
Chromosomal Damage, Chromosome Instability, and Polymorphisms in GSTP1 and XRCC1 as Biomarkers of Effect and Susceptibility in Farmers Exposed to Pesticides
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Study Groups
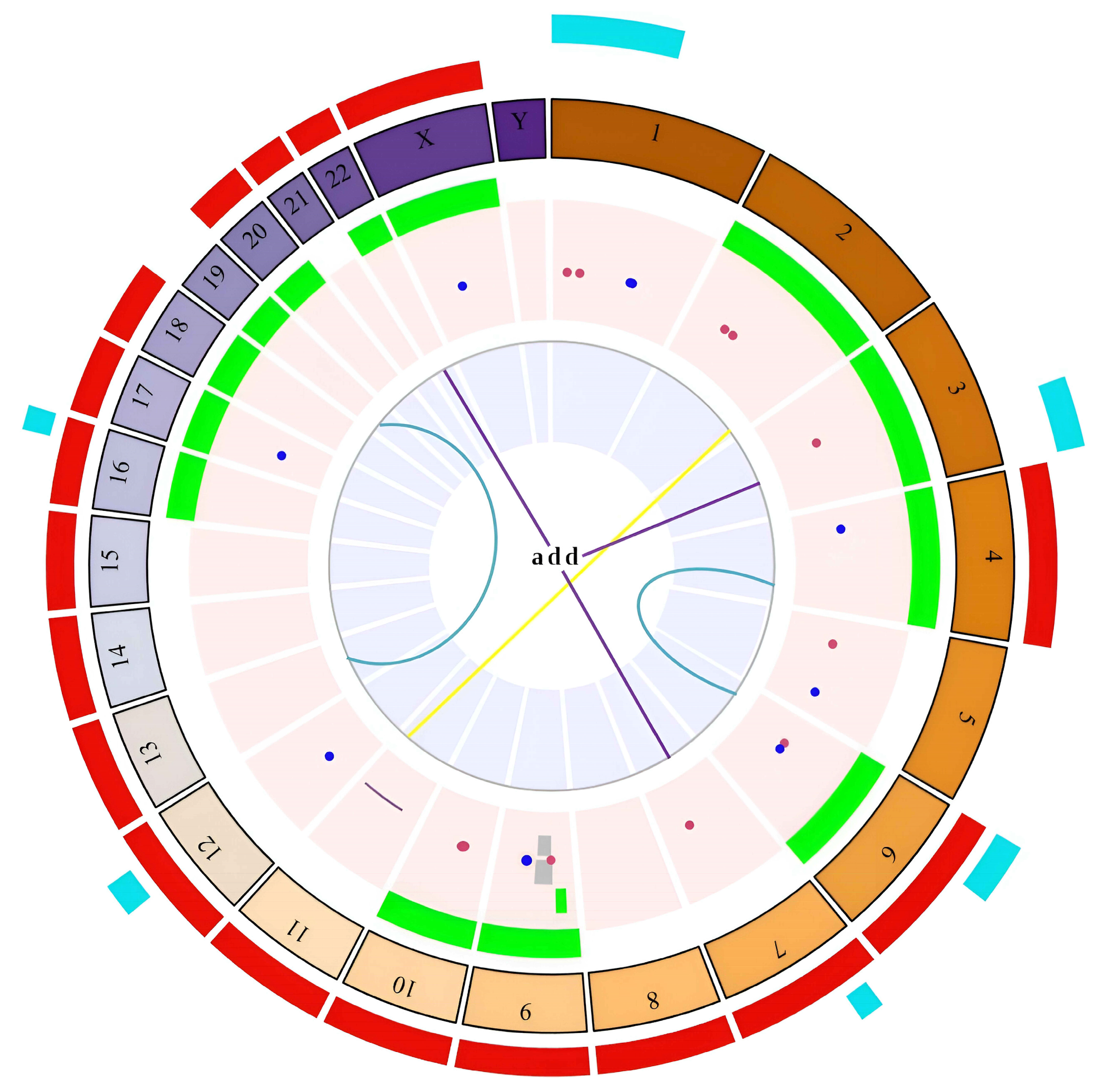
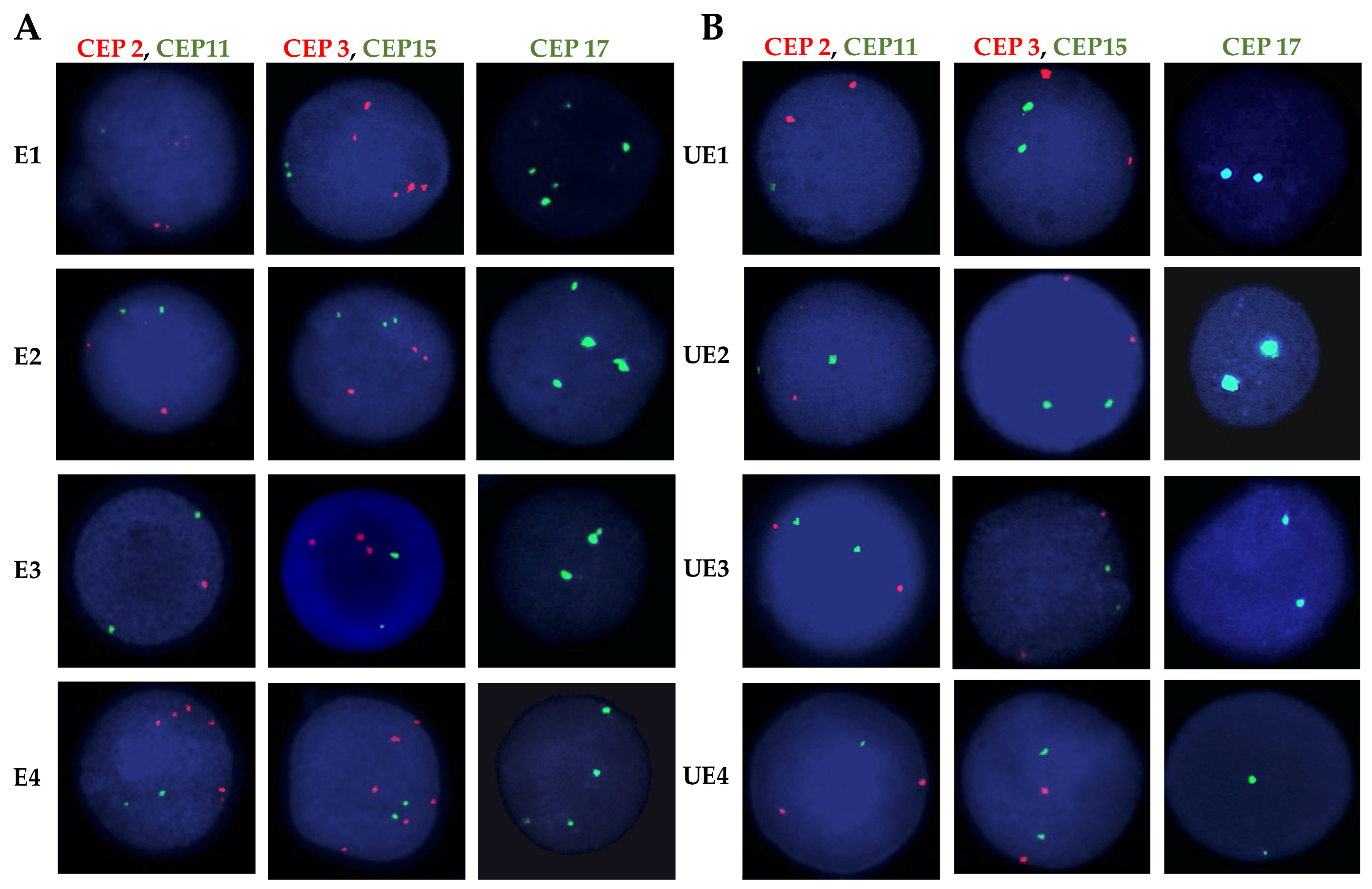
2.2. High Frequency of Chromosomal Alterations in Exposed Individuals
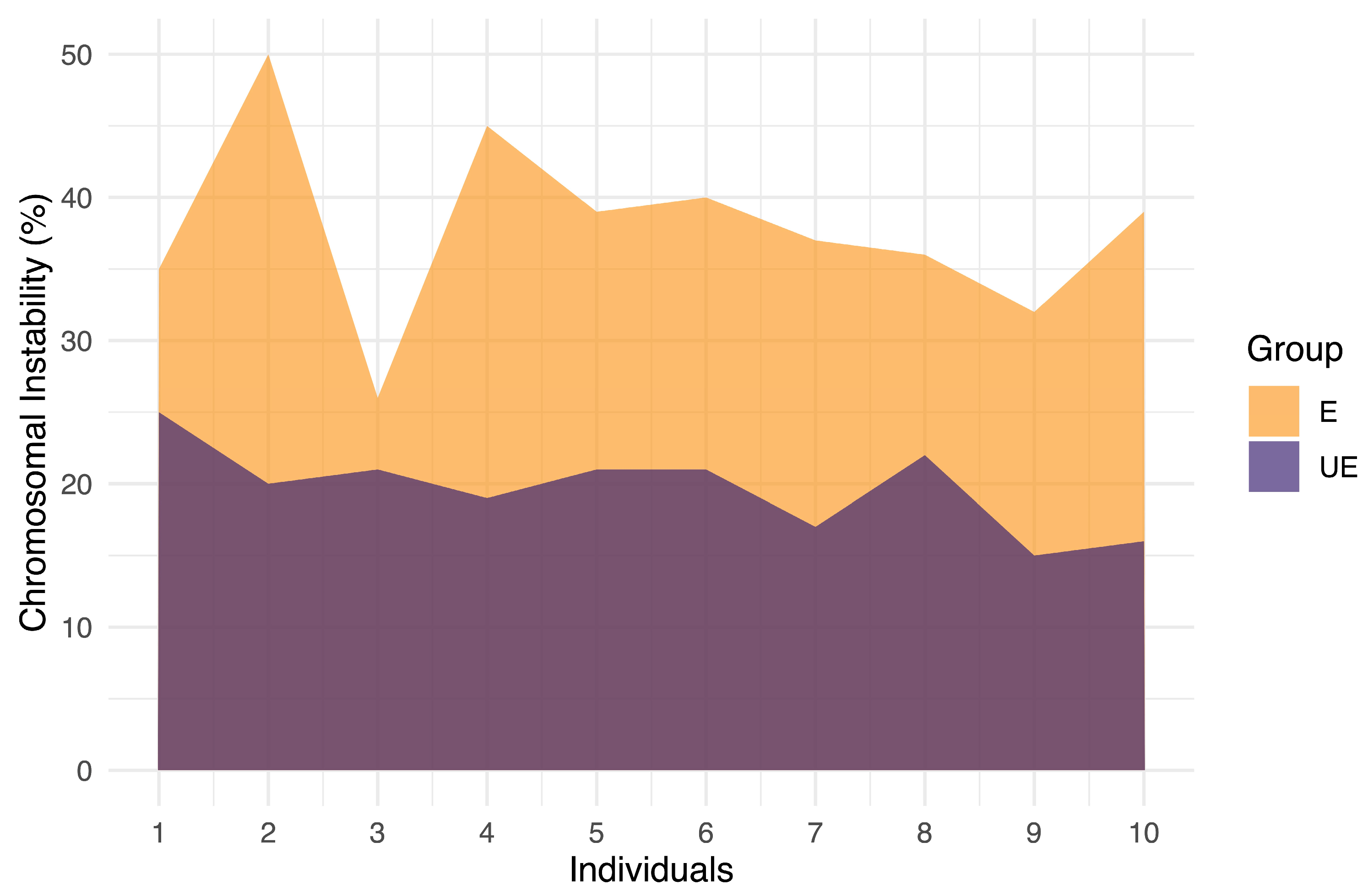
2.3. High Levels of Numerical Chromosomal Instability (CIN) in Exposed Individuals
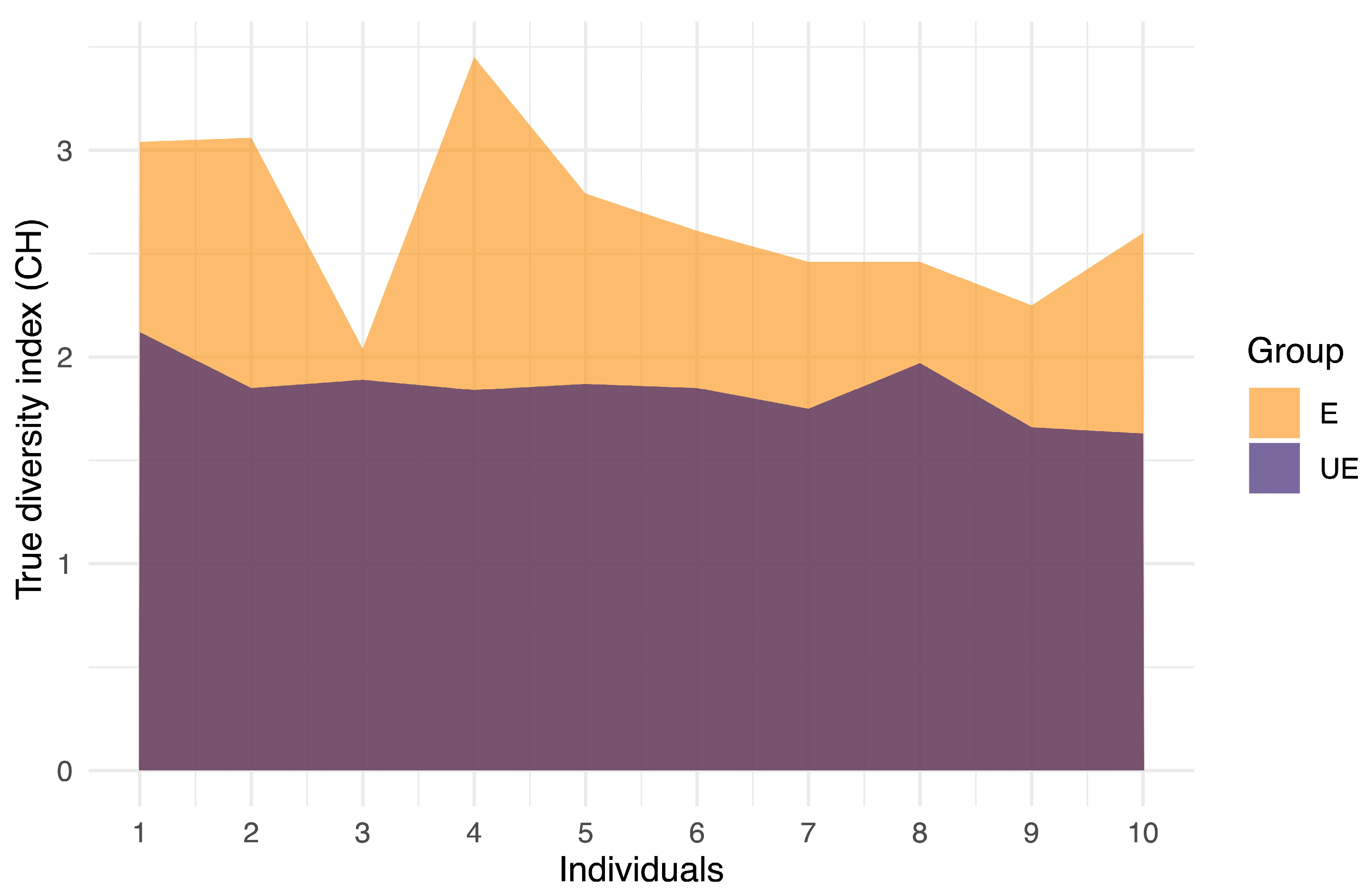
2.4. High CH in Exposed Individuals and Intermediate CH in Unexposed Individuals
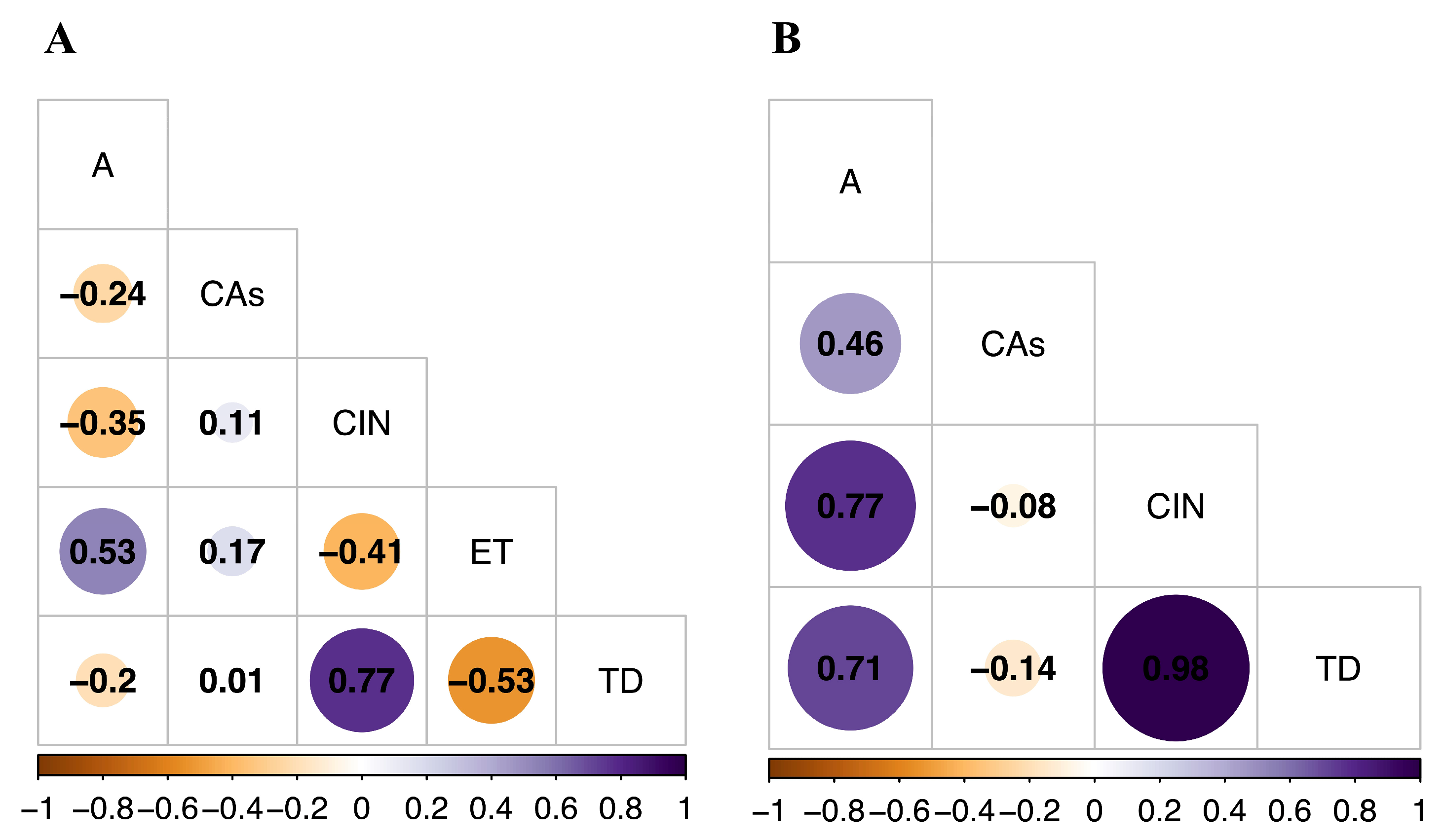
2.5. Association between Variables
2.6. Polymorphisms in the GSTP1 and XRCC1 Genes
3. Discussion
4. Materials and Methods
4.1. Study Groups
4.2. Blood Sample Collection
4.3. Cytogenetic Assays
4.4. Fluorescence In Situ Hybridization (FISH) Assays
4.5. CIN and CH Evaluation
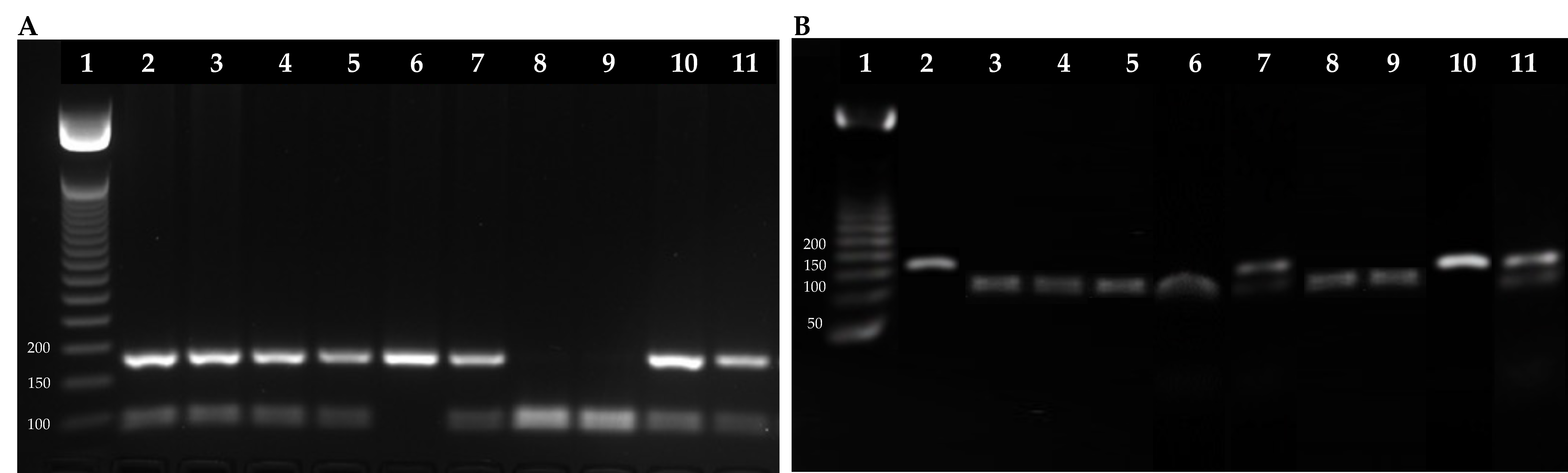
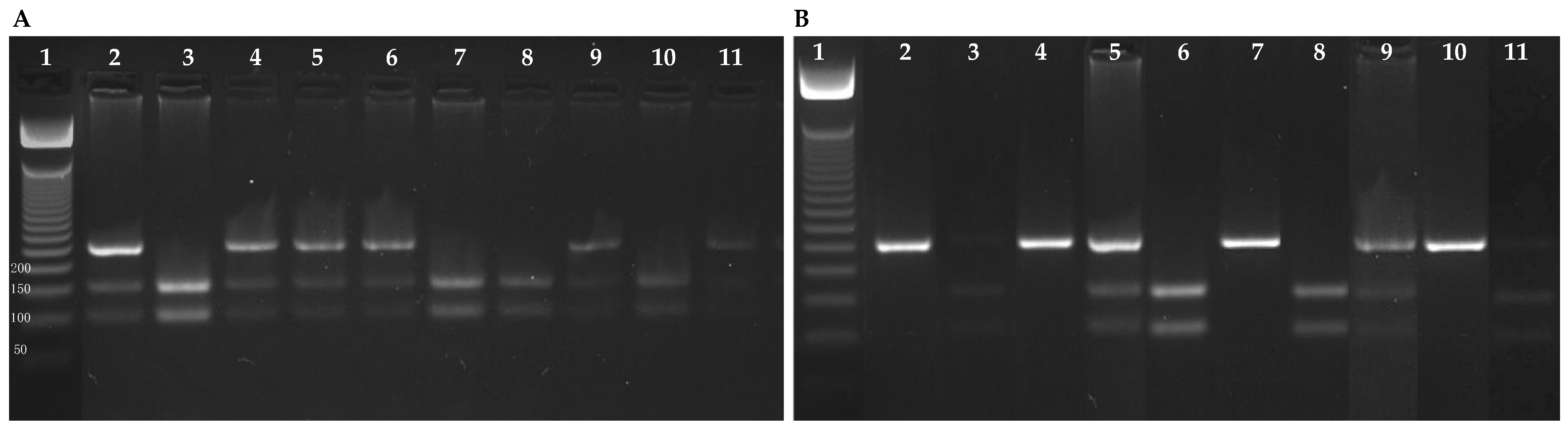
4.6. Polymerase Chain Reaction—Restriction Fragment Length Polymorphism (PCR-RFLP)
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balaji, M.; Sasikala, K. Cytogenetic effect of malathion in in vitro culture of human peripheral blood. Mutat. Res. Lett. 1993, 301, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Bréga, S.M.; Vassilieff, I.; Almeida, A.; Mercadante, A.; Bissacot, D.; Cury, P.R.; Freire-Maia, D.V. Cytogenetic and toxicological studies in rural workers exposed to pesticides in Botucatu, São Paulo, Brazil. Cad. Saúde Pública 1998, 14, 117–123. [Google Scholar] [CrossRef]
- Carbonell, E.; Xamena, N.; Creus, A.; Marcos, R. Cytogenetic biomonitoring in a Spanish group of agricultural workers exposed to pesticides. Mutagenesis 1993, 8, 511–517. [Google Scholar] [CrossRef] [PubMed]
- De Ferrari, M.; Artuso, M.; Bonassi, S.; Bonatti, S.; Cavalieri, Z.; Pescatore, D.; Marchini, E.; Pisano, V.; Abbondandolo, A. Cytogenetic biomonitoring of an Italian population exposed to pesticides: Chromosome aberration and sister-chromatid exchange analysis in peripheral blood lymphocytes. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 1991, 260, 105–113. [Google Scholar] [CrossRef]
- Dulout, F.; Pastori, M.; Olivero, O.; Cid, M.G.; Loria, D.; Matos, E.; Sobel, N.; de Bujan, E.; Albiano, N. Sister-chromatid exchanges and chromosomal aberrations in a population exposed to pesticides. Mutat. Res Lett. 1985, 143, 237–244. [Google Scholar] [CrossRef]
- Rupa, D.S.; Reddy, P.P.; Sreemannarayana, K.; Reddi, O.S.; Galloway, S.M. Frequency of sister chromatid exchange in peripheral lymphocytes of male pesticide applicators. Environ. Mol. Mutagen. 1991, 18, 136–138. [Google Scholar] [CrossRef]
- Carbonell, E.; Puig, M.; Xamena, N.; Creus, A.; Marcos, R. Sister chromatid exchange in lymphocytes of agricultural workers exposed to pesticides. Mutagénesis 1990, 5, 403–406. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Arroyo, S.; Noriega-Aldana, N.; Osorio, A.; Galicia, F.; Ling, S.; Villalobos-Pietrini, R. Sister-chromatid exchange analysis in a rural population of Mexico exposed to pesticides. Mutat. Res. Lett. 1992, 281, 173–179. [Google Scholar] [CrossRef]
- Prabhavathy Das, G.; Pasha Shaik, A.; Jamil, K. Cytotoxicity and genotoxicity induced by the pesticide profenofos on cultured human peripheral blood lymphocytes. Drug Chem. Toxicol. 2006, 29, 313–322. [Google Scholar] [CrossRef]
- Teodoro, M.; Briguglio, G.; Fenga, C.; Costa, C. Genetic polymorphisms as determinants of pesticide toxicity: Recent advances. Toxicol. Rep. 2019, 6, 564–570. [Google Scholar] [CrossRef]
- Wong, R.-H.; Chang, S.-Y.; Ho, S.-W.; Huang, P.-L.; Liu, Y.-J.; Chen, Y.-C.; Yeh, Y.-H.; Lee, H.-S. Polymorphisms in metabolic GSTP1 and DNA-repair XRCC1 genes with an increased risk of DNA damage in pesticide-exposed fruit growers. Mutat. Res. Toxicol. Environ. Mutagen. 2008, 654, 168–175. [Google Scholar] [CrossRef]
- Kapka-Skrzypczak, L.; Cyranka, M.; Skrzypczak, M.; Kruszewski, M. Biomonitoring and biomarkers of organophosphate pesticides exposure-state of the art. AAEM 2011, 18, 294–303. [Google Scholar] [PubMed]
- White, D.L.; Li, D.; Nurgalieva, Z.; El-Serag, H.B. Genetic variants of glutathione S-transferase as possible risk factors for hepatocellular carcinoma: A HuGE systematic review and meta-analysis. Ann. Epidemiol. 2008, 167, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kumar, V.; Singh, P.; Thakur, S.; Banerjee, B.D.; Rautela, R.S.; Grover, S.S.; Rawat, D.S.; Pasha, S.T.; Jain, S.K.; et al. Genetic polymorphisms of GSTM1, GSTT1 and GSTP1 and susceptibility to DNA damage in workers occupationally exposed to organophosphate pesticides. Mutat. ReGenet. Toxicol. Environ Mutagen. 2011, 725, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Cote, M.L.; Chen, W.; Smith, D.W.; Benhamou, S.; Bouchardy, C.; Butkiewicz, D.; Fong, K.M.; Gené, M.; Hirvonen, A.; Kiyohara, C.; et al. Meta-and pooled analysis of GSTP1 polymorphism and lung cancer: A HuGE-GSEC review. Am. J. Epidemiol. 2009, 169, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Al-Achkar, W.; Moassass, F.; Bassel, A.H.; Wafa, A. Glutathione S-transferase T1, M1 and P1 gene polymorphisms and susceptibility to colorectal cancer, a Syrian population study. Gene Rep. 2019, 15, 100365. [Google Scholar] [CrossRef]
- Galvez, H.R.C.; Flores, J.S.; Sanchez, E.D.T.; Bravo, D.R.; Villela, M.Z.R.; Uribe, E.R. Genetic profile for the detection of susceptibility to poisoning by exposure to pesticides. AAEM 2021, 28, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Otterlei, M.; Wong, H.K.; Tomkinson, A.E.; Wilson, D.M., III. XRCC1 co-localizes and physically interacts with PCNA. Nucleic Acids Res. 2004, 32, 2193–2201. [Google Scholar] [CrossRef]
- Nicolella, H.D.; de Assis, S. Epigenetic inheritance: Intergenerational effects of pesticides and other endocrine disruptors on cancer development. Int. J. Mol. Sci. 2022, 23, 4671. [Google Scholar] [CrossRef]
- Cavalier, H.; Trasande, L.; Porta, M. Exposures to pesticides and risk of cancer: Evaluation of recent epidemiological evidence in humans and paths forward. Int. J. Cancer 2023, 152, 879–912. [Google Scholar] [CrossRef]
- Ataei, M.; Abdollahi, M. A systematic review of mechanistic studies on the relationship between pesticide exposure and cancer induction. Toxicol. Appl. Pharmacol. 2022, 456, 116280. [Google Scholar] [CrossRef] [PubMed]
- Bolt, H.M.; Foth, H.; Hengstler, J.G.; Degen, G.H. Carcinogenicity categorization of chemicals—New aspects to be considered in a European perspective. Toxicol. Letts. 2004, 151, 29–41. [Google Scholar] [CrossRef]
- Gagos, S.; Irminger-Finger, I. Chromosome instability in neoplasia: Chaotic roots to continuous growth. Int. J. Biochem. Cell Biol. 2005, 37, 1014–1033. [Google Scholar] [CrossRef]
- Geigl, J.B.; Obenauf, A.C.; Schwarzbraun, T.; Speicher, M.R. Defining ‘chromosomal instability’. Trends Genet. 2008, 24, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Hirota, T. Chromosomal instability: A common feature and a therapeutic target of cancer. Biochim. Biophys. Acta Rev. Cancer 2016, 1866, 64–75. [Google Scholar] [CrossRef]
- Vargas-Rondón, N.; Villegas, V.E.; Rondón-Lagos, M. The role of chromosomal instability in cancer and therapeutic responses. Cancers 2017, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Durkin, S.G.; Glover, T.W. Chromosome fragile sites. Annu. Rev. Genet. 2007, 41, 169–192. [Google Scholar] [CrossRef]
- Hellman, A.; Zlotorynski, E.; Scherer, S.W.; Cheung, J.; Vincent, J.B.; Smith, D.I.; Trakhtenbrot, L.; Kerem, B. A role for common fragile site induction in amplification of human oncogenes. Cancer Cell 2002, 1, 89–97. [Google Scholar] [CrossRef]
- Arlt, M.F.; Durkin, S.G.; Ragland, R.L.; Glover, T.W. Common fragile sites as targets for chromosome rearrangements. DNA Repair 2006, 5, 1126–1135. [Google Scholar] [CrossRef]
- Debacker, K.; Kooy, R.F. Fragile sites and human disease. Hum. Mol. Genet. 2007, 16, 150–158. [Google Scholar] [CrossRef]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.J.R.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Børresen-Dale, A.-L.; et al. Signatures of mutational processes in human cancer. Nature 2013, 500, 415–421. [Google Scholar] [CrossRef]
- Kaur, R.; Kaur, M.; Singh, J. Endothelial dysfunction and platelet hyperactivity in type 2 diabetes mellitus: Molecular insights and therapeutic strategies. Cardiovasc. Diabetol. 2018, 17, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen-Pereira, C.H.; dos Santos, C.R.; Maraslis, F.T.; Pimentel, L.; Feijó, A.J.L.; Silva, C.I.; Medeiros, G.d.S.d.; Zeferino, R.C.; Pedrosa, R.C.; Maluf, S.W. Markers of genotoxicity and oxidative stress in farmers exposed to pesticides. Ecotoxicol. Environ. Saf. 2018, 148, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Shah, H.K.; Sharma, T.; Banerjee, B.D. Organochlorine pesticides induce inflammation, ROS production, and DNA damage in human epithelial ovary cells: An in vitro study. Chemosphere 2020, 246, 125691. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Wen, K.W.; Ki, A.; Prakash, S.; Kogan, S.; Yu, J. Genomic Features of Interstitial Deletions of Chromosome 9q in Acute Myeloid Leukemia. Cytogenet. Genome Res. 2022, 162, 119–123. [Google Scholar] [CrossRef]
- Deger, R.B.; Faruqi, S.A.; Noumoff, J.S. Karyotypic analysis of 32 malignant epithelial ovarian tumors. Cancer Genet. Cytogen 1997, 96, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.R.; Aguilera, N.I.; Reshmi, S.; Huang, X.; Yu, Z.; Gollin, S.M.; Abbondanzo, S.L.; Swerdlow, S.H. Deletion 6q is not a characteristic marker of nodal lymphoplasmacytic lymphoma. Cancer Genet Cytogen. 2005, 162, 85–88. [Google Scholar] [CrossRef]
- Sawyer, J.R.; Tian, E.; Heuck, C.J.; Epstein, J.; Johann, D.J.; Swanson, C.M.; Lukacs, J.L.; Johnson, M.; Binz, R.; Boast, A.; et al. Jumping translocations of 1q12 in multiple myeloma: A novel mechanism for deletion of 17p in cytogenetically defined high-risk disease. Blood 2014, 123, 2504–2512. [Google Scholar] [CrossRef]
- Thompson, S.L.; Bakhoum, S.F.; Compton, D.A. Mechanisms of chromosomal instability. Curr. Biol. 2010, 20, 285–295. [Google Scholar] [CrossRef]
- Heng, H.H.; Bremer, S.W.; Stevens, J.B.; Horne, S.D.; Liu, G.; Abdallah, B.Y.; Ye, K.J.; Ye, C.J. Chromosomal instability (CIN): What it is and why it is crucial to cancer evolution. Cancer Metastasis Rev. 2013, 32, 325–340. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, F.; Ou, J. Global pesticide consumption and pollution: With China as a focus. Proc. Int. Acad. 2011, 1, 125. [Google Scholar]
- Cepeda, S.; Forero-Castro, M.; Cárdenas-Nieto, D.; Martínez-Agüero, M.; Rondón-Lagos, M. Chromosomal instability in farmers exposed to pesticides: High prevalence of clonal and non-clonal chromosomal alterations. Risk Manag. Healthc. Policy 2020, 13, 97–110. [Google Scholar] [CrossRef]
- Dayal, J.H.; Albergante, L.; Newman, T.J.; South, A.P. Quantitation of multiclonality in control and drug-treated tumour populations using high-throughput analysis of karyotypic heterogeneity. Converg. Sci. Phys. Oncol. 2015, 1, 025001. [Google Scholar] [CrossRef]
- Fan, X.; Paetau, A.; Aalto, Y.; Välimäki, M.; Sane, T.; Poranen, A.; Castresana, J.S.; Knuutila, S. Gain of chromosome 3 and loss of 13q are frequent alterations in pituitary adenomas. Cancer Genet. Cytogenet. 2001, 128, 97–103. [Google Scholar] [CrossRef]
- Amato, E.; Barbi, S.; Malpeli, G.; Bersani, S.; Pelosi, G.; Capelli, P.; Scarpa, A. Chromosome 3p alterations in pancreatic endocrine neoplasia. Virchows Archiv. 2011, 458, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Yagyu, T.; Ohira, T.; Shimizu, R.; Morimoto, M.; Murakami, Y.; Hanaki, T.; Kihara, K.; Matsunaga, T.; Yamamoto, M.; Tokuyasu, N.; et al. Human chromosome 3p21. 3 carries TERT transcriptional regulators in pancreatic cancer. Sci. Rep. 2021, 11, 15355. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, B.; Pothof, J.; Vijg, J.; Hoeijmakers, J.H. The central role of DNA damage in the ageing process. Nature 2021, 592, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, T. Instability and decay of the primary structure of DNA. Nature 1993, 362, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Vijg, J. Aging of the Genome: The Dual Role of DNA in Life and Death; OUP: Oxford, UK, 2007. [Google Scholar] [CrossRef]
- Gutiérrez, J.B.; de Cerain Salsamendi, A.L. Fundamentos de Ciencia Toxicológica. 2001. Available online: http://www.diazdesantos.es (accessed on 10 February 2021).
- Frank, P.; Ottoboni, M.A. The Dose Makes the Poison: A Plain-Language Guide to Toxicology; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar] [CrossRef]
- Kokoszka, J.E.; Coskun, P.; Esposito, L.A.; Wallace, D.C. Increased mitochondrial oxidative stress in the Sod2 (+/−) mouse results in the age-related decline of mitochondrial function culminating in increased apoptosis. PNAS 2001, 98, 2278–2283. [Google Scholar] [CrossRef] [PubMed]
- Surniyantoro, H.N.E.; Yusuf, D.; Rahardjo, T.; Rahajeng, N.; Kisnanto, T.; Nurhayati, S.; Lusiyanti, Y.; Syaifudin, M.; Hande, M.P. Assessment of hOGG1 Genetic Polymorphism (rs1052133) and DNA Damage in Radiation-Exposed Workers. APJCP 2022, 23, 4005. [Google Scholar] [CrossRef] [PubMed]
- Zijno, A.; Marcon, F.; Leopardi, P.; Crebelli, R. Analysis of chromosome segregation in cytokinesis-blocked human lymphocytes: Non-disjunction is the prevalent damage resulting from low dose exposure to spindle poisons. Mutagenesis 1996, 11, 335–340. [Google Scholar] [CrossRef]
- Lushchak, V.I.; Matviishyn, T.M.; Husak, V.V.; Storey, J.M.; Storey, K.B. Pesticide toxicity: A mechanistic approach. EXCLI J. 2018, 17, 1101. [Google Scholar] [CrossRef] [PubMed]
- Parry, E.; Parry, J.; Corso, C.; Doherty, A.; Haddad, F.; Hermine, T.; Johnson, G.; Kayani, M.; Quick, E.; Warr, T.; et al. Detection and characterization of mechanisms of action of aneugenic chemicals. Mutagenesis 2002, 17, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Renzi, L.; Pacchierotti, F.; Russo, A. The centromere as a target for the induction of chromosome damage in resting and proliferating mammalian cells: Assessment of mitomycin C-induced genetic damage at kinetochores and centromeres by a micronucleus test in mouse splenocytes. Mutagenesis 1996, 11, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Mattiuzzo, M.; Fiore, M.; Ricordy, R.; Degrassi, F. Aneuploidy-inducing capacity of two widely used pesticides. Carcinog 2006, 27, 2511–2518. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-M.; Wu, M.-T.; Lee, Y.-C.; Yang, S.-Y.; Chen, J.-S.; Hsu, H.-H.; Huang, P.-M.; Kuo, S.-W.; Lee, C.-J.; Chen, C.-J. Association of GSTP1 polymorphism and survival for esophageal cancer. Clin. Cancer Res. 2005, 11, 4749–4753. [Google Scholar] [CrossRef]
- da Silva, M.C.; de Oliveira, R.A.A.; de Vasconcellos, A.C.S.; Rebouças, B.H.; Pinto, B.D.; Lima, M.d.O.; de Jesus, I.M.; Machado, D.E.; Hacon, S.S.; Basta, P.C.; et al. Chronic Mercury Exposure and GSTP1 Polymorphism in Munduruku Indigenous from Brazilian Amazon. Toxics 2023, 11, 138. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Du, Y.; Zhou, Q.; Ma, J.; Yu, J.; Tao, X.; Zhang, F. Association of GSTP1 Ile105Val polymorphism with risk of esophageal cancer: A meta-analysis of 21 case-control studies. Int. J. Clin. Exp. Med. 2014, 7, 3215. [Google Scholar] [PubMed]
- Alves, A.A.; Laurinho, K.; Franco, F.C.; de Araujo Nascimento, F.; Nunes, H.F.; de Melo e Silva, D. The incidence of the XRCC1 rs25487 and PON1 rs662 polymorphisms in a population from Central Brazil: Patterns in an area with a high level of agricultural activity. Biochem. Genet. 2023, 61, 1675–1703. [Google Scholar] [CrossRef] [PubMed]
- Saad-Hussein, A.; Noshy, M.; Taha, M.; El-Shorbagy, H.; Shahy, E.; Abdel-Shafy, E.A. GSTP1 and XRCC1 polymorphisms and DNA damage in agricultural workers exposed to pesticides. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2017, 819, 20–25. [Google Scholar] [CrossRef]
- Sabarwal, A.; Kumar, K.; Singh, R.P. Hazardous effects of chemical pesticides on human health–Cancer and other associated disorders. Environ. Toxicol. Pharmacol. 2018, 63, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.; Banerjee, B.D.; Thakur, G.K.; Guleria, K.; Mazumdar, D. Polymorphism of xenobiotic metabolizing gene and susceptibility of epithelial ovarian cancer with reference to organochlorine pesticides exposure. Exp. Biol. Med. 2019, 244, 1446–1453. [Google Scholar] [CrossRef] [PubMed]
- Ahluwalia, M.; Kaur, A. Modulatory role of GSTT1 and GSTM1 in Punjabi agricultural workers exposed to pesticides. Environ. Sci. Pollut. Res. 2018, 25, 11981–11986. [Google Scholar] [CrossRef]
- Saarikoski, S.T.; Voho, A.; Reinikainen, M.; Anttila, S.; Karjalainen, A.; Malaveille, C.; Vainio, H.; Husgafvel-Pursiainen, K.; Hirvonen, A. Combined effect of polymorphic GST genes on individual susceptibility to lung cancer. Int. J. Cancer 1998, 77, 516–521. [Google Scholar] [CrossRef]
- Furlong, C.E.; Li, W.F.; Brophy, V.H.; Jarvik, G.P.; Richter, R.J.; Shih, D.M.; Lusis, A.J.; Costa, L.G. The PON1 gene and detoxication. Neurotoxicology 2000, 21, 581–587. [Google Scholar] [PubMed]
- Chambers, J.E.; Meek, E.C.; Chambers, H.W. The metabolism of organophosphorus insecticides. In Hayes’ Handbook of Pesticide Toxicology; Academic Press: Cambridge, MA, USA, 2010; pp. 1399–1407. [Google Scholar] [CrossRef]
- Abou-Donia, M. Mammalian Toxicology; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Qadri, Q.; Sameer, A.S.; Shah, Z.A.; Hamid, A.; Alam, S.; Manzoor, S.; Siddiqi, M.A. Genetic polymorphism of the glutathione-S-transferase P1 gene (GSTP1) and susceptibility to prostate cancer in the Kashmiri population. Genet. Mol. Res. 2011, 10, 3038–3045. [Google Scholar] [CrossRef] [PubMed]
- Tumer, T.B.; Savranoglu, S.; Atmaca, P.; Terzioglu, G.; Sen, A.; Arslan, S. Modulatory role of GSTM1 null genotype on the frequency of micronuclei in pesticide-exposed agricultural workers. Toxicol. Ind. Health 2016, 32, 1942–1951. [Google Scholar] [CrossRef]
- McGowan-Jordan, J.; Hastings, R.J.; Moore, S. ISCN 2020: An international system for human cytogenomic nomenclature (2020). Cytogenet. Genome Res. 2020, 161, 225–226. [Google Scholar] [CrossRef]
- Lengauer, C.; Kinzler, K.W.; Vogelstein, B. Genetic instability in colorectal cancers. Nature 1997, 386, 623–627. [Google Scholar] [CrossRef]
- Munro, A.F.; Twelves, C.; Thomas, J.S.; Cameron, D.A.; Bartlett, J.M.S. Chromosome instability and benefit from adjuvant anthracyclines in breast cancer. Br. J. Cancer 2012, 107, 71–74. [Google Scholar] [CrossRef]
- Tsuji, K.; Kawauchi, S.; Saito, S.; Furuya, T.; Ikemoto, K.; Nakao, M.; Yamamoto, S.; Oka, M.; Hirano, T.; Sasaki, K. Breast cancer cell lines carry cell line-specific genomic alterations that are distinct from aberrations in breast cancer tissues: Comparison of the CGH profiles between cancer cell lines and primary cancer tissues. BMC Cancer 2010, 10, 15. [Google Scholar] [CrossRef] [PubMed]
- Talamo, A.; Chalandon, Y.; Marazzi, A.; Jotterand, M. Clonal heterogeneity and chromosomal instability at disease presentation in high hyperdiploid acute lymphoblastic leukemia. Cancer Genet Cytogenet. 2010, 203, 209–214. [Google Scholar] [CrossRef]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Roylance, R.; Endesfelder, D.; Gorman, P.; Burrell, R.A.; Sander, J.; Tomlinson, I.; Hanby, A.M.; Speirs, V.; Richardson, A.L.; Birkbak, N.J.; et al. Relationship of extreme chromosomal instability with long-term survival in a retrospective analysis of primary breast cancer. CEBP 2011, 20, 2183–2194. [Google Scholar] [CrossRef] [PubMed]
- Maley, C.C.; Galipeau, P.C.; Finley, J.C.; Wongsurawat, V.J.; Li, X.; Sanchez, C.A.; Paulson, T.G.; Blount, P.L.; Risques, R.-A.; Rabinovitch, P.S.; et al. Genetic clonal diversity predicts progression to esophageal adenocarcinoma. Nat. Genet. 2006, 38, 468–473. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposed | Unexposed | |
|---|---|---|
| Number | 10 | 10 |
| Age (mean ± SD) | 39.20 ± 17.78 | 39.00 ± 17.44 |
| Sex (n) | ||
| Male | 3 | 3 |
| Female | 7 | 7 |
| Exposure months (mean ± SD) | 180.2 ± 239.9 | 0 |
| Tobacco smoking (n) | ||
| Smokers | 0 | 0 |
| Non-smokers | 10 | 10 |
| Alcohol consumption (n) | ||
| Alcohol consumers (1/week) | 2 | 1 |
| Alcohol consumers (sporadic) | 7 | 8 |
| Non-alcohol consumers | 1 | 1 |
| Characteristic | Number |
|---|---|
| Type of exposure | |
| Dermal | 7 |
| Respiratory | 3 |
| Method of irrigation | |
| Machine | 7 |
| Bomb | 2 |
| Mixed | 1 |
| Frequency of irrigation | |
| 2 times a month | 4 |
| 1 time a week | 4 |
| Unknown | 2 |
| CAs and CVs | Number of Alterations | ||
|---|---|---|---|
| E | UE | p+ | |
| n (%) | n (%) | ||
| NCAs | 151 (43.0) | 33 (8.7) | 0.0001 ** |
| Monosomies | 42 (12.0) | 21 (5.5) | 0.2159 |
| Trisomies | 29 (8.3) | 2 (0.5) | 0.0349 * |
| mar | 38 (10.8) | 4 (1.1) | 0.0050 ** |
| end | 21 (6.0) | 2 (0.5) | 0.1184 |
| Polyploidies | 20 (5.7) | 3 (0.8) | 0.1184 |
| SCAs | 20 (5.7) | 15 (3.9) | 0.7475 |
| chtb/chrb | 20 (5.7) | 7 (1.8) | 0.2790 |
| fra | 42 (12.0) | 22 (5.8) | 0.2159 |
| fra(9)(q12) | 29 (8.3) | 16 (4.2) | 0.3727 |
| 9qh+ | 4 (1.1) | 5 (1.3) | 1 |
| Total alterations | 237 | 82 | |
| Mean | 47.4 | 16.4 | |
| SD | 59.4 | 11.5 | |
| p++ | 0.125 | ||
| CVs and CAs | Number of Individuals | ||
|---|---|---|---|
| E | UE | p | |
| n (%) | n (%) | ||
| NCAs | 10 (100) | 8 (80) | 0.0001 ** |
| Monosomies | 10 (100) | 8 (80) | 0.0001 ** |
| Trisomies | 10 (100) | 3 (30) | 0.0001 ** |
| mar | 9 (90) | 3 (30) | 0.0001 ** |
| end | 5 (50) | 1 (10) | 0.0001 ** |
| Polyploidies | 6 (60) | 2 (20) | 0.0001 ** |
| SCAs | 9 (90) | 7 (70) | 0.0007 ** |
| chtb/chrb | 6 (60) | 4 (40) | 0.0071 ** |
| fra | 8 (80) | 7 (70) | 0.1412 |
| 9qh+ | 2 (20) | 2 (20) | 1 |
| Total | 10 | 10 | |
| Gene | SNP Genotype | Exposed | Unexposed | p |
|---|---|---|---|---|
| GSTP1 Exon 5 | Ile105Val (A→G) | |||
| AA | 1 (10%) | 2 (20%) | 0.0734 | |
| AG | 7 (70%) | 2 (20%) | 0.0001 ** | |
| GG | 2 (20%) | 6 (60%) | 0.0001 ** | |
| XRCC1 Exon 6 | Arg194Trp (C→T) | |||
| CC | 8 (80%) | 8 (80%) | 1 | |
| CT | 2 (20%) | 2 (20%) | 1 | |
| TT | 0 | 0 | 1 | |
| XRCC1 Exon 10 | Arg399Gln (G→A) | |||
| GG | 0 | 4 (40%) | 0.0001 ** | |
| GA | 6 (60%) | 4 (40%) | 0.0071 ** | |
| AA | 4 (40%) | 2 (20%) | 0.0032 ** | |
| Gene | Primer | Primer Sequences | Exon | PCR Product | Restriction Enzyme |
|---|---|---|---|---|---|
| GSTP1 | Ile105Val (A→G) | F: 5′ACCCCAGGGCTCTATGGGAA3′ | 5 | 176 bp | BsmA1 |
| R: 5′TGAGGGCACAAGAAGCCCCT3′ | |||||
| XRCC1 | Arg194Trp (C→T) | F: 5′GCCAGGGCCCCTCCTTCAA3′ | 6 | 485 bp | Pvu II |
| R: 5′TACCCTCAGACCCACGAGT3′ | |||||
| Arg399Gln (G→A) | F: 5′CCCCAAGTACAGCCAGGTC3′ | 10 | 242 bp | MspI | |
| R: 5′TGTCCCGCTCCTCTCAGTAG3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aldana-Salazar, F.; Rangel, N.; Rodríguez, M.J.; Baracaldo, C.; Martínez-Agüero, M.; Rondón-Lagos, M. Chromosomal Damage, Chromosome Instability, and Polymorphisms in GSTP1 and XRCC1 as Biomarkers of Effect and Susceptibility in Farmers Exposed to Pesticides. Int. J. Mol. Sci. 2024, 25, 4167. https://doi.org/10.3390/ijms25084167
Aldana-Salazar F, Rangel N, Rodríguez MJ, Baracaldo C, Martínez-Agüero M, Rondón-Lagos M. Chromosomal Damage, Chromosome Instability, and Polymorphisms in GSTP1 and XRCC1 as Biomarkers of Effect and Susceptibility in Farmers Exposed to Pesticides. International Journal of Molecular Sciences. 2024; 25(8):4167. https://doi.org/10.3390/ijms25084167
Chicago/Turabian StyleAldana-Salazar, Fernando, Nelson Rangel, María José Rodríguez, César Baracaldo, María Martínez-Agüero, and Milena Rondón-Lagos. 2024. "Chromosomal Damage, Chromosome Instability, and Polymorphisms in GSTP1 and XRCC1 as Biomarkers of Effect and Susceptibility in Farmers Exposed to Pesticides" International Journal of Molecular Sciences 25, no. 8: 4167. https://doi.org/10.3390/ijms25084167