

Interaction of Nipah Virus F and G with the Cellular Protein Cortactin Discovered by a Proximity Interactome Assay

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
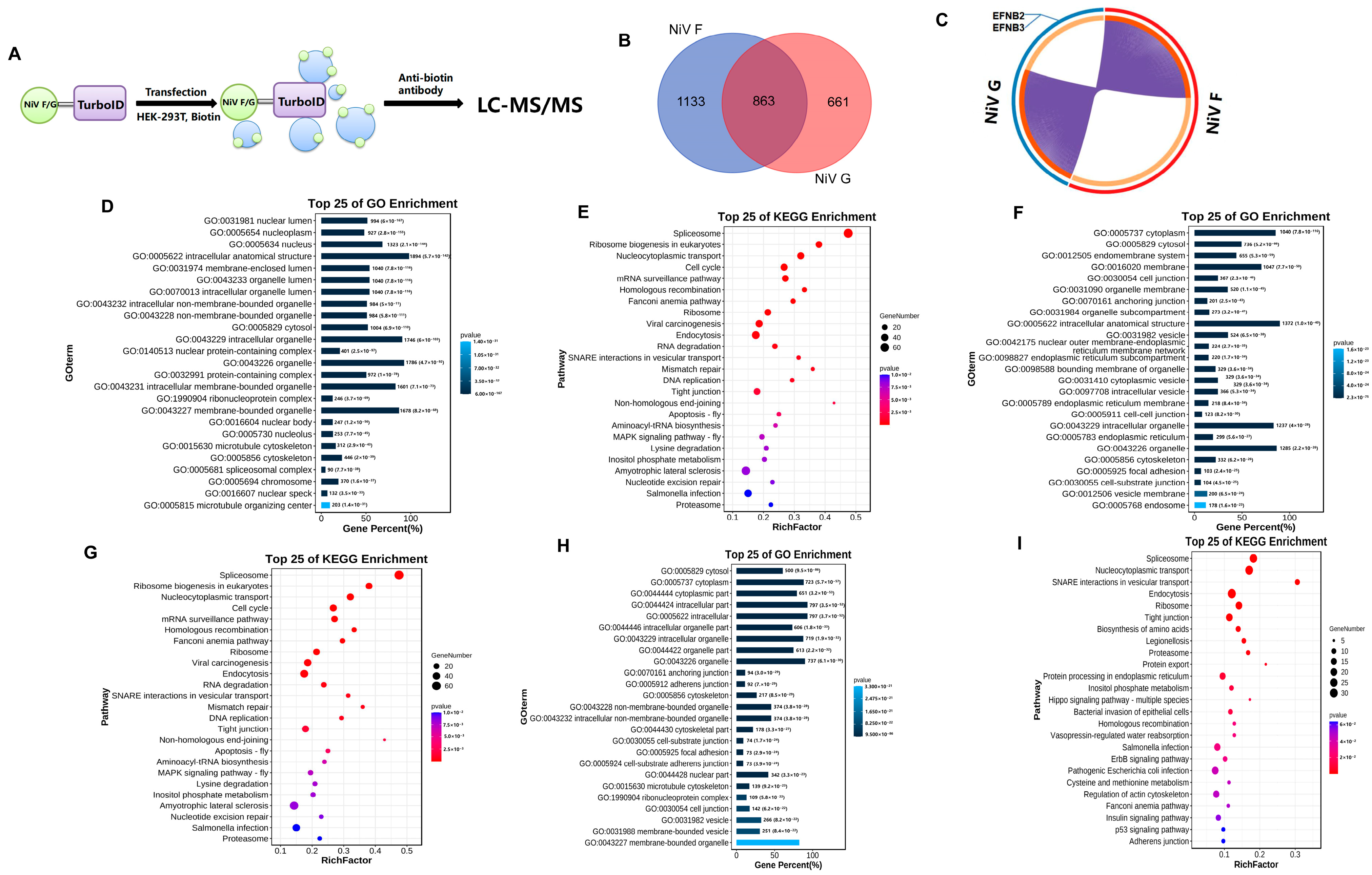
2.1. TurboID Proximity Labeling Combined with LC-MS/MS Reveals NiV F/G-Host Protein Interactomes
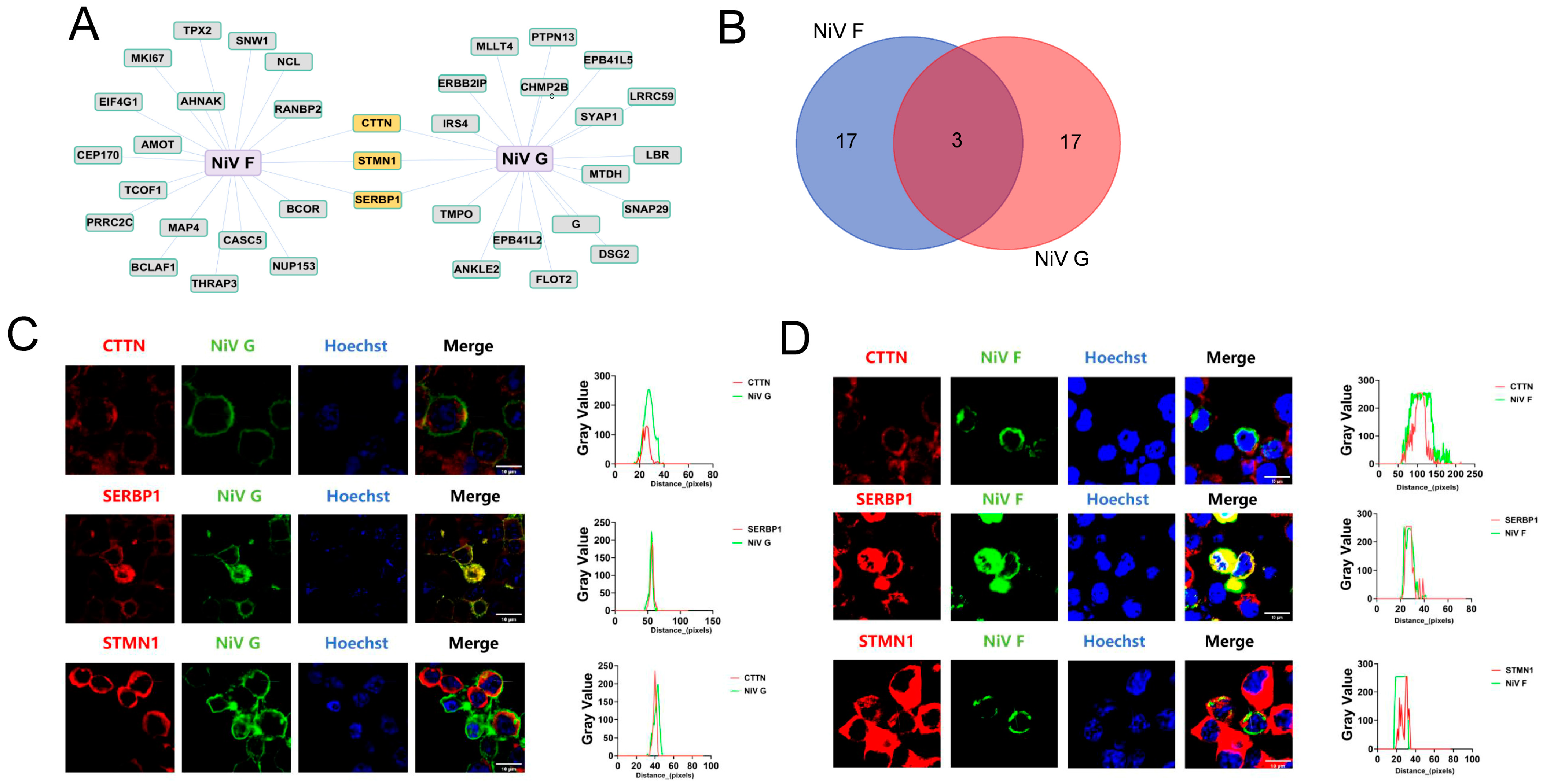
2.2. CTTN, SERBP1, and STMN1 Were Distributed on the Cell Membrane with NiV F and G
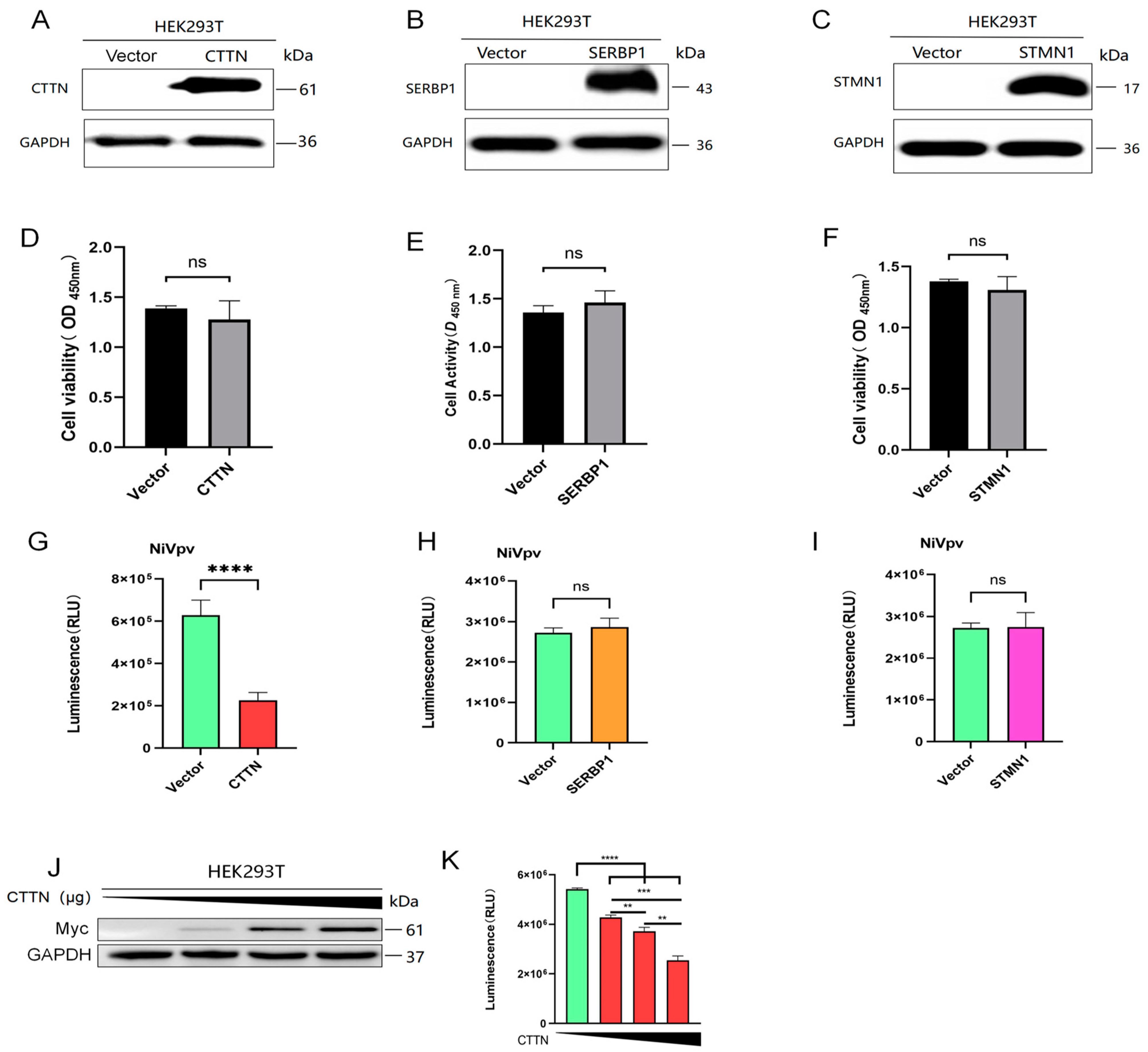
2.3. Overexpression of CTTN Restricted Infection of the NiVpv in HEK-293T Cells
2.4. Loss of Function of CTTN Promoted Infection of the NiVpv
2.5. The Infection of HeVpv Is Inhibited by CTTN
3. Discussion
4. Materials and Methods
4.1. Cells and Viruses
4.2. Plasmid Construction, siRNA, and Transfection
4.3. Cytotoxicity Assay
4.4. Pseudovirus Infection Assay
4.5. Proximity Labeling with TurboID Assays
4.6. Western Blotting
4.7. Immunolocalization and Confocal Assay
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pillai, V.S.; Krishna, G.; Veettil, M.V. Nipah Virus: Past Outbreaks and Future Containment. Viruses 2020, 12, 465. [Google Scholar] [CrossRef] [PubMed]
- Alam, A.M. Nipah virus, an emerging zoonotic disease causing fatal encephalitis. Clin. Med. (Lond. Engl.) 2022, 22, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Wagner, N.D.; Liu, H.; Rohrs, H.W.; Amarasinghe, G.K.; Gross, M.L.; Leung, D.W. Nipah Virus V Protein Binding Alters MDA5 Helicase Folding Dynamics. ACS Infect. Dis. 2022, 8, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Devnath, P.; Wajed, S.; Das, R.C.; Kar, S.; Islam, I.; Al Masud, H.M.A. The pathogenesis of Nipah virus: A review. Microb. Pathog. 2022, 170, 105693. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.J.; Gurley, E.S.; Montgomery, J.M.; Bell, M.; Carroll, D.S.; Hsu, V.P.; Formenty, P.; Croisier, A.; Bertherat, E.; Faiz, M.A.; et al. Clinical presentation of nipah virus infection in Bangladesh. Clin. Infect. Dis. 2008, 46, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Diederich, S.; Maisner, A. Molecular characteristics of the Nipah virus glycoproteins. Ann. N. Y. Acad. Sci. 2007, 1102, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Rajashankar, K.R.; Chan, Y.P.; Himanen, J.P.; Broder, C.C.; Nikolov, D.B. Host cell recognition by the henipaviruses: Crystal structures of the Nipah G attachment glycoprotein and its complex with ephrin-B3. Proc. Natl. Acad. Sci. USA 2008, 105, 9953–9958. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Stone, J.A.; Bradel-Tretheway, B.; Dabundo, J.; Benavides Montano, J.A.; Santos-Montanez, J.; Biering, S.B.; Nicola, A.V.; Iorio, R.M.; Lu, X.; et al. Unraveling a three-step spatiotemporal mechanism of triggering of receptor-induced Nipah virus fusion and cell entry. PLoS Pathog. 2013, 9, e1003770. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Bradel-Tretheway, B.; Monreal, A.I.; Saludes, J.P.; Lu, X.; Nicola, A.V.; Aguilar, H.C. Nipah virus attachment glycoprotein stalk C-terminal region links receptor binding to fusion triggering. J. Virol. 2015, 89, 1838–1850. [Google Scholar] [CrossRef]
- Chan, Y.P.; Lu, M.; Dutta, S.; Yan, L.; Barr, J.; Flora, M.; Feng, Y.R.; Xu, K.; Nikolov, D.B.; Wang, L.F.; et al. Biochemical, conformational, and immunogenic analysis of soluble trimeric forms of henipavirus fusion glycoproteins. J. Virol. 2012, 86, 11457–11471. [Google Scholar] [CrossRef]
- Branon, T.C.; Bosch, J.A.; Sanchez, A.D.; Udeshi, N.D.; Svinkina, T.; Carr, S.A.; Feldman, J.L.; Perrimon, N.; Ting, A.Y. Efficient proximity labeling in living cells and organisms with TurboID. Nat. Biotechnol. 2018, 36, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, G.; Lal, N.K.; Nagalakshmi, U.; Li, Y.; Zheng, W.; Dinesh-Kumar, S.P. TurboID-based proximity labeling reveals that UBR7 is a regulator of N NLR immune receptor-mediated immunity. Nat. Commun. 2019, 10, 3252. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.F.; Fan, S.Y.; Wang, Y.W.; Li, S.; Long, S.Y.; Gan, C.Y.; Hu, J.L. Identification of STAU1 as a regulator of HBV replication by TurboID-based proximity labeling. iScience 2022, 25, 104416. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Salemi, M.; Bhullar, R.; Guevara-Plunkett, S.; Lyu, Y.; Wang, K.-H.; Izumiya, C.; Campbell, M.; Nakajima, K.-I.; Izumiya, Y. Proximity Biotin Labeling Reveals Kaposi’s Sarcoma-Associated Herpesvirus Interferon Regulatory Factor Networks. J. Virol. 2021, 95, 02049-20. [Google Scholar] [CrossRef] [PubMed]
- Hao, P.; Qu, Q.; Pang, Z.; Li, L.; Du, S.; Shang, L.; Jin, C.; Xu, W.; Ha, Z.; Jiang, Y.; et al. Interaction of species A rotavirus VP4 with the cellular proteins vimentin and actin related protein 2 discovered by a proximity interactome assay. J. Virol. 2023, 97, e0137623. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Jiang, J.; Tang, Y.; Mei, L.; Wu, L.; Li, L.; Chen, H.; Long, F.; Xiao, J.; Peng, T. A Pseudovirus-Based Entry Assay to Evaluate Neutralizing Activity against Respiratory Syncytial Virus. Viruses 2023, 15, 1548. [Google Scholar] [CrossRef] [PubMed]
- Duprex, W.P.; Dutch, R.E. Paramyxoviruses: Pathogenesis, Vaccines, Antivirals, and Prototypes for Pandemic Preparedness. J. Infect. Dis. 2023, 228 (Suppl. S6), S390–S397. [Google Scholar] [CrossRef] [PubMed]
- Contreras, E.M.; Monreal, I.A.; Ruvalcaba, M.; Ortega, V.; Aguilar, H.C. Antivirals targeting paramyxovirus membrane fusion. Curr. Opin. Virol. 2021, 51, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Negrete, O.A.; Levroney, E.L.; Aguilar, H.C.; Bertolotti-Ciarlet, A.; Nazarian, R.; Tajyar, S.; Lee, B. EphrinB2 is the entry receptor for Nipah virus, an emergent deadly paramyxovirus. Nature 2005, 436, 401–405. [Google Scholar] [CrossRef]
- Priyadarsinee, L.; Sarma, H.; Sastry, G.N. Glycoprotein attachment with host cell surface receptor Ephrin B2 and B3 in mediating entry of nipah and hendra virus: A computational investigation. J. Chem. Sci. 2022, 134, 1–17. [Google Scholar] [CrossRef]
- Contreras, E.M.; Johnston, G.P.; Buchholz, D.W.; Ortega, V.; Monreal, I.A.; Zamora, J.L.R.; Cheung, T.; Aguilar, H.C. Roles of Cholesterol in Early and Late Steps of the Nipah Virus Membrane Fusion Cascade. J. Virol. 2021, 95. [Google Scholar] [CrossRef]
- Lo, M.K.; Spengler, J.R.; Krumpe, L.R.H.; Welch, S.R.; Chattopadhyay, A.; Harmon, J.R.; Coleman-McCray, J.D.; Scholte, F.E.M.; Hotard, A.L.; Fuqua, J.L.; et al. Griffithsin Inhibits Nipah Virus Entry and Fusion and Can Protect Syrian Golden Hamsters From Lethal Nipah Virus Challenge. J. Infect. Dis. 2020, 221 (Suppl. S4), S480–S492. [Google Scholar] [CrossRef]
- Cho, K.F.; Branon, T.C.; Udeshi, N.D.; Myers, S.A.; Carr, S.A.; Ting, A.Y. Proximity labeling in mammalian cells with TurboID and split-TurboID. Nat. Protoc. 2020, 15, 3971–3999. [Google Scholar] [CrossRef]
- Muto, A.; Sugihara, Y.; Shibakawa, M.; Oshima, K.; Matsuda, T.; Nadano, D. The mRNA-binding protein Serbp1 as an auxiliary protein associated with mammalian cytoplasmic ribosomes. Cell Biochem. Funct. 2018, 36, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Brugier, A.; Hafirrassou, M.L.; Pourcelot, M.; Baldaccini, M.; Kril, V.; Couture, L.; Amara, A. RACK1 Associates with RNA-Binding Proteins Vigilin and SERBP1 to Facilitate Dengue Virus Replication. J. Virol. 2022, 96, e0196221. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Li, D.; Dong, D.; Li, Y.; Zhang, Y.; Duan, L.; Zhou, J. HIV-1 exposure triggers autophagic degradation of stathmin and hyperstabilization of microtubules to disrupt epithelial cell junctions. Signal Transduct. Target. Ther. 2020, 5, 79. [Google Scholar] [CrossRef]
- Deletsu, S.D.; Kitamura, H.; Ishida, T.; Gohda, J.; Yamaoka, S.; Takeuchi, H. Identification and characterization of Stathmin 1 as a host factor involved in HIV-1 latency. Biochem. Biophys. Res. Commun. 2021, 567, 106–110. [Google Scholar] [CrossRef]
- Bandela, M.; Belvitch, P.; Garcia, J.G.; Dudek, S.M. Cortactin in Lung Cell Function and Disease. Int. J. Mol. Sci. 2022, 23, 4606. [Google Scholar] [CrossRef] [PubMed]
- Hunziker, A.; Glas, I.; Pohl, M.O.; Stertz, S. Phosphoproteomic profiling of influenza virus entry reveals infection-triggered filopodia induction counteracted by dynamic cortactin phosphorylation. Cell Rep. 2022, 38, 110306. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.P.; Nguyen, T.T.; Nguyen, H.C.; Pham, H.T.; Han, K.M.; Choi, D.H.; Hwang, S.B. Cortactin Interacts with Hepatitis C Virus Core and NS5A Proteins: Implications for Virion Assembly. J. Virol. 2020, 94, 01306-20. [Google Scholar] [CrossRef]
- Li, B.; Ding, S.; Feng, N.; Mooney, N.; Ooi, Y.S.; Ren, L.; Diep, J.; Kelly, M.R.; Yasukawa, L.L.; Patton, J.T.; et al. Drebrin restricts rotavirus entry by inhibiting dynamin-mediated endocytosis. Proc. Natl. Acad. Sci. USA 2017, 114, E3642–E3651. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Raduka, A.; Rezaee, F. Respiratory syncytial virus disrupts the airway epithelial barrier by decreasing cortactin and destabilizing F-actin. J. Cell Sci. 2022, 135, 259871. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shang, L.; Zhang, J.; Liu, Y.; Jin, C.; Zhao, Y.; Lei, X.; Wang, W.; Xiao, X.; Zhang, X.; et al. An antibody-based proximity labeling map reveals mechanisms of SARS-CoV-2 inhibition of antiviral immunity. Cell Chem. Biol. 2022, 29, 5–18.e6. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, C.; Hao, P.; Jin, C.; Xu, W.; Liu, Y.; Li, L.; Du, S.; Shang, L.; Jin, X.; Jin, N.; et al. Interaction of Nipah Virus F and G with the Cellular Protein Cortactin Discovered by a Proximity Interactome Assay. Int. J. Mol. Sci. 2024, 25, 4112. https://doi.org/10.3390/ijms25074112
Cui C, Hao P, Jin C, Xu W, Liu Y, Li L, Du S, Shang L, Jin X, Jin N, et al. Interaction of Nipah Virus F and G with the Cellular Protein Cortactin Discovered by a Proximity Interactome Assay. International Journal of Molecular Sciences. 2024; 25(7):4112. https://doi.org/10.3390/ijms25074112
Chicago/Turabian StyleCui, Chunmei, Pengfei Hao, Chaozhi Jin, Wang Xu, Yuchen Liu, Letian Li, Shouwen Du, Limin Shang, Xin Jin, Ningyi Jin, and et al. 2024. "Interaction of Nipah Virus F and G with the Cellular Protein Cortactin Discovered by a Proximity Interactome Assay" International Journal of Molecular Sciences 25, no. 7: 4112. https://doi.org/10.3390/ijms25074112
APA StyleCui, C., Hao, P., Jin, C., Xu, W., Liu, Y., Li, L., Du, S., Shang, L., Jin, X., Jin, N., Wang, J., & Li, C. (2024). Interaction of Nipah Virus F and G with the Cellular Protein Cortactin Discovered by a Proximity Interactome Assay. International Journal of Molecular Sciences, 25(7), 4112. https://doi.org/10.3390/ijms25074112