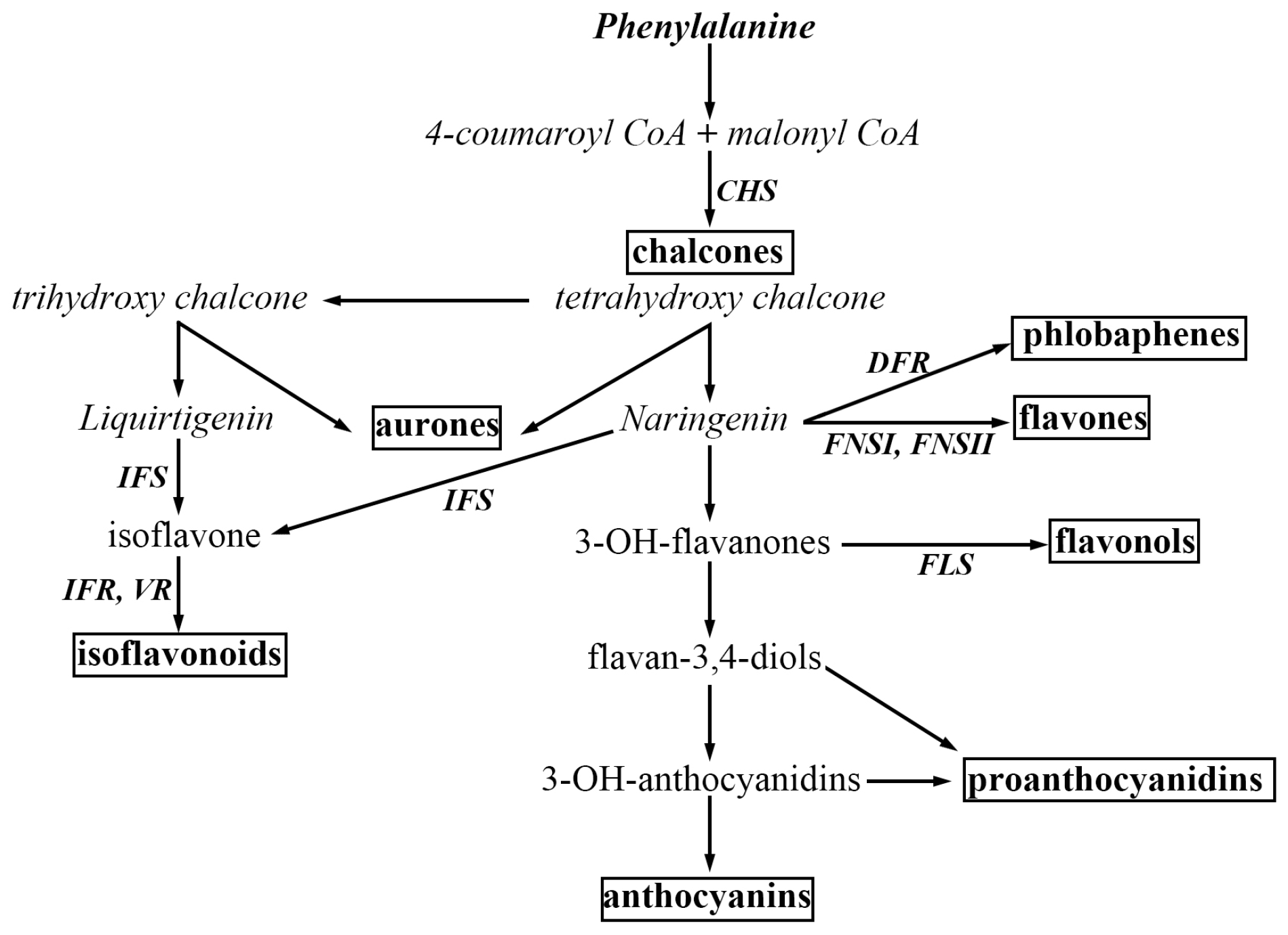
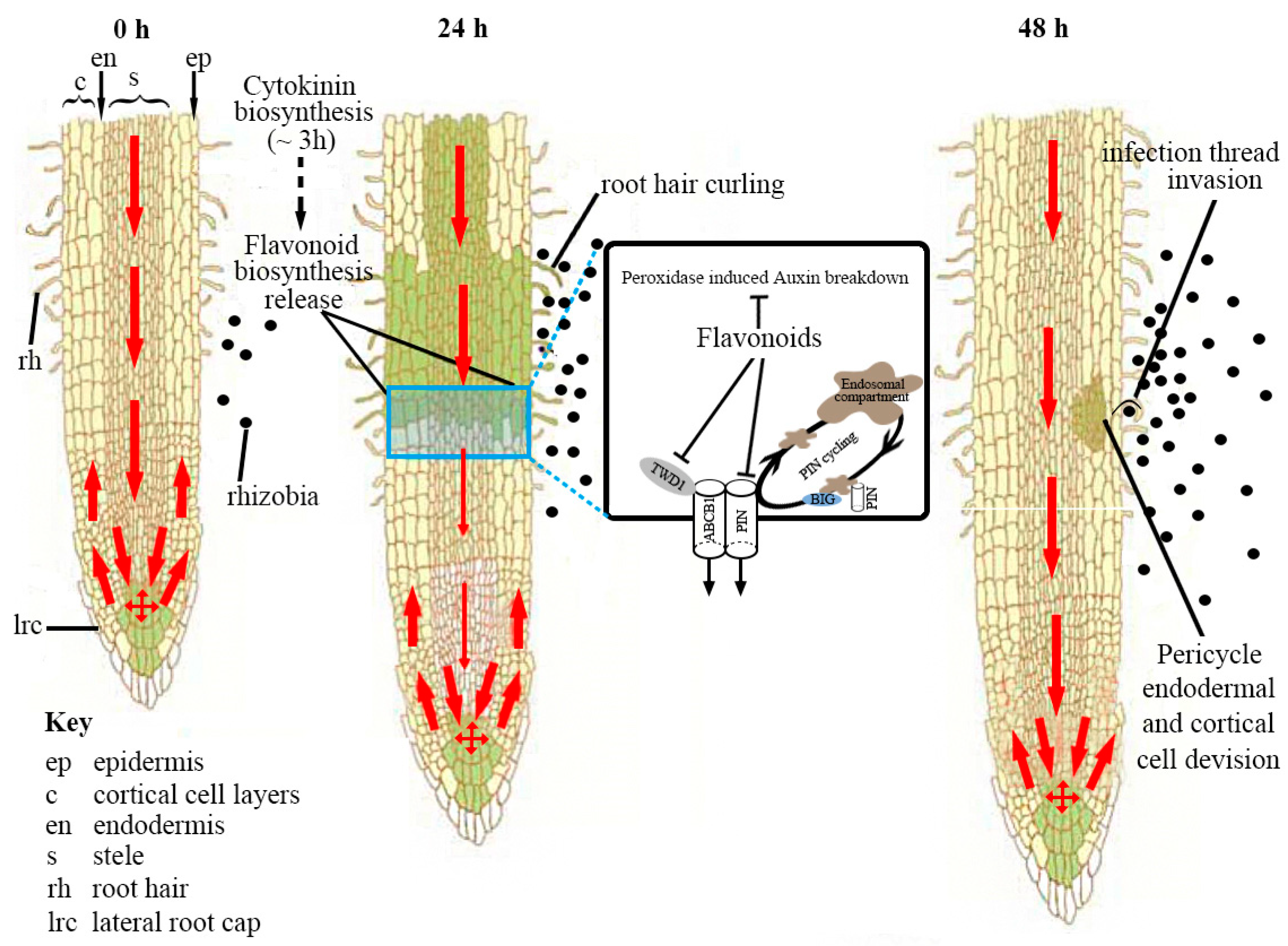
Correction: Dong, W.; Song, Y. The Significance of Flavonoids in the Process of Biological Nitrogen Fixation. Int. J. Mol. Sci. 2020, 21, 5926

{kind=link}
{kind=link}
- 66.
- Ng, J.L.; Perrine-Walker, F.; Wasson, A.P.; Mathesius, U. The Control of Auxin Transport in Parasitic and Symbiotic Root-Microbe Interactions. Plants 2015, 4, 606–643.
Reference
- Dong, W.; Song, Y. The Significance of Flavonoids in the Process of Biological Nitrogen Fixation. Int. J. Mol. Sci. 2020, 21, 5926. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, W.; Song, Y. Correction: Dong, W.; Song, Y. The Significance of Flavonoids in the Process of Biological Nitrogen Fixation. Int. J. Mol. Sci. 2020, 21, 5926. Int. J. Mol. Sci. 2024, 25, 4144. https://doi.org/10.3390/ijms25084144
Dong W, Song Y. Correction: Dong, W.; Song, Y. The Significance of Flavonoids in the Process of Biological Nitrogen Fixation. Int. J. Mol. Sci. 2020, 21, 5926. International Journal of Molecular Sciences. 2024; 25(8):4144. https://doi.org/10.3390/ijms25084144
Chicago/Turabian StyleDong, Wei, and Yuguang Song. 2024. "Correction: Dong, W.; Song, Y. The Significance of Flavonoids in the Process of Biological Nitrogen Fixation. Int. J. Mol. Sci. 2020, 21, 5926" International Journal of Molecular Sciences 25, no. 8: 4144. https://doi.org/10.3390/ijms25084144
APA StyleDong, W., & Song, Y. (2024). Correction: Dong, W.; Song, Y. The Significance of Flavonoids in the Process of Biological Nitrogen Fixation. Int. J. Mol. Sci. 2020, 21, 5926. International Journal of Molecular Sciences, 25(8), 4144. https://doi.org/10.3390/ijms25084144