
Organophosphorus Flame Retardant TPP-Induced Human Corneal Epithelial Cell Apoptosis through Caspase-Dependent Mitochondrial Pathway



Abstract
1. Introduction
2. Results and Discussion
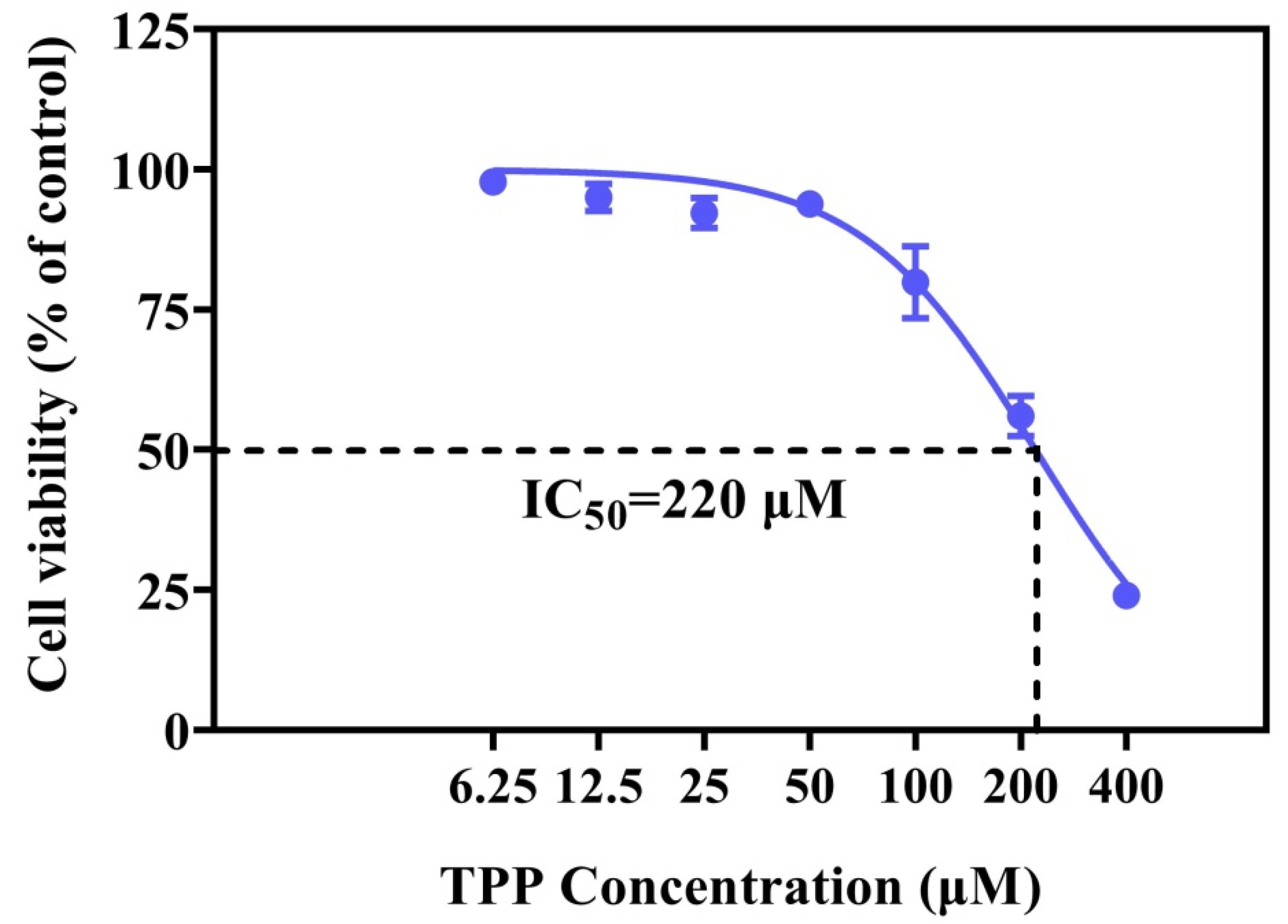
2.1. TPP Suppressed Cell Viability and Altered Cell Morphology
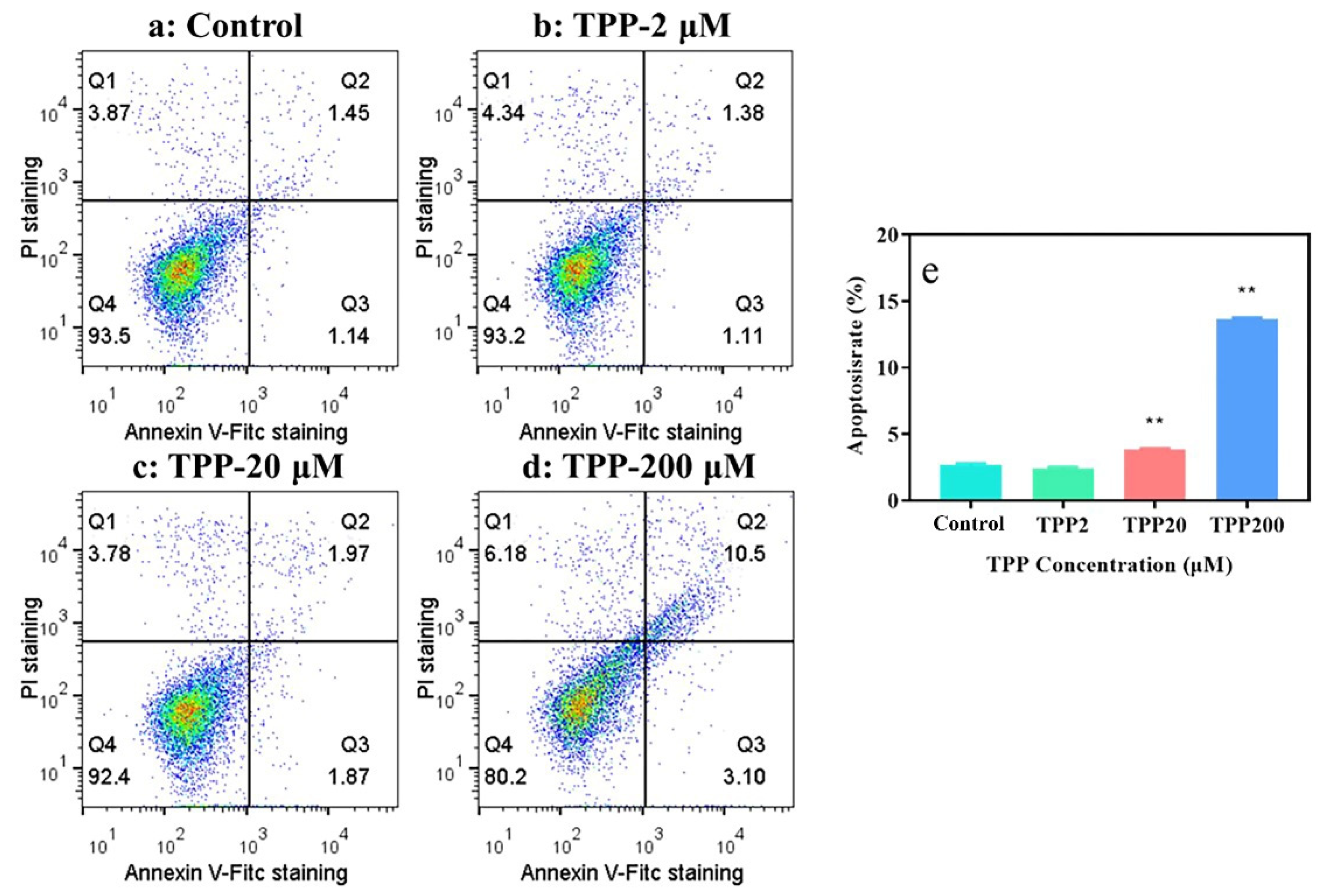
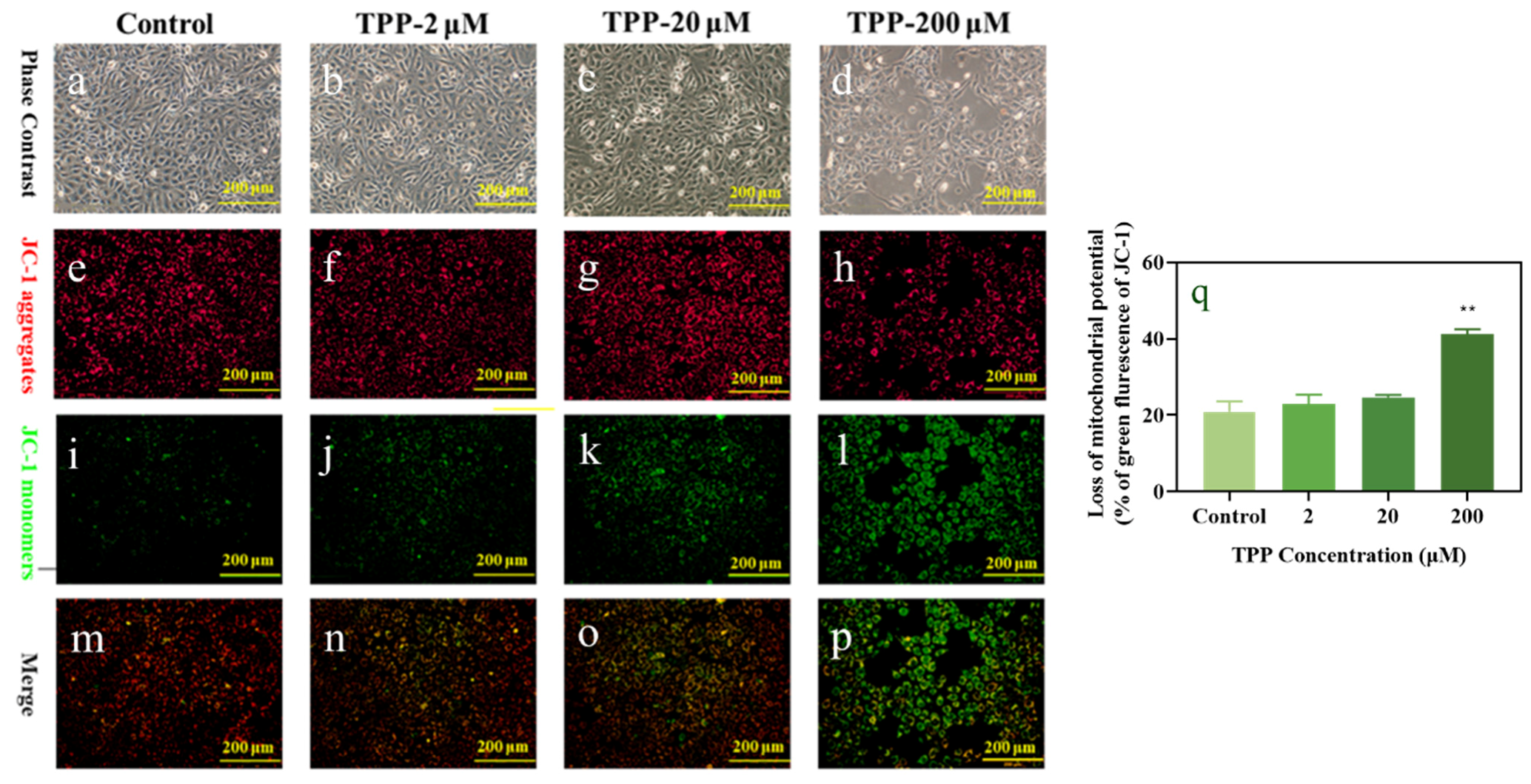
2.2. TPP Induced Apoptosis and Changed Mitochondrial Membrane Potential in HCECs
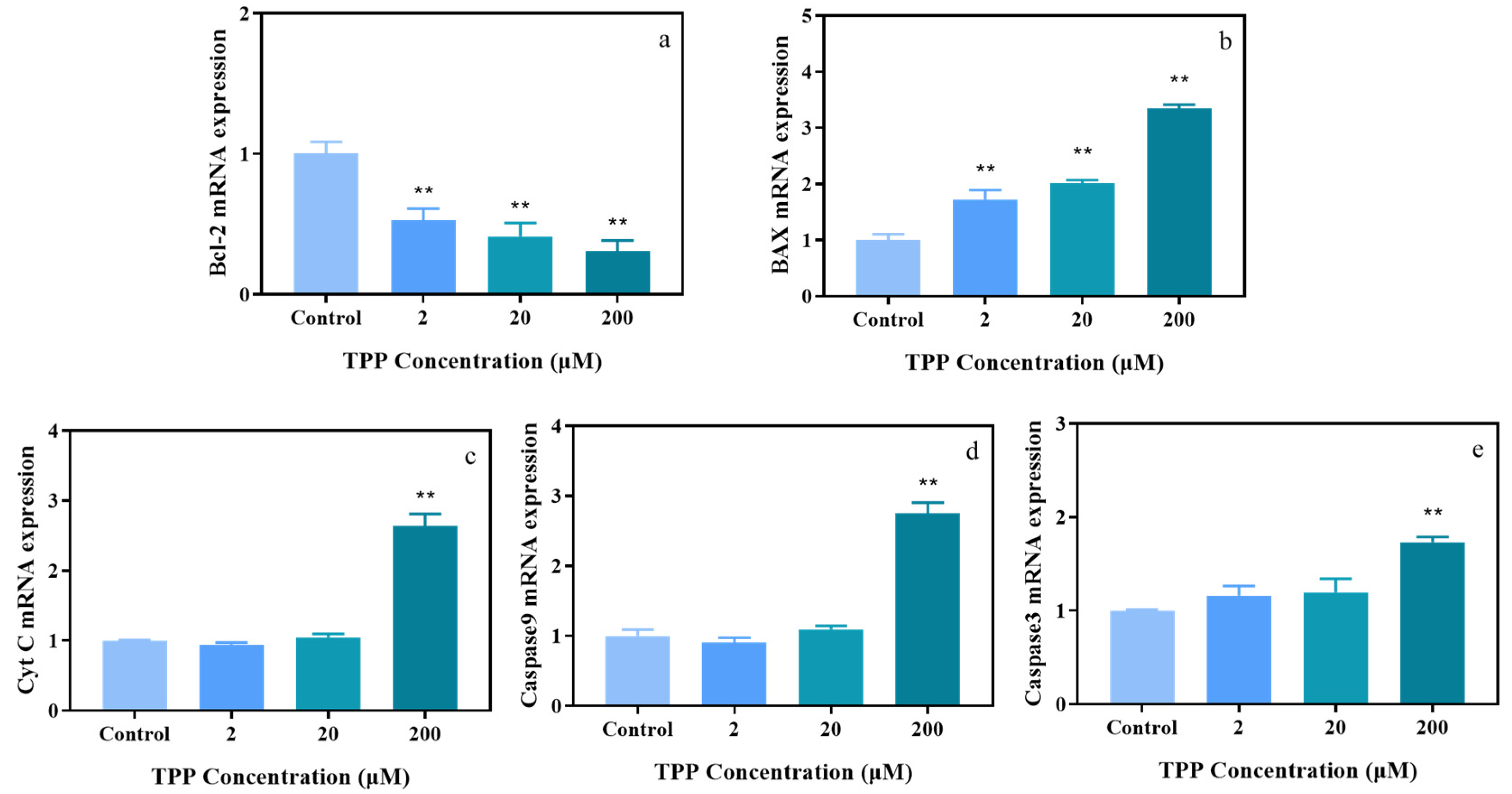
2.3. TPP Affects Expression Level of Genes Associated with Apoptosis in HCECs
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Cell Viability, Morphology, and Apoptosis
3.3. Mitochondrial Membrane Potential Assay
3.4. Cell Total RNA Extraction and cDNA Synthesis and Quantitative Real-Time PCR Analysis
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luo, Y.; Guo, W.; Ngo, H.H.; Nghiem, L.D.; Hai, F.I.; Zhang, J.; Liang, S.; Wang, X.C. A review on the occurrence of micropollutants in the aquatic environment and their fate and removal during wastewater treatment. Sci. Total Environ. 2014, 473–474, 619–641. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.Q.; Zhu, H.K.; Shi, Y.M.; Xu, K.; Chen, S.C.; Zou, Q.; Sun, H.W.; Kannan, K. Profiling of multiple classes of flame retardants in house dust in China: Pattern analysis and human exposure assessment. Environ. Pollut. 2022, 311, 9. [Google Scholar] [CrossRef] [PubMed]
- He, M.J.; Lu, J.F.; Ma, J.Y.; Wang, H.; Du, X.F. Organophosphate esters and phthalate esters in human hair from rural and urban areas, Chongqing, China: Concentrations, composition profiles and sources in comparison to street dust. Environ. Pollut. 2018, 237, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hou, M.; Zhang, Q.; Wu, X.; Zhao, H.; Xie, Q.; Chen, J. Organophosphorus flame retardants and plasticizers in building and decoration materials and their potential burdens in newly decorated houses in China. Environ. Sci. Technol. 2017, 51, 10991–10999. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Wang, Y.; Yao, Y.M.; Ren, C.; Lan, Z.H.; Fang, X.G.; Zhang, K.; Sun, W.J.; Alder, A.C.; Sun, H.W. Occurrence of organophosphate flame retardants in farmland soils from Northern China: Primary source analysis and risk assessment. Environ. Pollut. 2019, 247, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xu, C.; Song, N.; Zhang, S.; Bu, Y.; Xiong, L.; Yin, L.; Pu, Y.; Zhang, J. Seasonal variation and health risk assessment of organophosphate esters in surface and drinking water in Nanjing, China. Int. J. Environ. Sci. Technol. 2023, 20, 411–422. [Google Scholar] [CrossRef]
- Azizi, S.; Dehghani, M.H.; Naddafi, K.; Nabizadeh, R.; Yunesian, M. Occurrence of organophosphorus esters in outdoor air fine particulate matter and comprehensive assessment of human exposure: A global systematic review. Environ. Pollut. 2023, 318, 11. [Google Scholar] [CrossRef]
- Wu, M.; Yu, G.; Cao, Z.G.; Wu, D.K.; Liu, K.; Deng, S.B.; Huang, J.; Wang, B.; Wang, Y.J. Characterization and human exposure assessment of organophosphate flame retardants in indoor dust from several microenvironments of Beijing, China. Chemosphere 2016, 150, 465–471. [Google Scholar] [CrossRef]
- Kurt-Karakus, P.; Alegria, H.; Birgul, A.; Gungormus, E.; Jantunen, L. Organophosphate ester (OPEs) flame retardants and plasticizers in air and soil from a highly industrialized city in Turkey. Sci. Total Environ. 2018, 625, 555–565. [Google Scholar] [CrossRef]
- Han, X.; Hao, Y.F.; Li, Y.M.; Yang, R.Q.; Wang, P.; Zhang, G.X.; Zhang, Q.H.; Jiang, G.B. Occurrence and distribution of organophosphate esters in the air and soils of Ny-Alesund and London Island, Svalbard, Arctic. Environ. Pollut. 2020, 263, 7. [Google Scholar] [CrossRef]
- Yan, Z.F.; Feng, C.L.; Leung, K.M.Y.; Luo, Y.; Wang, J.D.; Jin, X.W.; Wu, F.C. Insights into the geographical distribution, bioaccumulation characteristics, and ecological risks of organophosphate esters. J. Hazard. Mater. 2023, 445, 16. [Google Scholar] [CrossRef] [PubMed]
- Schindler, B.K.; Weiss, T.; Schütze, A.; Koslitz, S.; Broding, H.C.; Bünger, J.; Brüning, T. Occupational exposure of air crews to tricresyl phosphate isomers and organophosphate flame retardants after fume events. Arch. Toxicol. 2013, 87, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-W.; Isobe, T.; Muto, M.; Tue, N.M.; Katsura, K.; Malarvannan, G.; Sudaryanto, A.; Chang, K.-H.; Prudente, M.; Viet, P.H.; et al. Organophosphorus flame retardants (PFRs) in human breast milk from several Asian countries. Chemosphere 2014, 116, 91–97. [Google Scholar] [CrossRef]
- Guo, Y.T.; Chen, M.Q.; Liao, M.X.; Su, S.J.; Sun, W.Y.; Gan, Z.W. Organophosphorus flame retardants and their metabolites in paired human blood and urine. Ecotoxicol. Environ. Saf. 2023, 268, 12. [Google Scholar] [CrossRef] [PubMed]
- Sutton, W.L.; Terhaar, C.J.; Miller, F.A.; Scherberger, R.F.; Riley, E.C.; Roudabush, R.L.; Fassett, D.W. Studies on the Industrial Hygiene and Toxicology of Triphenyl Phosphate. Arch. Environ. Health Int. J. 1960, 1, 33–46. [Google Scholar] [CrossRef]
- Zhang, Q.; Zheng, S.; Shi, X.; Luo, C.; Huang, W.; Lin, H.; Peng, J.; Tan, W.; Wu, K. Neurodevelopmental toxicity of organophosphate flame retardant triphenyl phosphate (TPhP) on zebrafish (Danio rerio) at different life stages. Environ. Int. 2023, 172, 107745. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Wang, C.; Zeng, L.; Peng, Y.; Li, Y.; Hao, H.; Zheng, Y.; Chen, C.; Chen, H.; Zhang, J.; et al. Triphenyl phosphate induced reproductive toxicity through the JNK signaling pathway in Caenorhabditis elegans. J. Hazard. Mater. 2023, 446, 130643. [Google Scholar] [CrossRef]
- Witchey, S.K.; Sutherland, V.; Collins, B.; Roberts, G.; Shockley, K.R.; Vallant, M.; Krause, J.; Cunny, H.; Waidyanatha, S.; Mylchreest, E.; et al. Reproductive and developmental toxicity following exposure to organophosphate ester flame retardants and plasticizers, triphenyl phosphate and isopropylated phenyl phosphate, in Sprague Dawley rats. Toxicol. Sci. Off. J. Soc. Toxicol. 2023, 191, 374–386. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Liu, G.; Yuan, L.-X.; Yang, J.; Liu, J.; Li, Z.; Yang, C.; Wang, J. Triphenyl phosphate (TPP) promotes hepatocyte toxicity via induction of endoplasmic reticulum stress and inhibition of autophagy flux. Sci. Total Environ. 2022, 840, 156461. [Google Scholar] [CrossRef]
- Liu, X.S.; Zhao, X.L.; Wang, Y.; Hong, J.B.; Shi, M.; Pfaff, D.; Guo, L.X.; Tang, H.W. Triphenyl phosphate permeates the blood brain barrier and induces neurotoxicity in mouse brain. Chemosphere 2020, 252, 12. [Google Scholar] [CrossRef]
- Wang, X.Q.; Li, F.; Liu, J.L.; Ji, C.L.; Wu, H.F. Transcriptomic, proteomic and metabolomic profiling unravel the mechanisms of hepatotoxicity pathway induced by triphenyl phosphate (TPP). Ecotoxicol. Environ. Saf. 2020, 205, 111126. [Google Scholar] [CrossRef]
- Jannuzzi, A.T.; Goler, A.M.Y.; Alpertunga, B. Ubiquitin proteasomal system is a potential target of the toxic effects of organophosphorus flame retardant triphenyl phosphate. Environ. Toxicol. Pharmacol. 2022, 96, 10. [Google Scholar] [CrossRef]
- Wang, Y.; Hong, J.B.; Shi, M.; Guo, L.X.; Liu, L.H.; Tang, H.W.; Liu, X.S. Triphenyl phosphate disturbs the lipidome and induces endoplasmic reticulum stress and apoptosis in JEG-3 cells. Chemosphere 2021, 275, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.N.; Yang, D.L.; Liu, H.; Bi, J.; Bao, Y.B.; Ma, J.Y.; Zheng, Q.X.; Cui, D.L.; Chen, W.; Xiang, P. Effects of TCPP and TCEP exposure on human corneal epithelial cells: Oxidative damage, cell cycle arrest, and pyroptosis. Chemosphere 2023, 331, 11. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Tong, L.; Mehta, J.S.; Ong, H.S. Impact of Exposomes on Ocular Surface Diseases. Int. J. Mol. Sci. 2023, 24, 1273. [Google Scholar] [CrossRef] [PubMed]
- Mimura, T.; Ichinose, T.; Yamagami, S.; Fujishima, H.; Kamei, Y.; Goto, M.; Takada, S. Matsubara M: Airborne particulate matter (PM2.5) and the prevalence of allergic conjunctivitis in Japan. Sci. Total Environ. 2014, 487, 493–499. [Google Scholar] [CrossRef]
- Xiang, P.; Liu, R.-Y.; Sun, H.-J.; Han, Y.-H.; He, R.-W.; Cui, X.-Y.; Ma, L.Q. Molecular mechanisms of dust-induced toxicity in human corneal epithelial cells: Water and organic extract of office and house dust. Environ. Int. 2016, 92–93, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Ma, J.-Y.; Li, M.-Y.; Qin, Y.-S.; Bao, X.-C.; Wang, C.-C.; Cui, D.-L.; Xiang, P.; Ma, L.Q. Mechanisms of Cd and Cu induced toxicity in human gastric epithelial cells: Oxidative stress, cell cycle arrest and apoptosis. Sci. Total Environ. 2021, 756, 143951. [Google Scholar] [CrossRef] [PubMed]
- Xiang, P.; Liu, R.-Y.; Li, C.; Gao, P.; Cui, X.-Y.; Ma, L.Q. Effects of organophosphorus flame retardant TDCPP on normal human corneal epithelial cells: Implications for human health. Environ. Pollut. 2017, 230, 22–30. [Google Scholar] [CrossRef]
- He, J.; Li, J.; Ma, L.; Wu, N.; Zhang, Y.; Niu, Z. Large-scale distribution of organophosphate esters (flame retardants and plasticizers) in soil from residential area across China: Implications for current level. Sci. Total Environ. 2019, 697, 133997. [Google Scholar] [CrossRef]
- Matsumoto, S.; Way, W.; Tarlo, K.; Short, B. Comparative toxicity of fluoroquinolone antibiotics on corneal cells in vitro. Cornea 2006, 25, S1–S7. [Google Scholar] [CrossRef]
- Clark, W.C. Sustainability science: A room of its own. Natl. Acad Sci. 2007, 104, 1737–1738. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Xiong, N.-X.; Fang, Z.-X.; Kuang, X.-Y.; Wang, F.; Ou, J.; Luo, S.-W. Upregulation of oxidative stress by triphenyl phosphate (TPhP) exposure causes antioxidant insult and apoptotic process in Epithelioma papulosum cyprini (EPC) cells. Environ. Sci. Pollut. Res. 2023, 30, 119217–119227. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Sato, E.F.; Nishikawa, M.; Park, A.M.; Kira, Y.; Imada, I.; Utsumi, K. Mitochondrial generation of reactive oxygen species and its role in aerobic life. Curr. Med. Chem. 2003, 10, 2495–2505. [Google Scholar] [CrossRef] [PubMed]
- Szabo, A.; Balog, M.; Mark, L.; Montsko, G.; Turi, Z.; Gallyas, F., Jr.; Sumegi, B.; Kalai, T.; Hideg, K.; Kovacs, K. Induction of mitochondrial destabilization and necrotic cell death by apolar mitochondria-directed SOD mimetics. Mitochondrion 2011, 11, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Perelman, A.; Wachtel, C.; Cohen, M.; Haupt, S.; Shapiro, H.; Tzur, A. JC-1: Alternative excitation wavelengths facilitate mitochondrial membrane potential cytometry. Cell Death Dis. 2012, 3, e430. [Google Scholar] [CrossRef] [PubMed]
- Xiang, P.; He, R.-W.; Liu, R.-Y.; Li, K.; Gao, P.; Cui, X.-Y.; Li, H.; Liu, Y.; Ma, L.Q. Cellular responses of normal (HL-7702) and cancerous (HepG2) hepatic cells to dust extract exposure. Chemosphere 2018, 193, 1189–1197. [Google Scholar] [CrossRef]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef]
- Javadov, S.; Karmazyn, M. Mitochondrial permeability transition pore opening as an endpoint to initiate cell death and as a putative target for cardioprotection. Cell. Physiol. Biochem. 2006, 20, 1–22. [Google Scholar] [CrossRef]
- Pal, M.K.; Jaiswar, S.P.; Srivastav, A.K.; Goyal, S.; Dwivedi, A.; Verma, A.; Singh, J.; Pathak, A.K.; Sankhwar, P.L.; Ray, R.S. Synergistic effect of piperine and paclitaxel on cell fate via cyt-c, Bax/Bcl-2-caspase-3 pathway in ovarian adenocarcinomas SKOV-3 cells. Eur. J. Pharmacol. 2016, 791, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Zaib, S.; Hayyat, A.; Ali, N.; Gul, A.; Naveed, M.; Khan, I. Role of Mitochondrial Membrane Potential and Lactate Dehydrogenase A in Apoptosis. Anti-Cancer Agents Med. Chem. 2022, 22, 2048–2062. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Yin, H.; Peng, H.; Lu, G.; Liu, Z.; Dang, Z. OPFRs and BFRs induced A549 cell apoptosis by caspase-dependent mitochondrial pathway. Chemosphere 2019, 221, 693–702. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| β-actin | GACATCCGCAAAGACCTG | GGAAGGTGGACAGCGAG |
| Bcl-2 | CATGGAAGCGAATCAATGGACT | CTGTACCAGACCGAGATGTCA |
| Bax | AAACTGGTGCTCAAGGCCC | CTTGGATCCAGACAAGCAGC |
| Caspase-3 | CATGGAAGCGAATCAATGGACT | CTGTACCAGACCGAGATGTCA |
| Caspase-9 | CTCAGACCAGAGATTCGCAAAC | GCATTTCCCCTCAAACTCTCAA |
| Cyt c | AGGCCCCTGGTACTCTTACACA | TCTGCCCTTTCTTCCTTCTTCTTA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Cui, D.; Li, J.; Ren, C.; Yang, D.; Xiang, P.; Liu, J. Organophosphorus Flame Retardant TPP-Induced Human Corneal Epithelial Cell Apoptosis through Caspase-Dependent Mitochondrial Pathway. Int. J. Mol. Sci. 2024, 25, 4155. https://doi.org/10.3390/ijms25084155
Chen C, Cui D, Li J, Ren C, Yang D, Xiang P, Liu J. Organophosphorus Flame Retardant TPP-Induced Human Corneal Epithelial Cell Apoptosis through Caspase-Dependent Mitochondrial Pathway. International Journal of Molecular Sciences. 2024; 25(8):4155. https://doi.org/10.3390/ijms25084155
Chicago/Turabian StyleChen, Chen, Daolei Cui, Jingya Li, Caixia Ren, Danlei Yang, Ping Xiang, and Jianxiang Liu. 2024. "Organophosphorus Flame Retardant TPP-Induced Human Corneal Epithelial Cell Apoptosis through Caspase-Dependent Mitochondrial Pathway" International Journal of Molecular Sciences 25, no. 8: 4155. https://doi.org/10.3390/ijms25084155
APA StyleChen, C., Cui, D., Li, J., Ren, C., Yang, D., Xiang, P., & Liu, J. (2024). Organophosphorus Flame Retardant TPP-Induced Human Corneal Epithelial Cell Apoptosis through Caspase-Dependent Mitochondrial Pathway. International Journal of Molecular Sciences, 25(8), 4155. https://doi.org/10.3390/ijms25084155