] [PubMed]
- Nigro, E.; Scudiero, O.; Monaco, M.L.; Palmieri, A.; Mazzarella, G.; Costagliola, C.; Bianco, A.; Daniele, A. New insight into adiponectin role in obesity and obesity-related diseases. BioMed Res. Int. 2014, 2014, 658913. [Google Scholar] [CrossRef] [PubMed]
- Fantuzzi, G. Adiponectin and inflammation: Consensus and controversy. J. Allergy Clin. Immunol. 2008, 121, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Anna, V.; Goropashnaya, J.H.; Sexton, M.; Havel, P.J.; Stanhope, K.L.; Plaetke, R.; Mohatt, G.V.; Boyera, B.B. Relationships Between Plasma Adiponectin and Body Fat Distribution, Insulin Sensitivity, and Plasma Lipoproteins in Alaskan Yup’ik Eskimos: The CANHR Study. Metabolism 2009, 58, 22–29. [Google Scholar] [CrossRef]
- Khoramipour, K.; Chamari, K.; Hekmatikar, A.A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.M.; Bragazzi, N.L. Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition. Nutrients 2021, 13, 1180. [Google Scholar] [CrossRef] [PubMed]
- Mallardo, M.; D’Alleva, M.; Lazzer, S.; Giovanelli, N.; Graniero, F.; Billat, V.; Fiori, F.; Marinoni, M.; Parpinel, M.; Daniele, A.; et al. Improvement of adiponectin in relation to physical performance and body composition in young obese males subjected to twenty-four weeks of training programs. Heliyon 2023, 9, e15790. [Google Scholar] [CrossRef] [PubMed]
- Sirico, F.; Bianco, A.; D’Alicandro, G.; Castaldo, C.; Montagnani, S.; Spera, R.; Di Meglio, F.; Nurzynska, D. Effects of Physical Exercise on Adiponectin, Leptin, and Inflammatory Markers in Childhood Obesity: Systematic Review and Meta-Analysis. Child. Obes. 2018, 14, 207–217. [Google Scholar] [CrossRef]
- Zaidi, H.; Byrkjeland, R.; Njerve, I.U.; Åkra, S.; Solheim, S.; Arnesen, H.; Seljeflot, I.; Opstad, T.B. Adiponectin in relation to exercise and physical performance in patients with type 2 diabetes and coronary artery disease. Adipocyte 2021, 10, 612–620. [Google Scholar] [CrossRef]
- Lakka, T.A.; Laaksonen, D.E. Physical activity in prevention and treatment of the metabolic syndrome. Appl. Physiol. Nutr. Metab. 2007, 32, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Jurimae, J.; Maestu, J.; Jurimae, T.; Mangus, B.; von Duvillard, S.P. Peripheral signals of energy homeostasis as possible markers of training stress in athletes: A review. Metabolism 2011, 60, 335–350. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.A.; Fiatarone Singh, M.A. Effects of exercise on adiponectin: A systematic review. Obes. Silver Spring 2008, 16, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Ihalainen, J.K.; Peltonen, H.; Paulsen, G.; Ahtiainen, J.P.; Taipale, R.S.; Hämäläinen, M.; Moilanen, E.; Mero, A.A. Inflammation status of healthy young men: Initial and specific responses to resistance training. Appl. Physiol. Nutr. Metab. 2018, 43, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Jurimae, J.; Purge, P.; Jurimae, T. Effect of prolonged training period on plasma adiponectin in elite male rowers. Horm. Metab. Res. 2007, 39, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Jurimae, J.; Purge, P.; Jurimae, T. Adiponectin and stress hormone responses to maximal sculling after volume-extended training season in elite rowers. Metabolism 2006, 55, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Bobbert, T.; Wegewitz, U.; Brechtel, L.; Freudenberg, M.; Mai, K.; Mi∂hlig, M.; Diederich, S.; Ristow, M.; Rochlitz, H.; Pfeiffer, A.F.H.; et al. Adiponectin oligomers in human serum during acute and chronic exercise: Relation to lipid metabolism and insulin sensitivity. Int. J. Sports Med. 2007, 28, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Macaluso, A.; De Vito, G. Muscle strength, power and adaptations to resistance training in older people. Eur. J. Appl. Physiol. 2004, 91, 450–472. [Google Scholar] [CrossRef] [PubMed]
- Bouassida, A.; Chamari, K.; Zaouali, M.; Feki, Y.; Zbidi, A.; Tabka, Z. Review on leptin and adiponectin responses and adaptations to acute and chronic exercise. Br. J. Sports Med. 2010, 44, 620–630. [Google Scholar] [CrossRef]
- Nigro, E.; Sangiorgio, D.; Scudiero, O.; Monaco, M.L.; Polito, R.; Villone, G.; Daniele, A. Gene molecular analysis and Adiponectin expression in professional Water Polo players. Cytokine 2016, 81, 88–93. [Google Scholar] [CrossRef]
- Jurimae, J.; Vaiksaar, S.; Muestu, J.; Purge, P.; Jurimae, T. Adiponectin and bone metabolism markers in female rowers: Eumenorrheic and oral contraceptive users. J. Endocrinol. Investig. 2011, 34, 835–839. [Google Scholar] [CrossRef]
- Kraemer, R.R.; Durand, R.J.; Acevedo, E.O.; Johnson, L.G.; Kraemer, G.R.; Hebert, E.P.; Castracane, V.D. Rigorous running increases growth hormone and insulin-like growth factor-I without altering ghrelin. Exp. Biol. Med. 2004, 229, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Jamurtas, A.Z.; Theocharis, V.; Koukoulis, G.; Stakias, N.; Fatouros, I.G.; Kouretas, D.; Koutedakis, Y. The effects of acute exercise on serum adiponectin and resistin levels and their relation to insulin sensitivity in overweight males. Eur. J. Appl. Physiol. 2006, 97, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, M.A.; White, L.J.; McCoy, S.; Kim, H.W.; Petty, T.; Wilsey, J. Plasma adiponectin response to acute exercise in healthy subjects. Eur. J. Appl. Physiol. 2004, 91, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Fallo, F.; Scarda, A.; Sonino, N.; Paoletta, A.; Boscaro, M.; Pagano, C.; Federspil, G.; Vettor, R. Effect of glucocorticoids on adiponectin: A study in healthy subjects and in Cushing’s syndrome. Eur. J. Endocrinol. 2004, 150, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Guo, M.; Zhang, D.; Cheng, S.-Y.; Liu, M.; Ding, J.; Scherer, P.E.; Liu, F.; Lu, X.-Y. Adiponectin is critical in determining susceptibility to depressive behaviors and has antidepressant-like activity. Proc. Natl. Acad. Sci. USA 2012, 109, 12248–12253. [Google Scholar] [CrossRef] [PubMed]
- La Cava, A. Leptin in inflammation and autoimmunity. Cytokine 2017, 98, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Sööt, T.; Jürimäe, T.; Jürimäe, J. Areal bone density in young females with different physical activity patterns: Relationships with plasma leptin and body composition. J. Sports Med. Phys. Fit. 2007, 47, 65–69. [Google Scholar]
- Jürimäe, J.; Jürimäe, T. Leptin responses to short term exercise in college level male rowers. Br. J. Sports Med. 2005, 39, 6–9. [Google Scholar] [CrossRef]
- de Oliveira, D.C.X.; Procida, I.R.; das Neves Borges-Silva, C. Effect of training judo in the competition period on the plasmatic levels of leptin and pro-inflammatory cytokines in high-performance male athletes. Biol. Trace Elem. Res. 2010, 135, 345–354. [Google Scholar] [CrossRef]
- Perseghin, G.; Burska, A.; Lattuada, G.; Alberti, G.; Costantino, F.; Ragogna, F.; Oggionni, S.; Scollo, A.; Terruzzi, I.; Luzi, L. Increased serum resistin in elite endurance athletes with high insulin sensitivity. Diabetologia 2006, 49, 1893–1900. [Google Scholar] [CrossRef] [PubMed]
- Guadalupe-Grau, A.; Perez-Gomez, J.; Olmedillas, H.; Chavarren, J.; Dorado, C.; Santana, A.; Serrano-Sanchez, J.A.; Calbet, J.A.L. Strength training combined with plyometric jumps in adults: Sex differences in fat-bone axis adaptations. J. Appl. Physiol. 2009, 106, 1100–1111. [Google Scholar] [CrossRef] [PubMed]
- Varady, K.A.; Bhutani, S.; Church, E.C.; Phillips, S.A. responses to acute resistance exercise in trained and untrained men. Med. Sci. Sports Exerc. 2010, 42, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Jürimäe, J.; Mäestu, J.; Jürimäe, T. Leptin as a marker of training stress in highly trained male rowers? Eur. J. Appl. Physiol. 2003, 90, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Joro, R.; Uusitalo, A.; DeRuisseau, K.C.; Atalay, M. Changes in cytokines, leptin, and IGF-1 levels in overtrained athletes during a prolonged recovery phase: A case-control study. J. Sports Sci. 2017, 35, 2342–2349. [Google Scholar] [CrossRef] [PubMed]
- Yamaner, F. The effect of overtraining on serum leptin levels in women national wrestlers. Pedagog. Psychol. Med. Biol. Probl. Phys. Train. Sports 2019, 23, 209–213. [Google Scholar] [CrossRef]
- Liu, Z.G. Molecular mechanism of TNF signaling and beyond. Cell Res. 2005, 15, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Cawthorn, W.P.; Sethi, J.K. TNF-alpha and adipocyte biology. FEBS Lett. 2008, 582, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.M.W.; Pedersen, B.K. The anti-inflammatory effect of Exercise. J. Appl. Physiol. 2005, 98, 1154–1162. [Google Scholar] [CrossRef]
- do Santiago, L.M.A.; Lima Neto, L.G.; Pereira, G.B.; Leite, R.D.; Texeira Mostarda, C.; de Oliveira Brito Monzani, J.; Rodrigues Sousa, W.; Rodrigues Pinheiro, A.J.M.; Navarro, F. Effects of Resistance Training on Immunoinflammatory Response, TNF-Alpha Gene Expression, and Body Composition in Elderly Women. J. Aging Res. 2018, 2018, 1467025. [Google Scholar] [CrossRef]
- Ostrowski, K.; Rohde, T.; Asp, S.; Schjerling, P.; Pedersen, B.K. Pro- and anti-inflammatory cytokine balance in strenuous exercise in humans. J. Physiol. 1999, 515 Pt 1, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Starkie, R.L.; Rolland, J.; Angus, D.J.; Anderson, M.J.; Febbraio, M.A. Circulating monocytes are not the source of elevations in plasma IL-6 and TNF-alpha levels after prolonged running. Am. J. Physiol. Cell Physiol. 2001, 280, C769–C774. [Google Scholar] [CrossRef]
- da Rocha, A.L.; Pinto, A.P.; Kohama, E.B.; Pauli, J.R.; de Moura, L.P.; Cintra, D.E.; Ropelle, E.R.; da Silva, A.S.R. The proinflammatory effects of chronic excessive exercise. Cytokine 2019, 119, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Nara, M.; Kanda, T.; Tsukui, S.; Inukai, T.; Shimomura, Y.; Inoue, S.; Kobayashi, I. Running exercise increases tumor necrosis factor-alpha secreting from mesenteric fat in insulin-resistant rats. Life Sci. 1999, 65, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Deb, A.; Deshmukh, B.; Ramteke, P.; Bhati, F.K.; Bhat, M.K. Resistin: A journey from metabolism to cancer. Transl. Oncol. 2021, 14, 101178. [Google Scholar] [CrossRef] [PubMed]
- Askin, L.; Abus, S.; Tanriverdi, O. Resistin and Cardiovascular Disease: A Review of the Current Literature Regarding Clinical and Pathological Relationships. Curr. Cardiol. Rev. 2022, 18, e290721195114. [Google Scholar] [CrossRef] [PubMed]
- Acquarone, E.; Monacelli, F.; Borghi, R.; Nencioni, A.; Odetti, P. Resistin: A reappraisal. Mech. Ageing Dev. 2019, 178, 46–63. [Google Scholar] [CrossRef] [PubMed]
- Prestes, J.; Shiguemoto, G.; Botero, J.P.; Frollini, A.; Dias, R.; Leite, R.; Pereira, G.; Magosso, R.; Baldissera, V.; Cavaglieri, C.; et al. Effects of resistance training on resistin, leptin, cytokines, and muscle force in elderly post-menopausal women. J. Sports Sci. 2009, 27, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Roupas, N.D.; Mamali, I.; Maragkos, S.; Leonidou, L.; Armeni, A.K.; Markantes, G.K.; Tsekouras, A.; Sakellaropoulos, G.C.; Markou, K.B.; Georgopoulos, N.A. The effect of prolonged aerobic exercise on serum adipokine levels during an ultra-marathon endurance race. Hormones 2013, 12, 275–282. [Google Scholar] [CrossRef]
- Vuolteenaho, K.; Leppinen, T.; Kekkonen, R.; Korpela, R.; Moilanen, E. Running a marathon induces changes in adipokine levels and in markers of cartilage degradation--novel role for resistin. PLoS ONE 2014, 9, e110481. [Google Scholar] [CrossRef]
- Sansoni, V.; Vernillo, G.; Perego, S.; Barbuti, A.; Merati, G.; Schena, F.; La Torre, A.; Banfi, G.; Lombardi, G. Bone turnover response is linked to both acute and established metabolic changes in ultra-marathon runners. Endocrine 2017, 56, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Czajkowska, A.; Ambroszkiewicz, J.; Mróz, A.; Witek, K.; Nowicki, D.; Małek, Ł. The Effect of the Ultra-Marathon Run at a Distance of 100 Kilometers on the Concentration of Selected Adipokines in Adult Men. Int. J. Environ. Res. Public Health 2020, 17, 4289. [Google Scholar] [CrossRef] [PubMed]
- Bjersing, J.L.; Erlandsson, M.; Bokarewa, M.I.; Mannerkorpi, K. Exercise and obesity in fibromyalgia: Beneficial roles of IGF-1 and resistin? Arthritis Res. Ther. 2013, 15, R34. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, P.; Silvestre, R.A.; Díez, J.J. Growth differentiation factor 15 (GDF-15) in endocrinology. Endocrine 2023, 81, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Keipert, S.; Ost, M. Stress-induced FGF21 and GDF15 in obesity and obesity resistance. Trends Endocrinol. Metab. 2021, 32, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Barma, M.; Khan, F.; Price, R.J.G.; Donnan, P.T.; Messow, C.M.; Ford, I.; McConnachie, A.; Struthers, A.D.; McMurdo, M.E.T.; Witham, M.D. Association between GDF-15 levels and changes in vascular and physical function in older patients with hypertension. Aging Clin. Exp. Res. 2017, 29, 1055–1059. [Google Scholar] [CrossRef] [PubMed]
- Semba, R.D.; Gonzalez-Freire, M.; Tanaka, T.; Biancotto, A.; Zhang, P.; Shardell, M.; Moaddel, R.; CHI Consortium; Ferrucci, L. Elevated Plasma Growth and Differentiation Factor 15 Is Associated with Slower Gait Speed and Lower Physical Performance in Healthy Community-Dwelling Adults. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Rochette, L.; Zeller, M.; Cottin, Y.; Vergely, C. Insights into Mechanisms of GDF15 and Receptor GFRAL: Therapeutic Targets. Trends Endocrinol. Metab. 2020, 31, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Tavenier, J.; Jee, L.; Rasmussen, H.; Andersen, A.L.; Houlind, M.B.; Langkilde, A.; Andersen, O.; Petersen, J.; Nehlin, J.O. Association of GDF15 with Inflammation and Physical Function during Aging and Recovery After Acute Hospitalization: A Longitudinal Study of Older Patients and Age-Matched Controls. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 964–974. [Google Scholar] [CrossRef]
- Tchou, I.; Margeli, A.; Tsironi, M.; Skenderi, K.; Barnet, M.; Kanaka-Gantenbein, C.; Papassotiriou, I.; Beris, P. Growth-differentiation factor-15, endoglin and N-terminal pro-brain natriuretic peptide induction in athletes participating in an ultramarathon foot race. Biomarkers 2009, 14, 418–422. [Google Scholar] [CrossRef]
- Galliera, E.; Lombardi, G.; Marazzi, M.G.; Grasso, D.; Vianello, E.; Pozzoni, R.; Banfi, G.; Corsi Romanelli, M.M. Acute exercise in elite rugby players increases the circulating level of the cardiovascular biomarker GDF-15. Scand. J. Clin. Lab. Investig. 2014, 74, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Igual Gil, C.; Ost, M.; Kasch, J.; Schumann, S.; Heider, S.; Klaus, S. Role of GDF15 in active lifestyle induced metabolic adaptations and acute exercise response in mice. Sci. Rep. 2019, 9, 20120. [Google Scholar] [CrossRef] [PubMed]
- Kleinert, M.; Clemmensen, C.; Sjøberg, K.A.; Strini, C.C.; Fuglsbjerg Jeppesen, J.; Wojtaszewski, J.F.P.; Kiens, B.; Richter, E.A. Exercise increases circulating GDF15 in humans. Mol. Metab. 2018, 9, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Kim-Muller, J.Y.; Song, L.; LaCarubba, P.B.; Pashos, E.; Li, X.; Rinaldi, A.; Joaquim, S.; Stansfield, J.C.; Zhang, J.; Robertson, A.; et al. GDF15 neutralization restores muscle function and physical performance in a mouse model of cancer cachexia. Cell Rep. 2023, 42, 111947. [Google Scholar] [CrossRef] [PubMed]
- Merchant, R.A.; Chan, Y.H.; Duque, G. GDF-15 Is Associated with Poor Physical Function in Prefrail Older Adults with Diabetes. J. Diabetes Res. 2023, 2023, 2519128. [Google Scholar] [CrossRef] [PubMed]
- Jena, J.; García-Peña, L.M.; Weatherford, E.T.; Marti, A.; Bjorkman, S.H.; Kato, K.; Koneru, J.; Chen, J.H.; Seeley, R.J.; Abel, E.D.; et al. GDF15 is required for cold-induced thermogenesis and contributes to improved systemic metabolic health following loss of OPA1 in brown adipocytes. eLife 2023, 12, e86452. [Google Scholar] [CrossRef] [PubMed]
- Renqing, Z. Irisin at the crossroads of inter-organ communications: Challenge and implications. Front. Endocrinol. 2022, 13, 989135. [Google Scholar] [CrossRef]
- Arhire, L.I.; Mihalache, L.; Covasa, M. Irisin: A Hope in Understanding and Managing Obesity and Metabolic Syndrome. Front. Endocrinol. 2019, 10, 524. [Google Scholar] [CrossRef] [PubMed]
- Kazeminasab, F.; Sadeghi, E.; Afshari-Safavi, A. Comparative Impact of Various Exercises on Circulating Irisin in Healthy Subjects: A Systematic Review and Network Meta-Analysis. Oxid. Med. Cell Longev. 2022, 2022, 8235809. [Google Scholar] [CrossRef]
- Roggio, F.; Petrigna, L.; Trovato, B.; Di Rosa, M.; Musumeci, G. The Role of Lubricin, Irisin and Exercise in the Prevention and Treatment of Osteoarthritis. Int. J. Mol. Sci. 2023, 24, 5126. [Google Scholar] [CrossRef]
- González, D.A.; Del Cristo Rodríguez-Pérez, M.; Fuentes Ferrer, M.; Cuevas Fernández, F.J.; Rodríguez, I.M.A.; de León, C. Irisin, in women and men: Blood pressure, heart rate, obesity and insulin resistance. Front. Endocrinol. 2023, 14, 1193110. [Google Scholar] [CrossRef] [PubMed]
- Missaglia, S.; Tommasini, E.; Vago, P.; Pecci, C.; Galvani, C.; Silvestrini, A.; Mordente, A.; Tavian, D. Salivary and serum irisin in healthy adults before and after exercise. Eur. J. Transl. Myol. 2023, 33, 11093. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Trettel, C.; de Avila Pelozin, B.R.; Paes Barros, M.; Lacerda Bachi, A.L.; Senger Braga, P.G.; Momesso, C.M.; Furtado, G.E.; Valente, P.A.; Menezes Oliveira, E.; Hogervorst, E.; et al. Irisin: An anti-inflammatory exerkine in aging and redox-mediated comorbidities. Front. Endocrinol. 2023, 14, 1106529. [Google Scholar] [CrossRef] [PubMed]
- Leoni De Sousa, R.A.; Improta-Caria, A.C.; de Freitas Souza, B.S. Exercise–Linked Irisin: Consequences on Mental and Cardiovascular Health in Type 2 Diabetes. Int. J. Mol. Sci. 2021, 22, 2199. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Navarrete, J.M.; Ortega, F.; Serrano, M.; Guerra, E.; Pardo, G.; Tinahones, R.F.W.; Fernández-Real, J.M. Irisin is expressed and produced by human muscle and adipose tissue in association with obesity and insulin resistance. J. Clin. Endocrinol. Metab. 2013, 98, E769–E778. [Google Scholar] [CrossRef] [PubMed]
- Joro, R.; Korkmaz, A.T.; Lakka, A.; Uusitalo, A.L.T.; Atalay, M. Plasma irisin and its associations with oxidative stress in athletes suffering from overtraining syndrome. Physiol. Int. 2020, 107, 513–526. [Google Scholar] [CrossRef]
- Morgan, W.P.; Brown, D.R.; Raglin, J.S.; O’Connor, P.J.; Ellickson, K.A. Psychological monitoring of overtraining and staleness. Br. J. Sports Med. 1987, 21, 107–114. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameters | Physical Exercise | Training | Overtraining |
|---|---|---|---|
| Glycogen | Glycogen levels were significantly reduced immediately after exercise in both untrained and trained conditions [45]. | Glycogen depletion during training occurs in an intensity-dependent manner [46]. | Limited literature data exist regarding the relationship between low glycogen levels and overtrained athletes. It’s worth noting that athletes with normal glycogen levels still experience OTS [18]. |
| Central fatigue | Physical exercise decreases the levels of branched-chain amino acids (BCAA), favoring the entry of tryptophan into the brain and its conversion into 5-HT. Changes in performance in relation to fatigue need to be investigated in response to different types of exercise [23]. | Training influences the mechanism of central fatigue, which may vary depending on the type of training. There is a positive correlation between increased serotonin (5-HT) synthesis in the brain and fatigue among trained athletes [47]. | Excessive levels of 5-hydroxytryptamine (5-HT) in the central nervous system (CNS) can lead to OTS [doi: 10.1249/00005768-199307000-00015] [25]. Increased concentrations of this neurotransmitter in the CNS are related to mental and physical underperformance [48]. |
| Glutamine | The response of blood glutamine varies depending on the duration of exercise. Short-term exercise leads to increased muscle release of glutamine, resulting in elevated blood concentrations. Conversely, during long-term exercises, muscle synthesis of glutamine fails to meet the body’s demand for this amino acid, leading to a decrease in its levels [49]. | Prolonged periods of intense training correlate with a reduction in plasma glutamine concentration, which has been proposed as a potential contributor to exercise-induced immune suppression and susceptibility to infections among athletes [27]. | The decrease in glutamine blood levels is closely associated with OTS. Prolonged periods of exercise can lead to temporary reductions in plasma glutamine concentrations among athletes experiencing OTS [28]. |
| Oxidative Stress | ROS are produced during exercise and play a role in modulating muscle contraction levels. However, the significant increase in ROS production during intense exercise contributes to the onset of acute muscle fatigue [50]. | Training can reduce systemic oxidative stress and enhance antioxidant defenses. However, the effects on oxidative stress depend on training load, specificity, and baseline fitness levels [51]. | Athletes undergoing OTS seem to be more vulnerable to oxidative damage due to diminished responses to exercise-induced stress [29,33]. |
| Autonomic Nervous System | Exercise leads to a lower resting heart rate by boosting parasympathetic activity and reducing sympathetic activity. It also reduces plasma catecholamines and sympathetic outflow [52]. | Training induces significant hemodynamic and autonomic changes, which are dependent on training intensity, frequency, and duration [52]. | In OTS athletes, a reduction in sympathetic activation and an increase in parasympathetic dominance may contribute to performance inhibition, fatigue and depression; reduced nocturnal urinary excretion of catecholamines are also observed [4]. |
| Hypothalamic hormones | Aerobic, endurance or continuous exercise can cause variations in hypothalamic hormone levels, depending on the intensity and duration of the activity [52]. | The hormonal adaptations differ between acute and long-term training. Specifically, short-term training tends to elevate plasma levels of glucocorticoids, whereas long-term training is associated with reduced cortisol levels [53]. | Lack of relevant data. |
| Cytokines | Regular exercise is associated with an increase of anti-inflammatory cytokines in circulation, while acute exercise may initiate inflammatory cascades depending on intensity and duration [54]. | Training induces a temporary rise in pro-inflammatory cytokines, followed by anti-inflammatory cytokine production. This is attributed to the development of chronic low-grade inflammation through regular training, which primes the immune system and diminishes the cytokine response cascade to subsequent exercise. The cytokine response is influenced by the intensity and duration of training [55]. | Injuries to the muscular, skeletal, or joint systems initiate OTS. Continuing training after an acute injury without sufficient recovery can worsen the original condition and exacerbate exercise-related injuries. These injuries prompt an increase in pro-inflammatory cytokines [56]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mallardo, M.; Daniele, A.; Musumeci, G.; Nigro, E. A Narrative Review on Adipose Tissue and Overtraining: Shedding Light on the Interplay among Adipokines, Exercise and Overtraining. Int. J. Mol. Sci. 2024, 25, 4089. https://doi.org/10.3390/ijms25074089
Mallardo M, Daniele A, Musumeci G, Nigro E. A Narrative Review on Adipose Tissue and Overtraining: Shedding Light on the Interplay among Adipokines, Exercise and Overtraining. International Journal of Molecular Sciences. 2024; 25(7):4089. https://doi.org/10.3390/ijms25074089
Chicago/Turabian StyleMallardo, Marta, Aurora Daniele, Giuseppe Musumeci, and Ersilia Nigro. 2024. "A Narrative Review on Adipose Tissue and Overtraining: Shedding Light on the Interplay among Adipokines, Exercise and Overtraining" International Journal of Molecular Sciences 25, no. 7: 4089. https://doi.org/10.3390/ijms25074089
APA StyleMallardo, M., Daniele, A., Musumeci, G., & Nigro, E. (2024). A Narrative Review on Adipose Tissue and Overtraining: Shedding Light on the Interplay among Adipokines, Exercise and Overtraining. International Journal of Molecular Sciences, 25(7), 4089. https://doi.org/10.3390/ijms25074089