
Integrated Omics Analysis Uncovers the Culprit behind Exacerbated Atopic Dermatitis in a Diet-Induced Obesity Model

Abstract
1. Introduction
2. Results
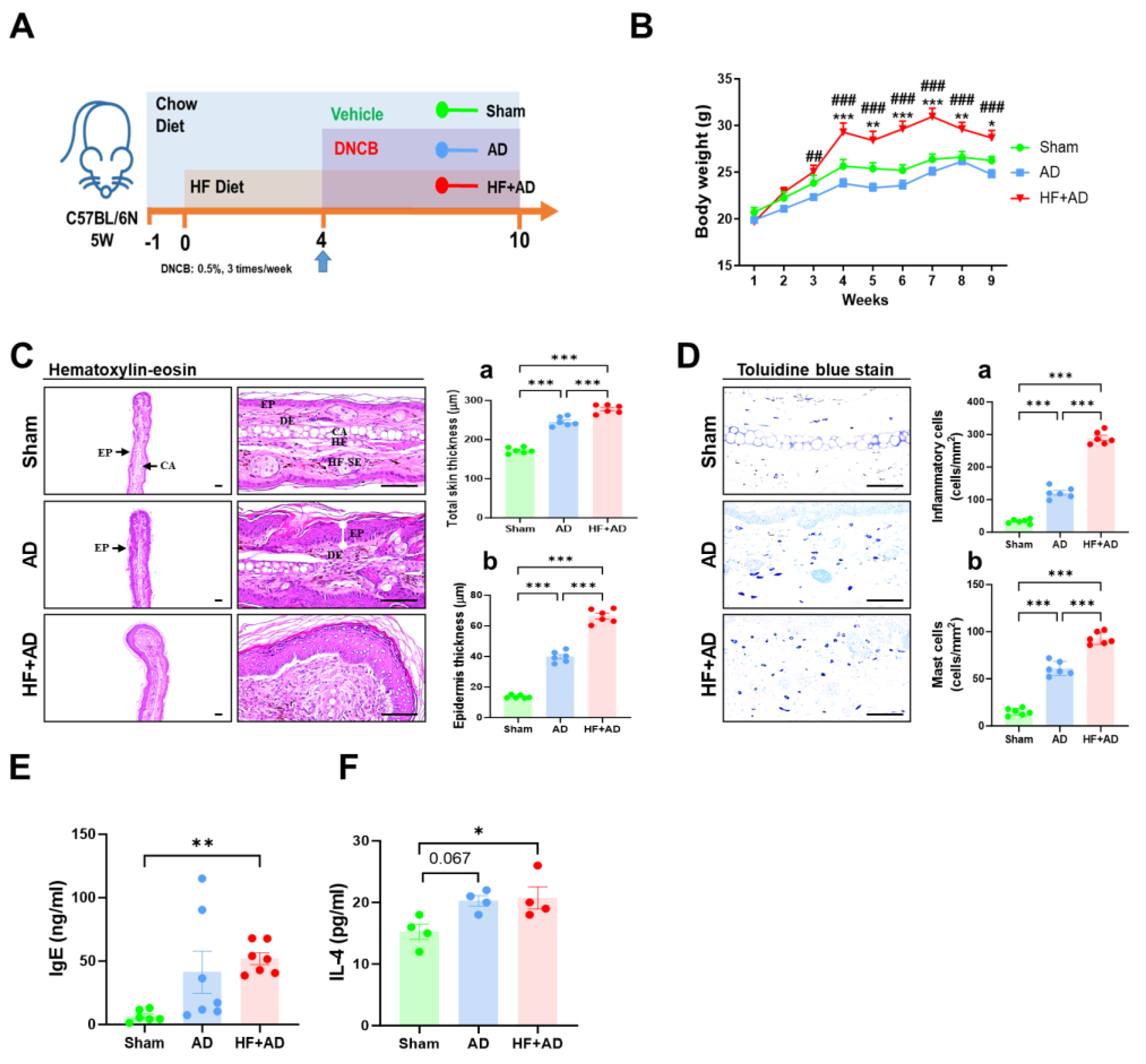
2.1. High-Fat-Diet-Induced Obesity Exacerbates DNCB-Induced AD-Like Lesions
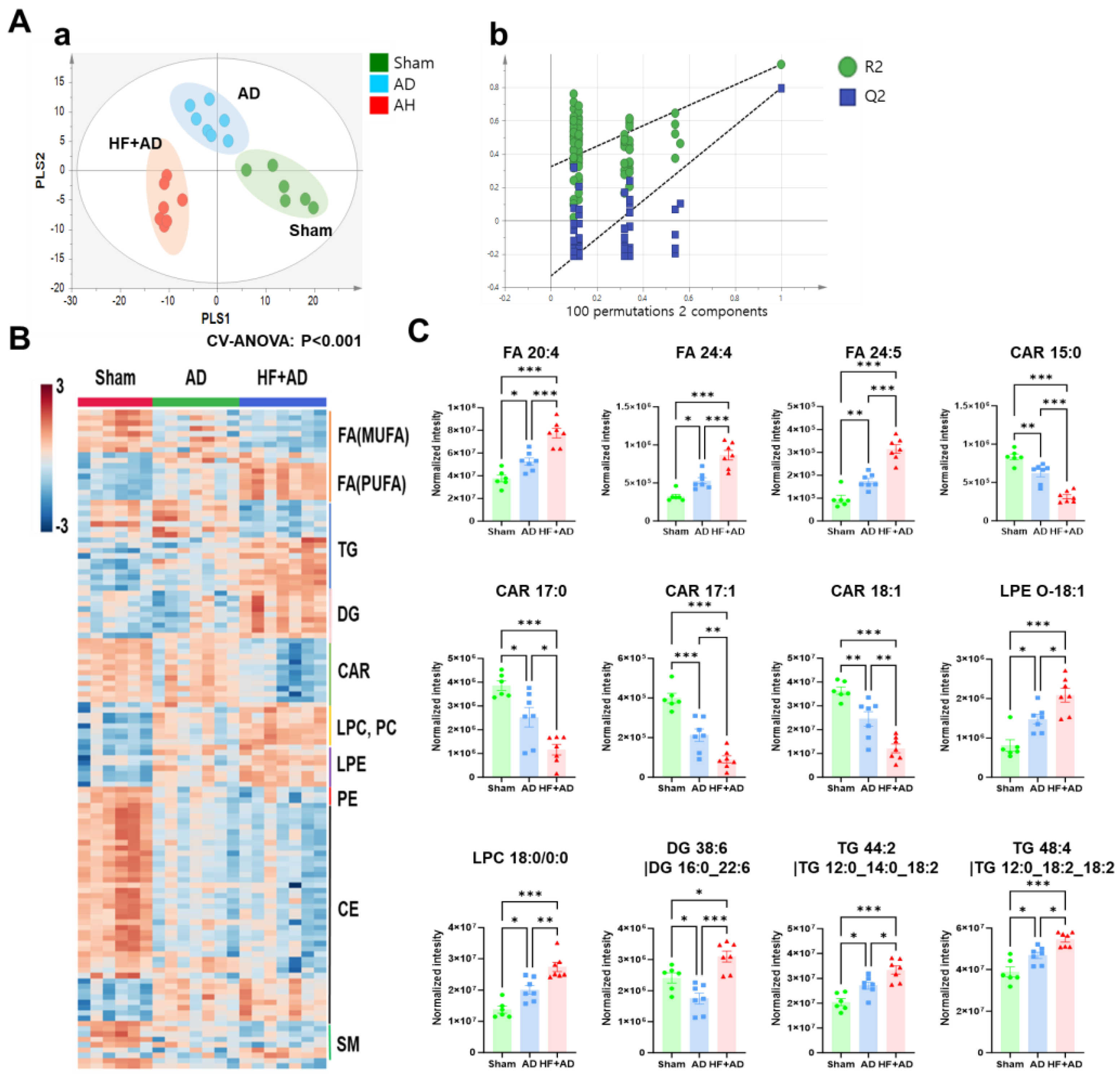
2.2. Untargeted Skin Lipidomics Demonstrates Changes in Lipid Metabolism in HF + AD Mice
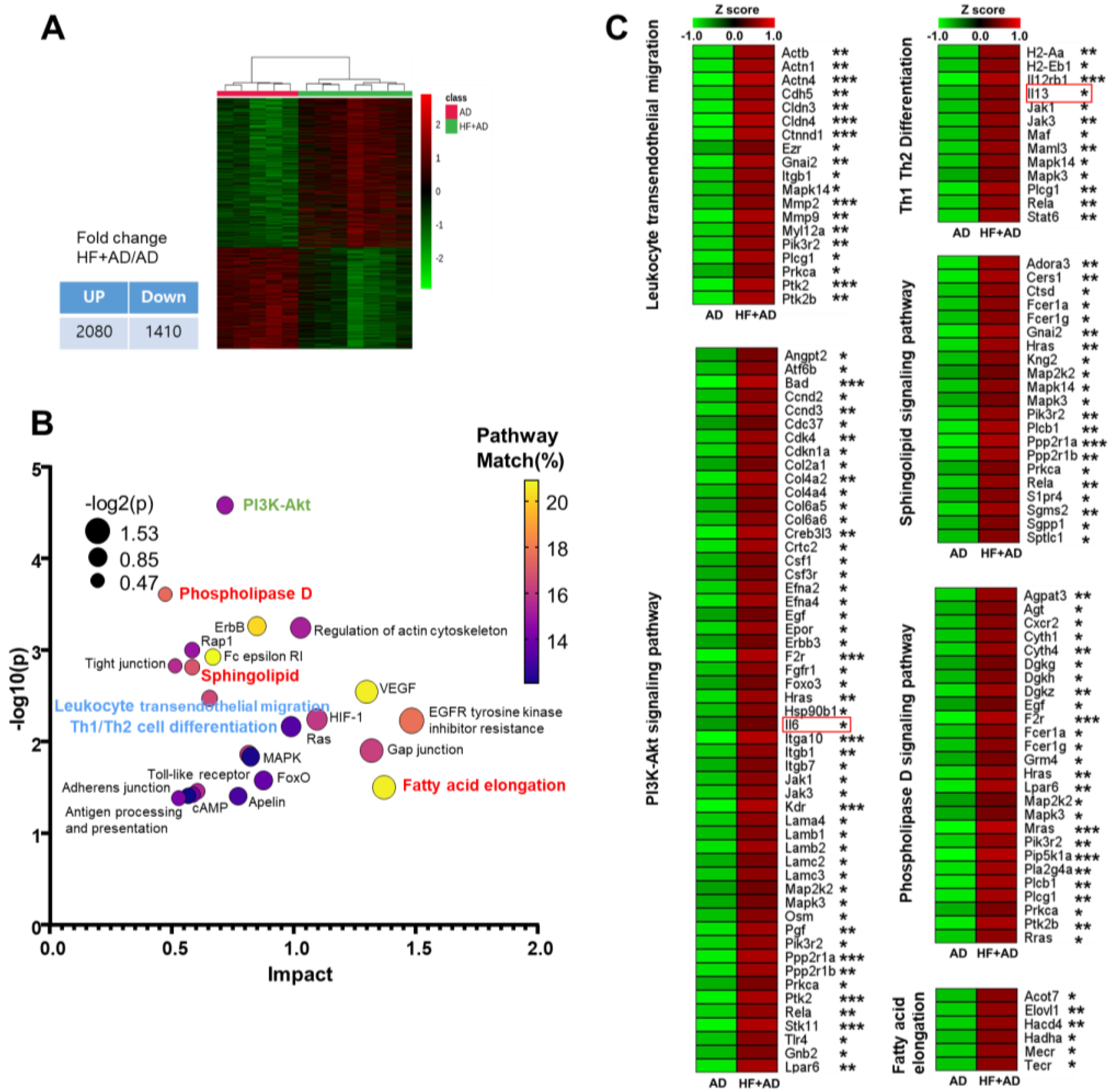
2.3. Transcriptomic Analysis Highlights Distinctions in Pathways Associated with Atopic Immune Responses and Lipid Metabolism between AD and HF + AD
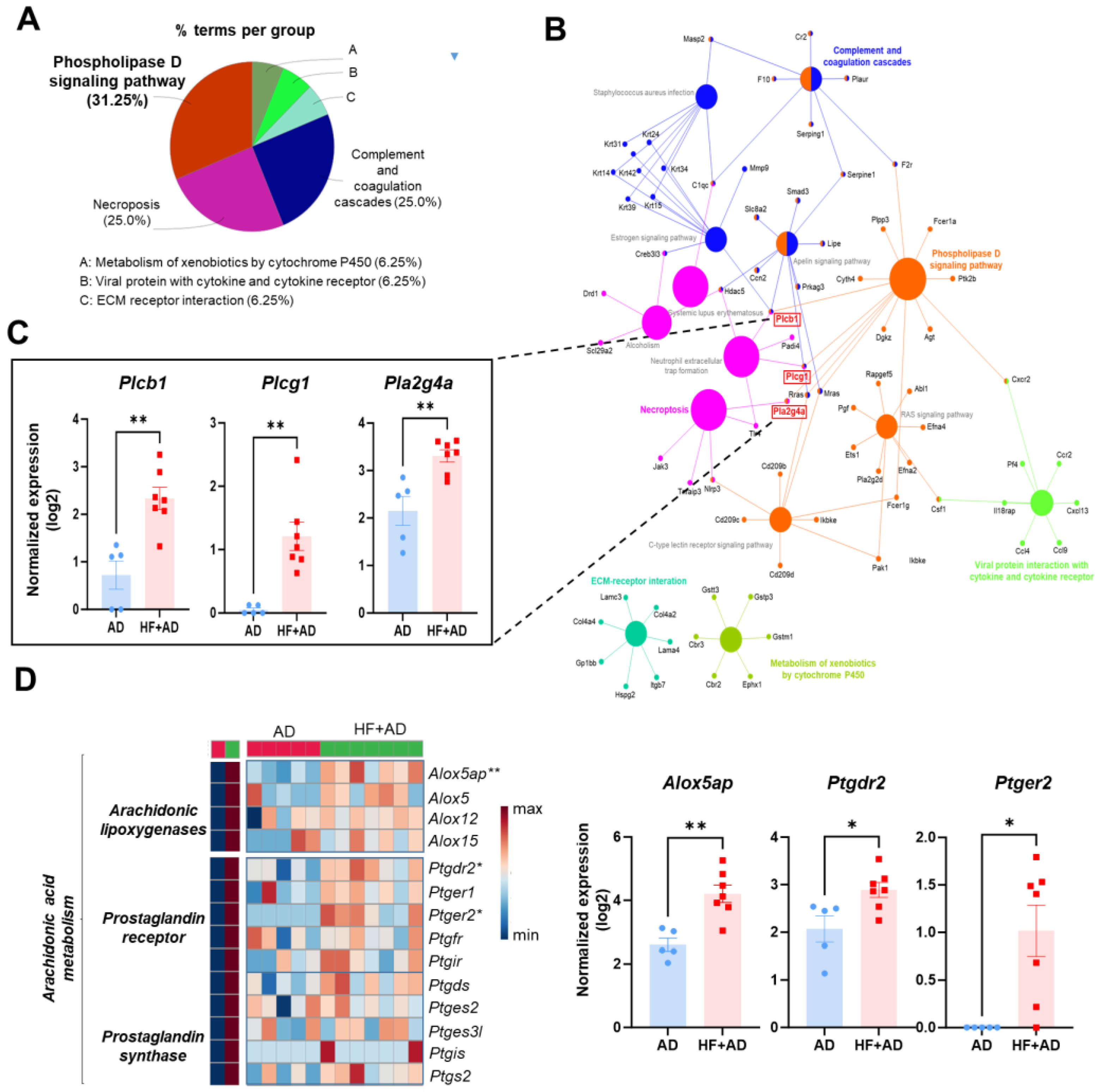
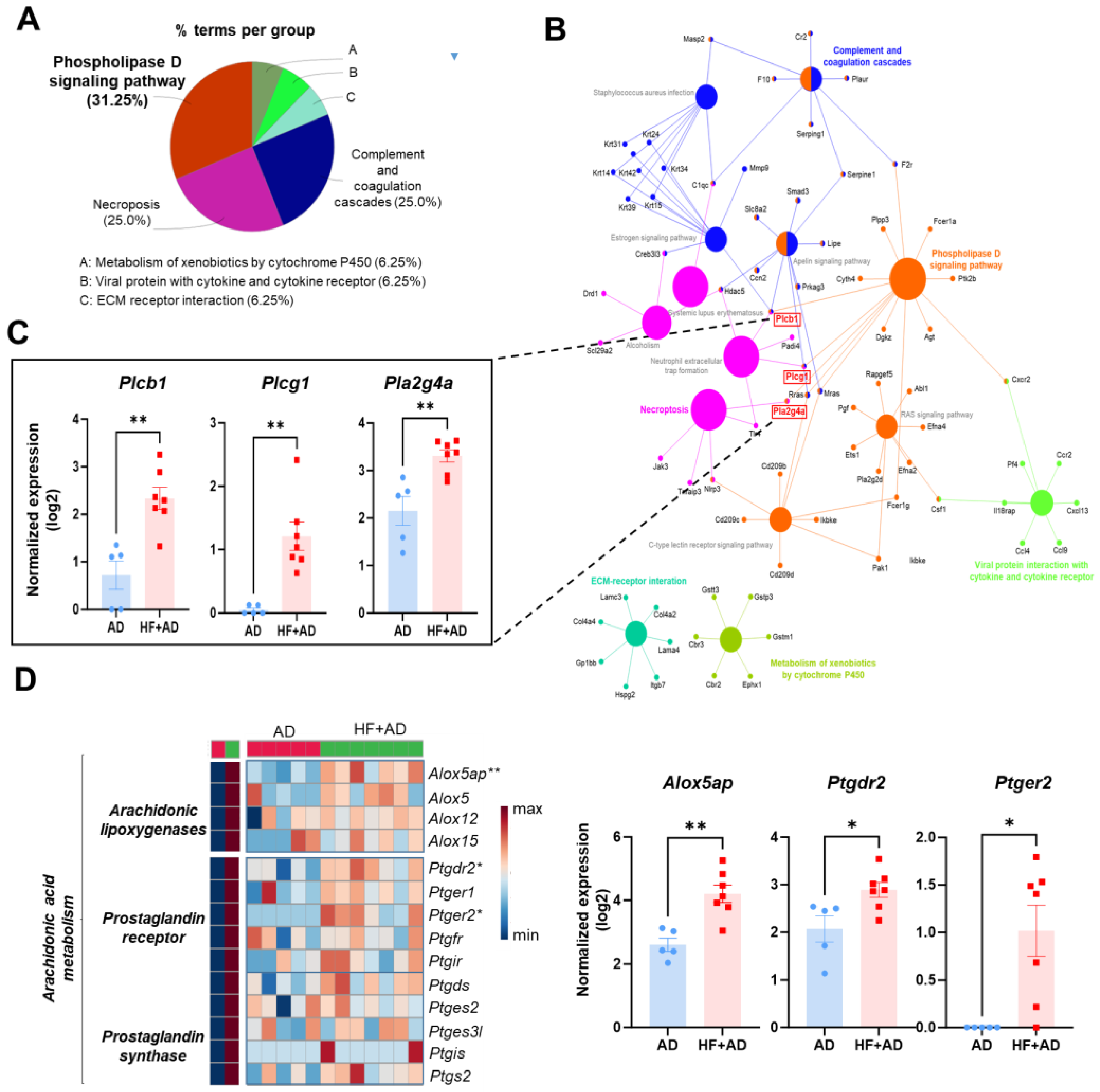
2.4. The Enriched KEGG Pathway Network Reveals the Involvement of Phospholipase Activation in Mediating the Inflammatory Response in the Skin of HF + AD Mice
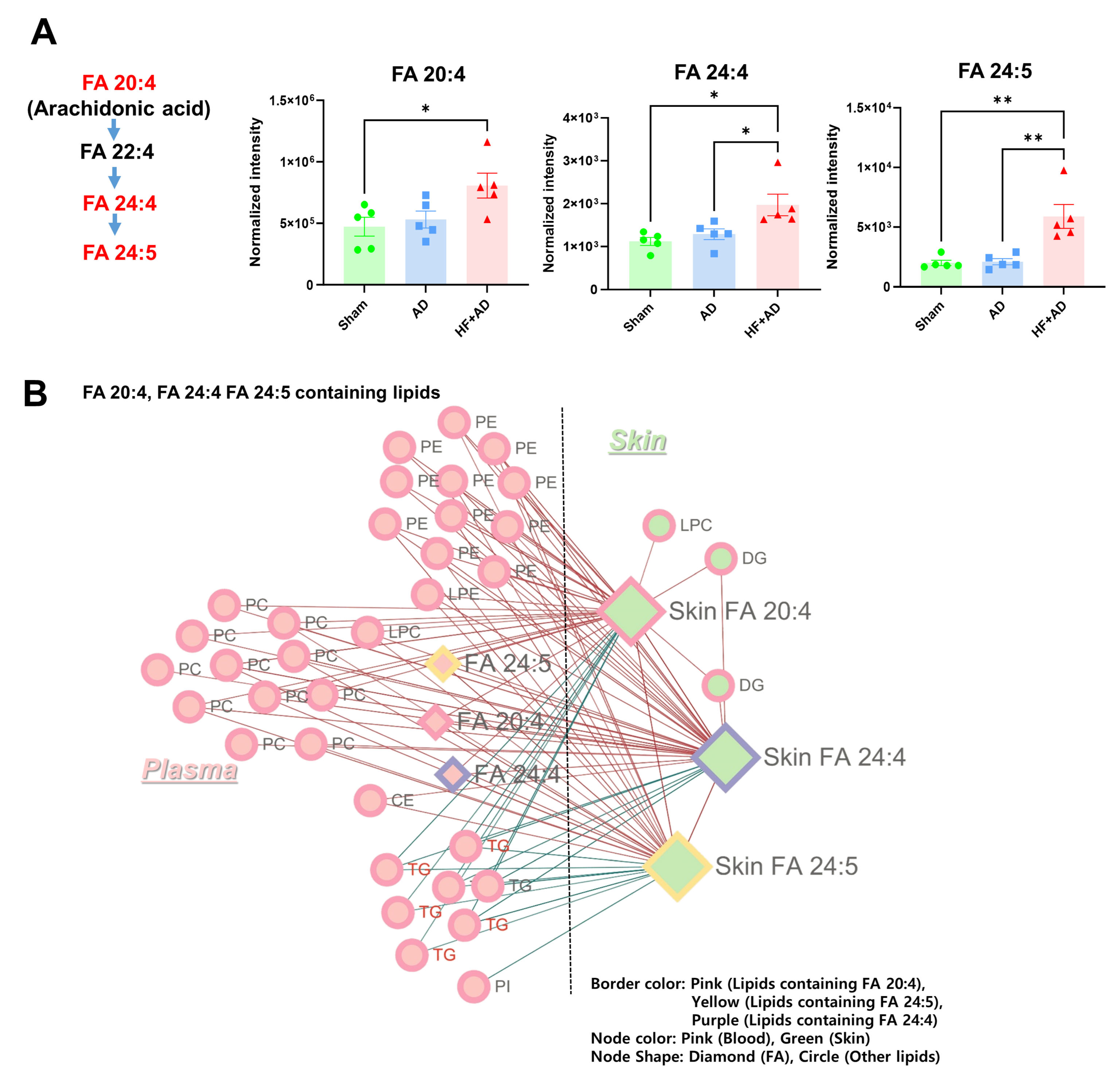
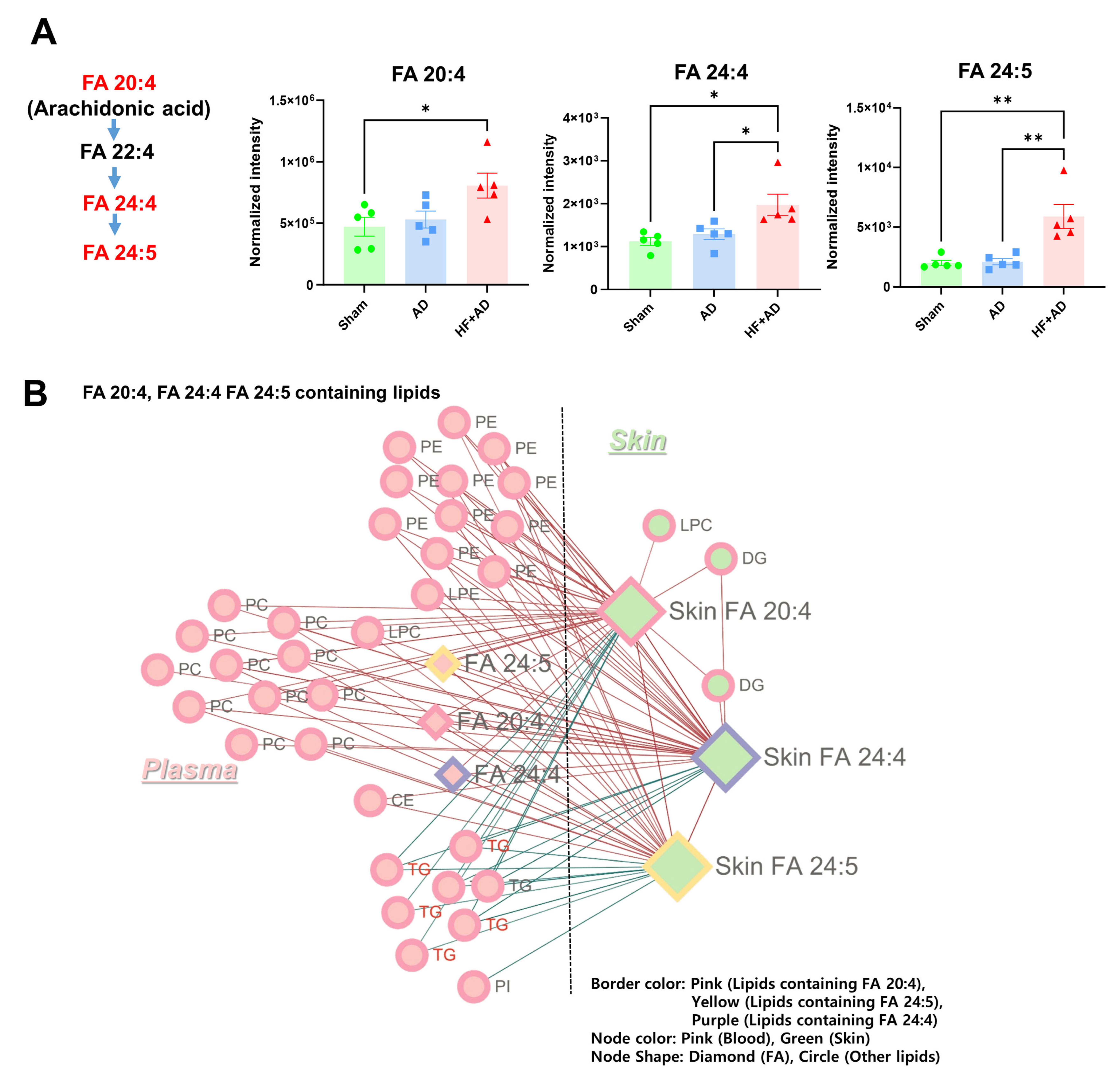
2.5. Circulating AA Correlated with the Changes in AA and Its Metabolites in Skin of HF + AD Mice
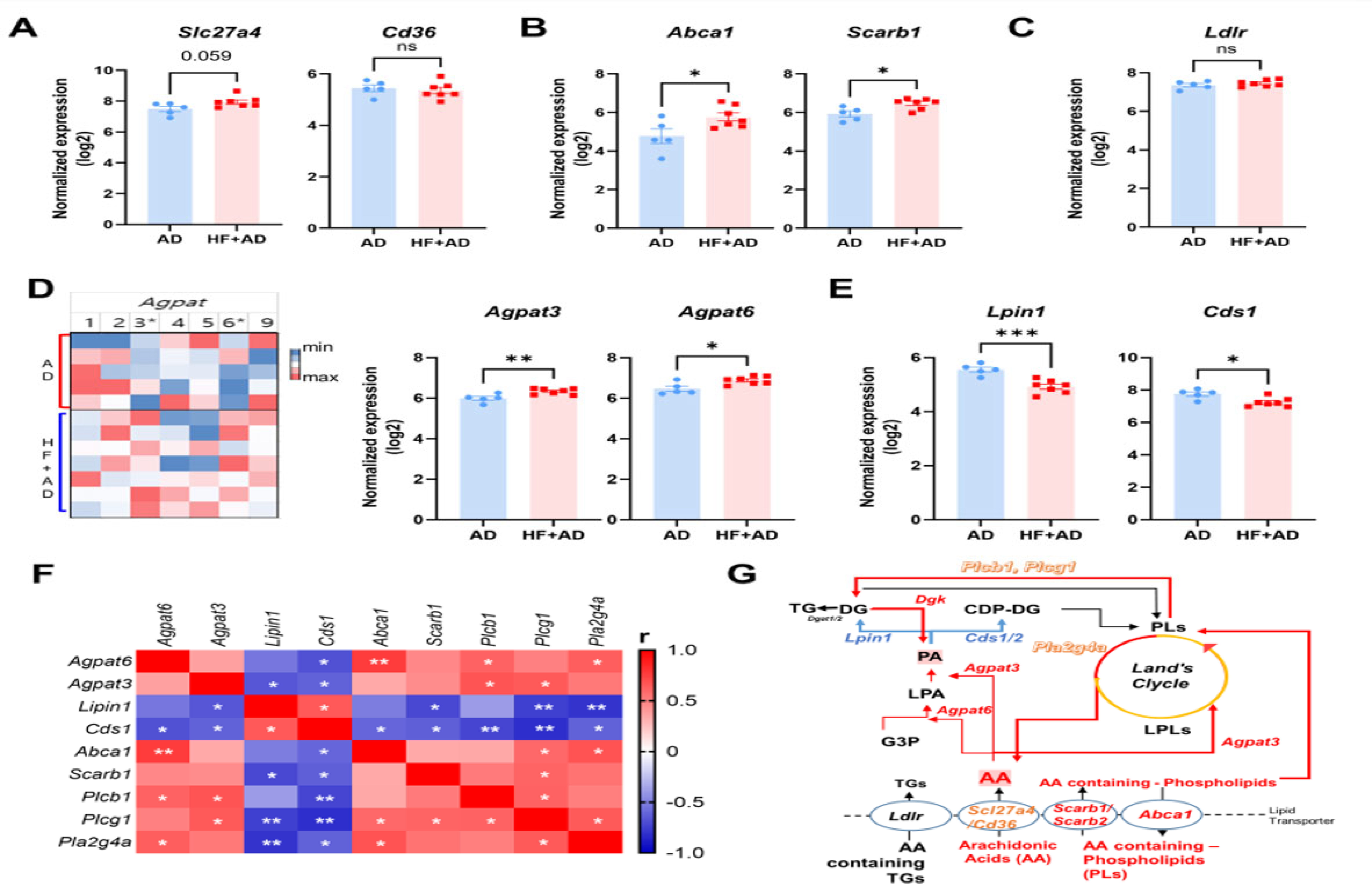
2.6. HF-Diet-Induced Obesity Altered the Expression of Lipid Transporter Genes in Skin of DNCB-Induced AD-like Lesions
2.7. Imbalanced Phospholipid Metabolism Induced by High-Fat-Diet-Aggravated AD in HF + AD Mice
3. Discussion
4. Materials and Methods
4.1. Animal Studies
4.2. Histological Analysis
4.3. Measurement of Plasma Immunoglobulin E (IgE) and Interleukin-4 (IL-4)
4.4. Lipidomics
4.5. RNA-Sequencing (RNA-Seq)
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abuabara, K.; Magyari, A.; McCulloch, C.E.; Linos, E.; Margolis, D.J.; Langan, S.M. Prevalence of Atopic Eczema Among Patients Seen in Primary Care: Data From The Health Improvement Network. Ann. Intern. Med. 2019, 170, 354–356. [Google Scholar] [CrossRef] [PubMed]
- Zissler, U.M.; Chaker, A.M.; Effner, R.; Ulrich, M.; Guerth, F.; Piontek, G.; Dietz, K.; Regn, M.; Knapp, B.; Theis, F.J.; et al. Interleukin-4 and interferon-γ orchestrate an epithelial polarization in the airways. Mucosal Immunol. 2016, 9, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Williams, H.; Stewart, A.; von Mutius, E.; Cookson, W.; Anderson, H.R. Is eczema really on the increase worldwide? J. Allergy Clin. Immunol. 2008, 121, 947–954.e15. [Google Scholar] [CrossRef] [PubMed]
- Nahm, D.H. Associations of Atopic Dermatitis with Obesity and Unmarried Status in Young Adults: Evidence for Atopic Dermatitis as a Life-Style Disorder With High Social Impact. Allergy Asthma Immunol. Res. 2016, 8, 89–91. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Wu, X.; Zhou, Y.; Sheng, L.; Jena, P.K.; Han, D.; Wan, Y.J.Y.; Hwang, S.T. A Western Diet, but Not a High-Fat and Low-Sugar Diet, Predisposes Mice to Enhanced Susceptibility to Imiquimod-Induced Psoriasiform Dermatitis. J. Investig. Dermatol. 2019, 139, 1404–1407. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Silverberg, J.I. Association of atopic dermatitis with being overweight and obese: A systematic review and metaanalysis. J. Am. Acad. Dermatol. 2015, 72, 606–616.e4. [Google Scholar] [CrossRef]
- Jeong, K.Y.; Lee, J.; Li, C.; Han, T.; Lee, S.B.; Lee, H.; Back, S.K.; Na, H.S. Juvenile obesity aggravates disease severity in a rat model of atopic dermatitis. Allergy Asthma Immunol. Res. 2015, 7, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Savetsky, I.L.; Albano, N.J.; Cuzzone, D.A.; Gardenier, J.C.; Torrisi, J.S.; García Nores, G.D.; Nitti, M.D.; Hespe, G.E.; Nelson, T.S.; Kataru, R.P.; et al. Lymphatic Function Regulates Contact Hypersensitivity Dermatitis in Obesity. J. Investig. Dermatol. 2015, 135, 2742–2752. [Google Scholar] [CrossRef] [PubMed]
- Jakwerth, C.A.; Chaker, A.M.; Guerth, F.; Oelsner, M.; Pechtold, L.; Zur Bonsen, L.S.; Ullmann, J.T.; Krauss-Etschmann, S.; Erb, A.; Kau, J.; et al. Sputum microRNA-screening reveals Prostaglandin EP3 receptor as selective target in allergen-specific immunotherapy. Clin. Exp. Allergy 2021, 51, 1577–1591. [Google Scholar] [CrossRef]
- Gutmann, D.; Dressler, M.; Eickmeier, O.; Herrmann, E.; Kirwil, M.; Schubert, R.; Zielen, S.; Zissler, U.M. Proinflammatory pattern in the lower airways of non-asthmatic obese adolescents. Cytokine 2024, 173, 156452. [Google Scholar] [CrossRef]
- Hornburg, D.; Wu, S.; Moqri, M.; Zhou, X.; Contrepois, K.; Bararpour, N.; Traber, G.M.; Su, B.; Metwally, A.A.; Avina, M.; et al. Dynamic lipidome alterations associated with human health, disease and ageing. Nat. Metab. 2023, 5, 1578–1594. [Google Scholar] [CrossRef] [PubMed]
- Knox, S.; O’Boyle, N.M. Skin lipids in health and disease: A review. Chem. Phys. Lipids 2021, 236, 105055. [Google Scholar] [CrossRef] [PubMed]
- Lydic, T.A.; Goo, Y.H. Lipidomics unveils the complexity of the lipidome in metabolic diseases. Clin. Transl. Med. 2018, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Lowe, R.; Shirley, N.; Bleackley, M.; Dolan, S.; Shafee, T. Transcriptomics technologies. PLoS Comput. Biol. 2017, 13, e1005457. [Google Scholar] [CrossRef]
- Basil, M.C.; Levy, B.D. Specialized pro-resolving mediators: Endogenous regulators of infection and inflammation. Nat. Rev. Immunol. 2016, 16, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Bieber, T. Interleukin-13: Targeting an underestimated cytokine in atopic dermatitis. Allergy 2020, 75, 54–62. [Google Scholar] [CrossRef]
- Xiao, D.; Xu, P.; Deng, H.; Chen, L.-S.; Ding, X.-A.; Bai, M.; Li, P.-C.; Yuan, D.-F. The characterization and clinical significance of PI3K/Akt signaling pathway activation in the peripheral T cells of pediatric patients with atopic dermatitis. Int. J. Clin. Exp. Med. 2017, 10, 2904–2910. [Google Scholar]
- Zhu, L.; Jones, C.; Zhang, G. The Role of Phospholipase C Signaling in Macrophage-Mediated Inflammatory Response. J. Immunol. Res. 2018, 2018, 5201759. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Q.; Kuo, M.S.; Li, S.; Bui, H.H.; Peake, D.A.; Sanders, P.E.; Thibodeaux, S.J.; Chu, S.; Qian, Y.W.; Zhao, Y.; et al. AGPAT6 is a novel microsomal glycerol-3-phosphate acyltransferase. J. Biol. Chem. 2008, 283, 10048–10057. [Google Scholar] [CrossRef]
- Prasad, S.S.; Garg, A.; Agarwal, A.K. Enzymatic activities of the human AGPAT isoform 3 and isoform 5: Localization of AGPAT5 to mitochondria. J. Lipid Res. 2011, 52, 451–462. [Google Scholar] [CrossRef]
- Pérez-Pérez, A.; Sánchez-Jiménez, F.; Vilariño-García, T.; Sánchez-Margalet, V. Role of Leptin in Inflammation and Vice Versa. Int. J. Mol. Sci. 2020, 21, 5887. [Google Scholar] [CrossRef]
- Jiménez-Cortegana, C.; Ortiz-García, G.; Serrano, A.; Moreno-Ramírez, D.; Sánchez-Margalet, V. Possible Role of Leptin in Atopic Dermatitis: A Literature Review. Biomolecules 2021, 11, 1642. [Google Scholar] [CrossRef]
- Zheng, H.; Wu, D.; Wu, X.; Zhang, X.; Zhou, Q.; Luo, Y.; Yang, X.; Chock, C.J.; Liu, M.; Yang, X.O. Leptin Promotes Allergic Airway Inflammation through Targeting the Unfolded Protein Response Pathway. Sci. Rep. 2018, 8, 8905. [Google Scholar] [CrossRef]
- Lin, L.L.; Wartmann, M.; Lin, A.Y.; Knopf, J.L.; Seth, A.; Davis, R.J. cPLA2 is phosphorylated and activated by MAP kinase. Cell 1993, 72, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, L.; Kragballe, K. Abnormalities in epidermal lipid metabolism in patients with atopic dermatitis. J. Investig. Dermatol. 1991, 96, 10–15. [Google Scholar] [CrossRef]
- Nakano, N.; Nakao, A.; Uchida, T.; Shirasaka, N.; Yoshizumi, H.; Okumura, K.; Tsuboi, R.; Ogawa, H. Effects of arachidonic acid analogs on FcεRI-mediated activation of mast cells. Biochim. Et Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2005, 1738, 19–28. [Google Scholar] [CrossRef]
- Serrano-Mollar, A.; Closa, D. Arachidonic acid signaling in pathogenesis of allergy: Therapeutic implications. Curr. Drug Targets Inflamm. Allergy 2005, 4, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Ryborg, A.K.; Grøn, B.; Kragballe, K. Increased lysophosphatidylcholine content in lesional psoriatic skin. Br. J. Dermatol. 1995, 133, 398–402. [Google Scholar] [CrossRef]
- Ryborg, A.K.; Deleuran, B.; Søgaard, H.; Kragballe, K. Intracutaneous injection of lysophosphatidylcholine induces skin inflammation and accumulation of leukocytes. Acta Derm. Venereol. 2000, 80, 242–246. [Google Scholar]
- Moreno, J.J.; Ferrer, X.; Ortega, E.; Carganico, G. PLA2-induced oedema in rat skin and histamine release in rat mast cells. Evidence for involvement of lysophospholipids in the mechanism of action. Agents Actions 1992, 36, 258–263. [Google Scholar]
- Dan, P.; Rosenblat, G.; Yedgar, S. Phospholipase A2 activities in skin physiology and pathology. Eur. J. Pharmacol. 2012, 691, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Ohata, H.; Honda, K. Lysophosphatidic acid (LPA) induces plasma exudation and histamine release in mice via LPA receptors. J. Pharmacol. Sci. 2006, 100, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Mruwat, R.; Cohen, Y.; Yedgar, S. Phospholipase A(2) inhibition as potential therapy for inflammatory skin diseases. Immunotherapy 2013, 5, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Urban, S.; Zieseniss, S.; Werder, M.; Hauser, H.; Budzinski, R.; Engelmann, B. Scavenger Receptor BI Transfers Major Lipoprotein-associated Phospholipids into the Cells*. J. Biol. Chem. 2000, 275, 33409–33415. [Google Scholar] [CrossRef]
- Kadamur, G.; Ross, E.M. Mammalian phospholipase C. Annu. Rev. Physiol. 2013, 75, 127–154. [Google Scholar] [CrossRef]
- Sakane, F.; Hoshino, F.; Murakami, C. New Era of Diacylglycerol Kinase, Phosphatidic Acid and Phosphatidic Acid-Binding Protein. Int. J. Mol. Sci. 2020, 21, 6794. [Google Scholar] [CrossRef]
- Harris, T.E.; Finck, B.N. Dual function lipin proteins and glycerolipid metabolism. Trends Endocrinol. Metab. 2011, 22, 226–233. [Google Scholar] [CrossRef]
- Kuchler, K.; Daum, G.; Paltauf, F. Subcellular and submitochondrial localization of phospholipid-synthesizing enzymes in Saccharomyces cerevisiae. J. Bacteriol. 1986, 165, 901–910. [Google Scholar] [CrossRef]
- Lim, H.K.; Choi, Y.A.; Park, W.; Lee, T.; Ryu, S.H.; Kim, S.Y.; Kim, J.R.; Kim, J.H.; Baek, S.H. Phosphatidic acid regulates systemic inflammatory responses by modulating the Akt-mammalian target of rapamycin-p70 S6 kinase 1 pathway. J. Biol. Chem. 2003, 278, 45117–45127. [Google Scholar] [CrossRef]
- Fang, Y.; Vilella-Bach, M.; Bachmann, R.; Flanigan, A.; Chen, J. Phosphatidic Acid-Mediated Mitogenic Activation of mTOR Signaling. Science 2001, 294, 1942–1945. [Google Scholar] [CrossRef]
- Mao-Qiang, M.; Feingold, K.R.; Jain, M.; Elias, P.M. Extracellular processing of phospholipids is required for permeability barrier homeostasis. J. Lipid Res. 1995, 36, 1925–1935. [Google Scholar] [CrossRef]
- Lu, B.; Jiang, Y.J.; Man, M.Q.; Brown, B.; Elias, P.M.; Feingold, K.R. Expression and regulation of 1-acyl-sn-glycerol- 3-phosphate acyltransferases in the epidermis. J. Lipid Res. 2005, 46, 2448–2457. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Ohn, J.; Kim, J.W.; Kang, S.M.; Jeon, D.; Heo, C.Y.; Lee, Y.S.; Kwon, O.; Kim, K.H. Caffeoyl-Pro-His amide relieve DNCB-Induced Atopic Dermatitis-Like phenotypes in BALB/c mice. Sci. Rep. 2020, 10, 8417. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.G.; Kang, M.; Lee, Y.-H.; Min, W.G.; Kim, Y.H.; Kang, S.J.; Song, C.H.; Park, S.J.; Park, J.H.; Han, C.H.; et al. Bathing Effects of Various Seawaters on Allergic (Atopic) Dermatitis-Like Skin Lesions Induced by 2,4-Dinitrochlorobenzene in Hairless Mice. Evid.-Based Complement. Altern. Med. 2015, 2015, 179185. [Google Scholar] [CrossRef]
- Kim, K.H.; Park, S.J.; Lee, Y.J.; Lee, J.E.; Song, C.H.; Choi, S.H.; Ku, S.K.; Kang, S.J. Inhibition of UVB-induced skin damage by exopolymers from Aureobasidium pullulans SM-2001 in hairless mice. Basic Clin. Pharmacol. Toxicol. 2015, 116, 73–86. [Google Scholar] [CrossRef]
- Kim, W.Y.; Kim, H.G.; Lee, H.W.; Lee, J.S.; Im, H.J.; Kim, H.S.; Lee, S.B.; Son, C.G. A Herbal Formula, Atofreellage, Ameliorates Atopic Dermatitis-Like Skin Lesions in an NC/Nga Mouse Model. Molecules 2015, 21, E35. [Google Scholar] [CrossRef]
- Park, S.H.; Lee, J.E.; Lee, S.M.; Lee, J.; Seo, C.S.; Hwang, G.S.; Jung, J. An unbiased lipidomics approach identifies key lipid molecules as potential therapeutic targets of Dohongsamul-tang against non-alcoholic fatty liver diseases in a mouse model of obesity. J. Ethnopharmacol. 2020, 260, 112999. [Google Scholar] [CrossRef]
- Lee, J.E.; Lee, S.M.; Jung, J. Integrated omics analysis unraveled the microbiome-mediated effects of Yijin-Tang on hepatosteatosis and insulin resistance in obese mouse. Phytomedicine 2020, 79, 153354. [Google Scholar] [CrossRef]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 2004, 5, R80. [Google Scholar] [CrossRef]
- Barsky, A.; Gardy, J.L.; Hancock, R.E.W.; Munzner, T. Cerebral: A Cytoscape plugin for layout of and interaction with biological networks using subcellular localization annotation. Bioinformatics 2007, 23, 1040–1042. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway Name | Converted Lipids | p-Value | Benjamin Correction | Bonferroni Correction |
|---|---|---|---|---|
| Arachidonic acid metabolism | 100 | 8.52 × 10−7 | 5.11 × 10−6 | 5.11 × 10−6 |
| Inflammatory mediator regulation of TRP channels | 28.57 | 0.02 | 0.05 | 0.11 |
| PPAR signaling pathway | 14.29 | 0.06 | 0.12 | 0.39 |
| Vascular smooth muscle contraction | 14.29 | 0.06 | 0.12 | 0.61 |
| Aldosterone synthesis and secretion | 14.29 | 0.1 | 0.12 | 0.61 |
| Serotonergic synapse | 14.29 | 0.36 | 0.36 | 1 |
| Mediators | LLODs | HLODs | |
|---|---|---|---|
| Basic | IgE | 1.37 ng/mL | 1000 ng/mL |
| Th1/Th2 | IL–4 | 0.36 pg/mL | 5947 pg/mL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, Y.M.; Jung, J.; Lee, S.M. Integrated Omics Analysis Uncovers the Culprit behind Exacerbated Atopic Dermatitis in a Diet-Induced Obesity Model. Int. J. Mol. Sci. 2024, 25, 4143. https://doi.org/10.3390/ijms25084143
Ahn YM, Jung J, Lee SM. Integrated Omics Analysis Uncovers the Culprit behind Exacerbated Atopic Dermatitis in a Diet-Induced Obesity Model. International Journal of Molecular Sciences. 2024; 25(8):4143. https://doi.org/10.3390/ijms25084143
Chicago/Turabian StyleAhn, You Mee, Jeeyoun Jung, and So Min Lee. 2024. "Integrated Omics Analysis Uncovers the Culprit behind Exacerbated Atopic Dermatitis in a Diet-Induced Obesity Model" International Journal of Molecular Sciences 25, no. 8: 4143. https://doi.org/10.3390/ijms25084143
APA StyleAhn, Y. M., Jung, J., & Lee, S. M. (2024). Integrated Omics Analysis Uncovers the Culprit behind Exacerbated Atopic Dermatitis in a Diet-Induced Obesity Model. International Journal of Molecular Sciences, 25(8), 4143. https://doi.org/10.3390/ijms25084143