
Two-Month Voluntary Ethanol Consumption Promotes Mild Neuroinflammation in the Cerebellum but Not in the Prefrontal Cortex, Hippocampus, or Striatum of Mice

 ,
,  and
and 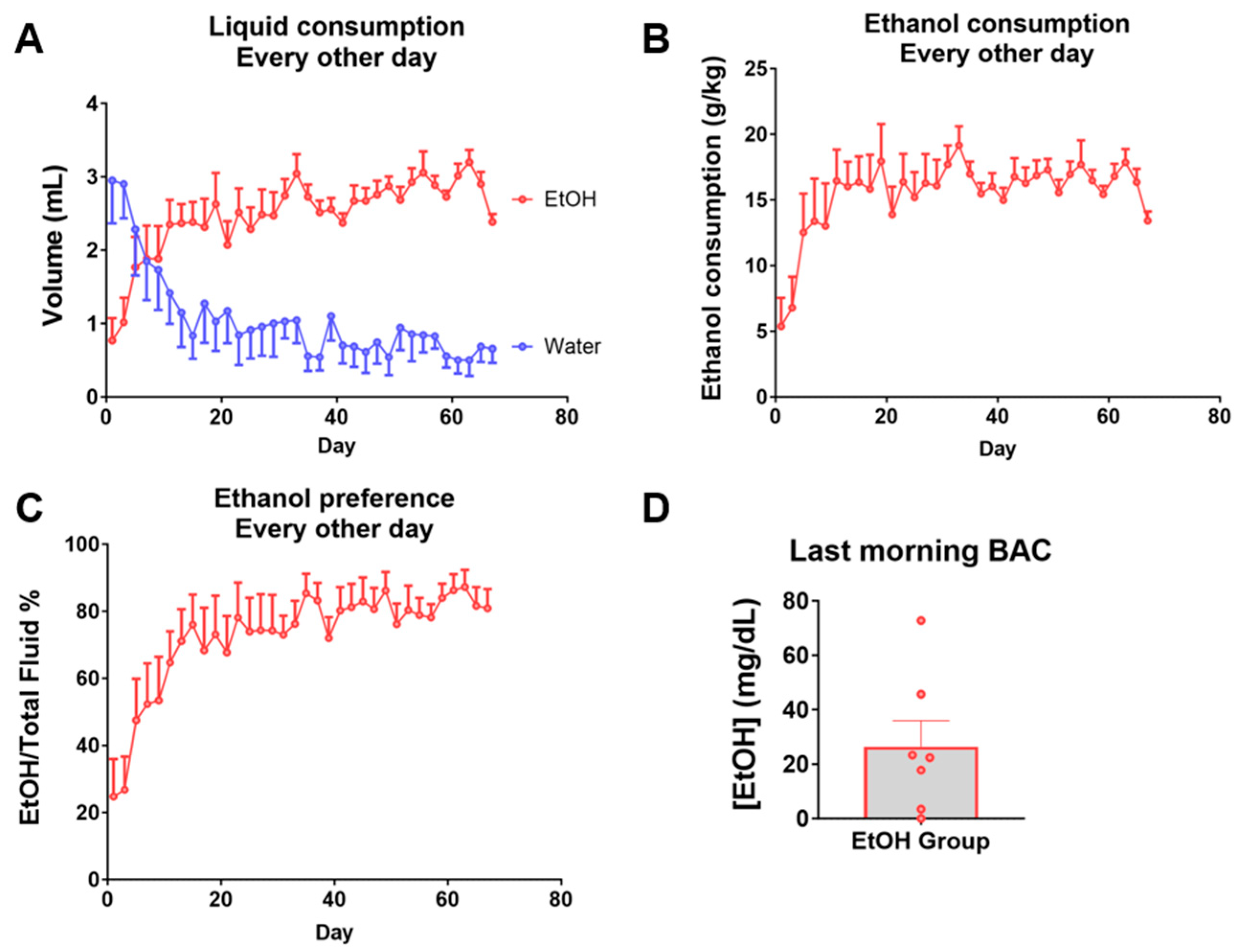{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Intermittent Chronic Voluntary Ethanol Consumption Model in C57BL/6 Female Mice
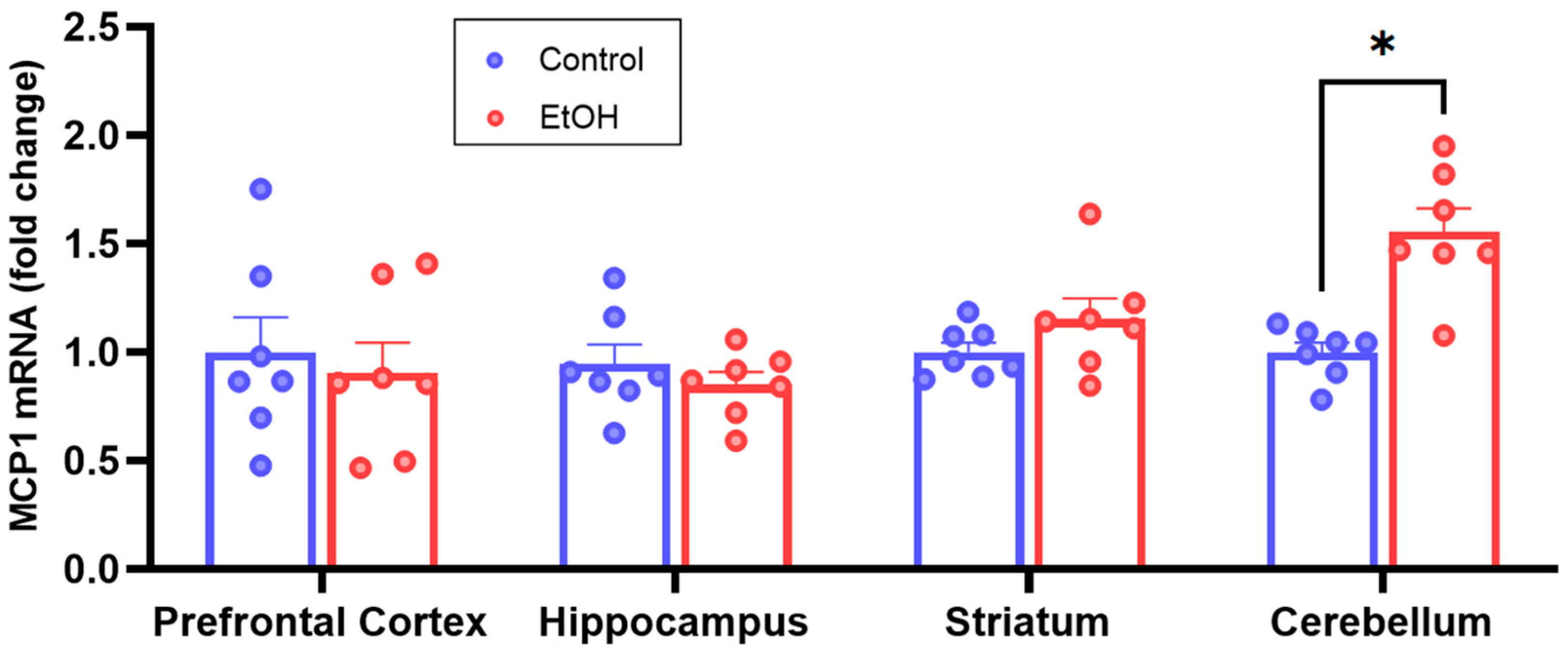
2.2. RT-qPCR MCP1 Analyses
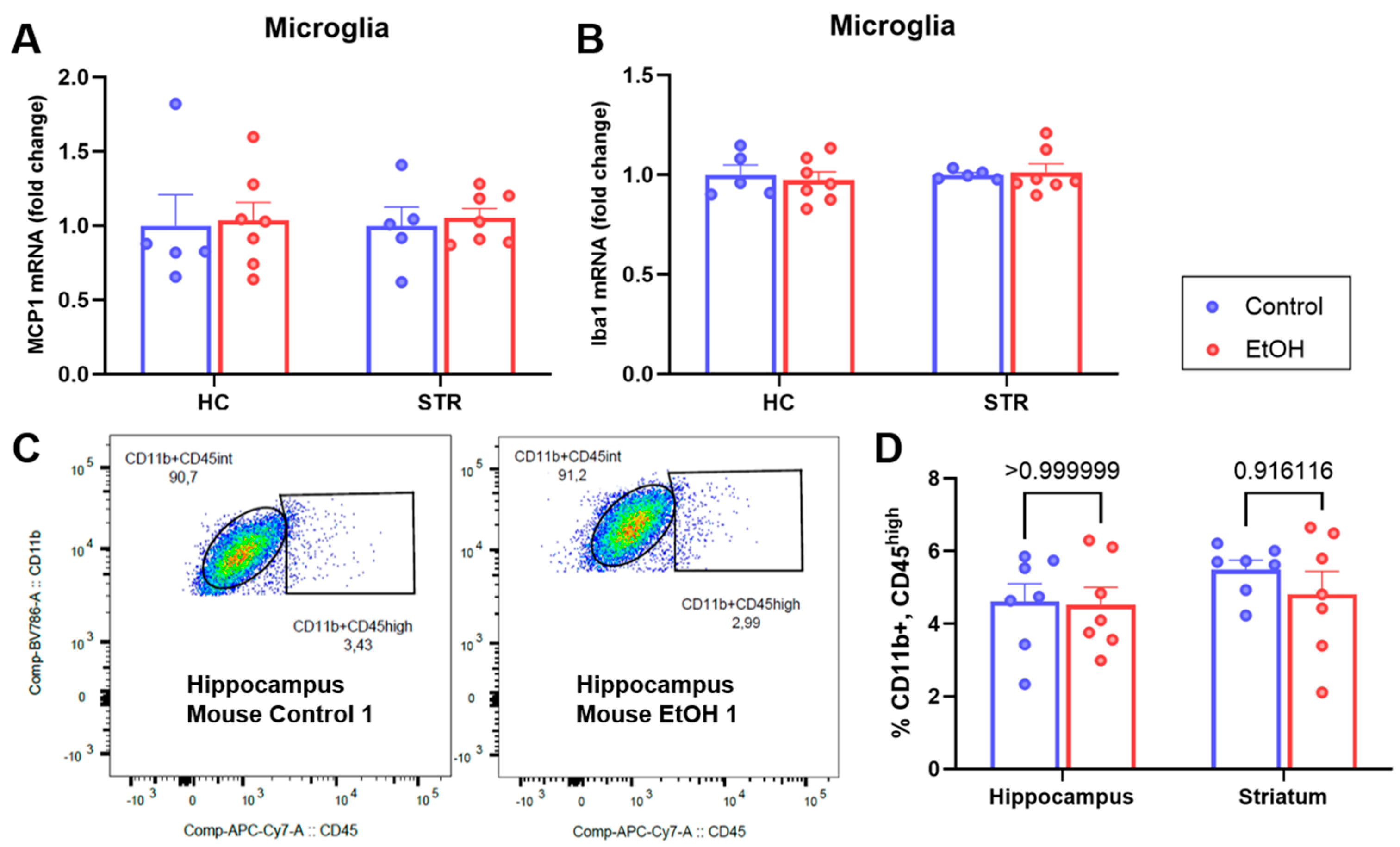
2.3. Analyses of Isolated Microglia
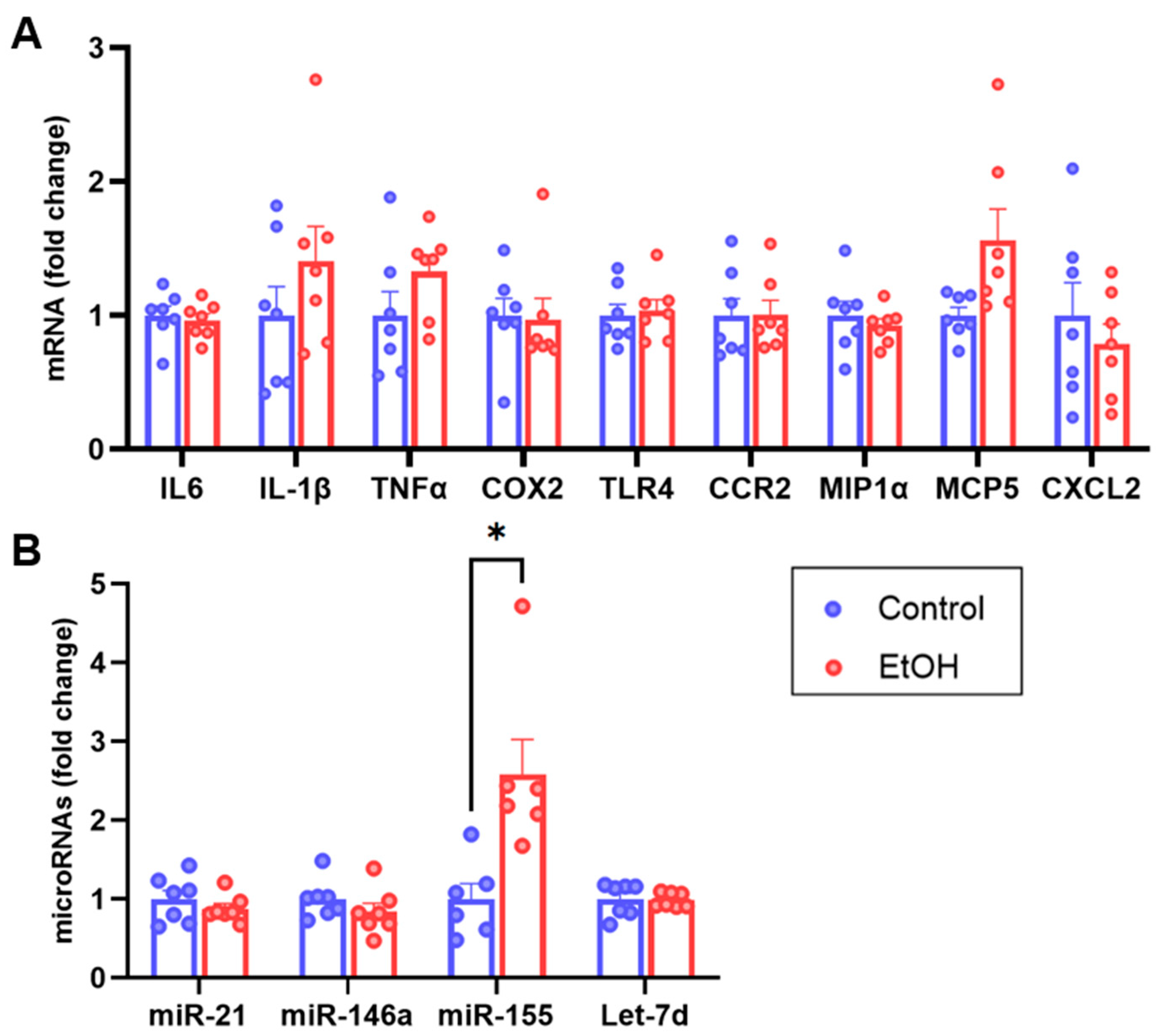
2.4. Cerebellum RT-qPCR Characterization
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Ethanol Voluntary Consumption Design
4.3. Euthanasia
4.4. Microglia Purification by FACS and RNA Extraction
4.5. Tissue Homogenization and RNA Extraction
4.6. cDNA Synthesis for mRNA Determination by RT-qPCR
4.7. miQPCR Method for Determination of microRNA Levels by RT-qPCR
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Global Status Report on Alcohol and Health 2018; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Degenhardt, L.; Charlson, F.; Ferrari, A.; Santomauro, D.; Erskine, H.; Mantilla-Herrara, A.; Whiteford, H.; Leung, J.; Naghavi, M.; Griswold, M. The global burden of disease attributable to alcohol and drug use in 195 countries and territories, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Psychiatry 2018, 5, 987–1012. [Google Scholar] [CrossRef] [PubMed]
- Grant, B.F.; Goldstein, R.B.; Saha, T.D.; Chou, S.P.; Jung, J.; Zhang, H.; Pickering, R.P.; Ruan, W.J.; Smith, S.M.; Huang, B.; et al. Epidemiology of DSM-5 Alcohol Use Disorder: Results From the National Epidemiologic Survey on Alcohol and Related Conditions III. JAMA Psychiatry 2015, 72, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Ray, L.A.; Roche, D.J.; Heinzerling, K.; Shoptaw, S. Opportunities for the development of neuroimmune therapies in addiction. Int. Rev. Neurobiol. 2014, 118, 381–401. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.; Most, D.; Ferguson, L.B.; Mayfield, J.; Harris, R.A.; Blednov, Y.A. Neuroimmune pathways in alcohol consumption: Evidence from behavioral and genetic studies in rodents and humans. Int. Rev. Neurobiol. 2014, 118, 13–39. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, J.; Harris, R.A. The Neuroimmune Basis of Excessive Alcohol Consumption. Neuropsychopharmacology 2017, 42, 376. [Google Scholar] [CrossRef] [PubMed]
- Berrios-Carcamo, P.; Quezada, M.; Quintanilla, M.E.; Morales, P.; Ezquer, M.; Herrera-Marschitz, M.; Israel, Y.; Ezquer, F. Oxidative Stress and Neuroinflammation as a Pivot in Drug Abuse. A Focus on the Therapeutic Potential of Antioxidant and Anti-Inflammatory Agents and Biomolecules. Antioxidants 2020, 9, 830. [Google Scholar] [CrossRef] [PubMed]
- Meredith, L.R.; Burnette, E.M.; Grodin, E.N.; Irwin, M.R.; Ray, L.A. Immune treatments for alcohol use disorder: A translational framework. Brain Behav. Immun. 2021, 97, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Pascual, M.; Calvo-Rodriguez, M.; Nunez, L.; Villalobos, C.; Urena, J.; Guerri, C. Toll-like receptors in neuroinflammation, neurodegeneration, and alcohol-induced brain damage. IUBMB Life 2021, 73, 900–915. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Ohsawa, K.; Kanazawa, H.; Kohsaka, S.; Imai, Y. Iba1 is an actin-cross-linking protein in macrophages/microglia. Biochem. Biophys. Res. Commun. 2001, 286, 292–297. [Google Scholar] [CrossRef]
- Borst, K.; Dumas, A.A.; Prinz, M. Microglia: Immune and non-immune functions. Immunity 2021, 54, 2194–2208. [Google Scholar] [CrossRef]
- Wise, R.A. Dopamine, learning and motivation. Nat. Rev. Neurosci. 2004, 5, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.A. Roles for nigrostriatal—Not just mesocorticolimbic—Dopamine in reward and addiction. Trends Neurosci. 2009, 32, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Kutlu, M.G.; Gould, T.J. Effects of drugs of abuse on hippocampal plasticity and hippocampus-dependent learning and memory: Contributions to development and maintenance of addiction. Learn. Mem. 2016, 23, 515–533. [Google Scholar] [CrossRef] [PubMed]
- Conductier, G.; Blondeau, N.; Guyon, A.; Nahon, J.L.; Rovere, C. The role of monocyte chemoattractant protein MCP1/CCL2 in neuroinflammatory diseases. J. Neuroimmunol. 2010, 224, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Melbourne, J.K.; Chandler, C.M.; Van Doorn, C.E.; Bardo, M.T.; Pauly, J.R.; Peng, H.; Nixon, K. Primed for addiction: A critical review of the role of microglia in the neurodevelopmental consequences of adolescent alcohol drinking. Alcohol. Clin. Exp. Res. 2021, 45, 1908–1926. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Luo, J. Role of MCP-1 and CCR2 in alcohol neurotoxicity. Pharmacol. Res. 2019, 139, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; He, J.; Hanes, R.N.; Pluzarev, O.; Hong, J.S.; Crews, F.T. Increased systemic and brain cytokine production and neuroinflammation by endotoxin following ethanol treatment. J. Neuroinflammation 2008, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Lippai, D.; Bala, S.; Csak, T.; Kurt-Jones, E.A.; Szabo, G. Chronic alcohol-induced microRNA-155 contributes to neuroinflammation in a TLR4-dependent manner in mice. PLoS ONE 2013, 8, e70945. [Google Scholar] [CrossRef] [PubMed]
- Kane, C.J.; Phelan, K.D.; Douglas, J.C.; Wagoner, G.; Johnson, J.W.; Xu, J.; Drew, P.D. Effects of ethanol on immune response in the brain: Region-specific changes in aged mice. J. Neuroinflammation 2013, 10, 66. [Google Scholar] [CrossRef]
- Kane, C.J.; Phelan, K.D.; Douglas, J.C.; Wagoner, G.; Johnson, J.W.; Xu, J.; Phelan, P.S.; Drew, P.D. Effects of ethanol on immune response in the brain: Region-specific changes in adolescent versus adult mice. Alcohol. Clin. Exp. Res. 2014, 38, 384–391. [Google Scholar] [CrossRef]
- Pascual, M.; Baliño, P.; Aragón, C.M.; Guerri, C. Cytokines and chemokines as biomarkers of ethanol-induced neuroinflammation and anxiety-related behavior: Role of TLR4 and TLR2. Neuropharmacology 2015, 89, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Niedzwiedz-Massey, V.M.; Douglas, J.C.; Rafferty, T.; Johnson, J.W.; Holloway, K.N.; Berquist, M.D.; Kane, C.J.M.; Drew, P.D. Effects of chronic and binge ethanol administration on mouse cerebellar and hippocampal neuroinflammation. Am. J. Drug Alcohol. Abuse 2023, 49, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Holloway, K.N.; Pinson, M.R.; Douglas, J.C.; Rafferty, T.M.; Kane, C.J.M.; Miranda, R.C.; Drew, P.D. Cerebellar Transcriptomic Analysis in a Chronic plus Binge Mouse Model of Alcohol Use Disorder Demonstrates Ethanol-Induced Neuroinflammation and Altered Glial Gene Expression. Cells 2023, 12, 745. [Google Scholar] [CrossRef]
- Holloway, K.N.; Douglas, J.C.; Rafferty, T.M.; Kane, C.J.M.; Drew, P.D. Ethanol Induces Neuroinflammation in a Chronic Plus Binge Mouse Model of Alcohol Use Disorder via TLR4 and MyD88-Dependent Signaling. Cells 2023, 12, 2109. [Google Scholar] [CrossRef] [PubMed]
- Whitman, B.A.; Knapp, D.J.; Werner, D.F.; Crews, F.T.; Breese, G.R. The cytokine mRNA increase induced by withdrawal from chronic ethanol in the sterile environment of brain is mediated by CRF and HMGB1 release. Alcohol. Clin. Exp. Res. 2013, 37, 2086–2097. [Google Scholar] [CrossRef]
- Ehrlich, D.; Pirchl, M.; Humpel, C. Effects of long-term moderate ethanol and cholesterol on cognition, cholinergic neurons, inflammation, and vascular impairment in rats. Neuroscience 2012, 205, 154–166. [Google Scholar] [CrossRef]
- Freeman, K.; Brureau, A.; Vadigepalli, R.; Staehle, M.M.; Brureau, M.M.; Gonye, G.E.; Hoek, J.B.; Hooper, D.C.; Schwaber, J.S. Temporal changes in innate immune signals in a rat model of alcohol withdrawal in emotional and cardiorespiratory homeostatic nuclei. J. Neuroinflammation 2012, 9, 97. [Google Scholar] [CrossRef]
- Beattie, M.C.; Reguyal, C.S.; Porcu, P.; Daunais, J.B.; Grant, K.A.; Morrow, A.L. Neuroactive Steroid (3α,5α)3-hydroxypregnan-20-one (3α,5α-THP) and Pro-inflammatory Cytokine MCP-1 Levels in Hippocampus CA1 are Correlated with Voluntary Ethanol Consumption in Cynomolgus Monkey. Alcohol. Clin. Exp. Res. 2018, 42, 12–20. [Google Scholar] [CrossRef]
- Cruz, C.; Meireles, M.; Silva, S.M. Chronic ethanol intake induces partial microglial activation that is not reversed by long-term ethanol withdrawal in the rat hippocampal formation. Neurotoxicology 2017, 60, 107–115. [Google Scholar] [CrossRef]
- Alfonso-Loeches, S.; Pascual-Lucas, M.; Blanco, A.M.; Sanchez-Vera, I.; Guerri, C. Pivotal role of TLR4 receptors in alcohol-induced neuroinflammation and brain damage. J. Neurosci. 2010, 30, 8285–8295. [Google Scholar] [CrossRef]
- Pascual, M.; Balino, P.; Alfonso-Loeches, S.; Aragon, C.M.; Guerri, C. Impact of TLR4 on behavioral and cognitive dysfunctions associated with alcohol-induced neuroinflammatory damage. Brain Behav. Immun. 2011, 25 (Suppl. S1), S80–S91. [Google Scholar] [CrossRef] [PubMed]
- Urena-Peralta, J.R.; Alfonso-Loeches, S.; Cuesta-Diaz, C.M.; Garcia-Garcia, F.; Guerri, C. Deep sequencing and miRNA profiles in alcohol-induced neuroinflammation and the TLR4 response in mice cerebral cortex. Sci. Rep. 2018, 8, 15913. [Google Scholar] [CrossRef] [PubMed]
- Morud, J.; Ashouri, A.; Larsson, E.; Ericson, M.; Soderpalm, B. Transcriptional profiling of the rat nucleus accumbens after modest or high alcohol exposure. PLoS ONE 2017, 12, e0181084. [Google Scholar] [CrossRef] [PubMed]
- Wooden, J.I.; Peacoe, L.E.; Anasooya Shaji, C.; Melbourne, J.K.; Chandler, C.M.; Bardo, M.T.; Nixon, K. Adolescent Intermittent Ethanol Drives Modest Neuroinflammation but Does Not Escalate Drinking in Male Rats. Cells 2023, 12, 2572. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, G.M.; Farris, S.P.; Blednov, Y.A.; Harris, R.A.; Mayfield, R.D. Microglial-specific transcriptome changes following chronic alcohol consumption. Neuropharmacology 2018, 128, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Ezquer, F.; Morales, P.; Quintanilla, M.E.; Santapau, D.; Lespay-Rebolledo, C.; Ezquer, M.; Herrera-Marschitz, M.; Israel, Y. Intravenous administration of anti-inflammatory mesenchymal stem cell spheroids reduces chronic alcohol intake and abolishes binge-drinking. Sci. Rep. 2018, 8, 4325. [Google Scholar] [CrossRef] [PubMed]
- Ezquer, F.; Quintanilla, M.E.; Morales, P.; Santapau, D.; Ezquer, M.; Kogan, M.J.; Salas-Huenuleo, E.; Herrera-Marschitz, M.; Israel, Y. Intranasal delivery of mesenchymal stem cell-derived exosomes reduces oxidative stress and markedly inhibits ethanol consumption and post-deprivation relapse drinking. Addict. Biol. 2019, 24, 994–1007. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla, M.E.; Ezquer, F.; Morales, P.; Santapau, D.; Berríos-Cárcamo, P.; Ezquer, M.; Herrera-Marschitz, M.; Israel, Y. Intranasal mesenchymal stem cell secretome administration markedly inhibits alcohol and nicotine self-administration and blocks relapse-intake: Mechanism and translational options. Stem Cell Res. Ther. 2019, 10, 205. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Crews, F.T. Increased MCP-1 and microglia in various regions of the human alcoholic brain. Exp. Neurol. 2008, 210, 349–358. [Google Scholar] [CrossRef]
- Liu, W.; Vetreno, R.P.; Crews, F.T. Hippocampal TNF-death receptors, caspase cell death cascades, and IL-8 in alcohol use disorder. Mol. Psychiatry 2021, 26, 2254–2262. [Google Scholar] [CrossRef]
- Qin, L.; Crews, F.T. NADPH oxidase and reactive oxygen species contribute to alcohol-induced microglial activation and neurodegeneration. J. Neuroinflammation 2012, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Hillmer, A.T.; Sandiego, C.M.; Hannestad, J.; Angarita, G.A.; Kumar, A.; McGovern, E.M.; Huang, Y.; O’Connor, K.C.; Carson, R.E.; O’Malley, S.S.; et al. In vivo imaging of translocator protein, a marker of activated microglia, in alcohol dependence. Mol. Psychiatry 2017, 22, 1759–1766. [Google Scholar] [CrossRef] [PubMed]
- Kalk, N.J.; Guo, Q.; Owen, D.; Cherian, R.; Erritzoe, D.; Gilmour, A.; Ribeiro, A.S.; McGonigle, J.; Waldman, A.; Matthews, P.; et al. Decreased hippocampal translocator protein (18 kDa) expression in alcohol dependence: A [(11)C]PBR28 PET study. Transl. Psychiatry 2017, 7, e996. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Wiers, C.E.; Tyler, R.; Shokri-Kojori, E.; Jang, Y.J.; Zehra, A.; Freeman, C.; Ramirez, V.; Lindgren, E.; Miller, G.; et al. Influence of alcoholism and cholesterol on TSPO binding in brain: PET [(11)C]PBR28 studies in humans and rodents. Neuropsychopharmacology 2018, 43, 1832–1839. [Google Scholar] [CrossRef] [PubMed]
- Tyler, R.E.; Kim, S.W.; Guo, M.; Jang, Y.J.; Damadzic, R.; Stodden, T.; Vendruscolo, L.F.; Koob, G.F.; Wang, G.J.; Wiers, C.E.; et al. Detecting neuroinflammation in the brain following chronic alcohol exposure in rats: A comparison between in vivo and in vitro TSPO radioligand binding. Eur. J. Neurosci. 2019, 50, 1831–1842. [Google Scholar] [CrossRef] [PubMed]
- Luo, J. Effects of Ethanol on the Cerebellum: Advances and Prospects. Cerebellum 2015, 14, 383–385. [Google Scholar] [CrossRef] [PubMed]
- DePaula-Silva, A.B.; Gorbea, C.; Doty, D.J.; Libbey, J.E.; Sanchez, J.M.S.; Hanak, T.J.; Cazalla, D.; Fujinami, R.S. Differential transcriptional profiles identify microglial- and macrophage-specific gene markers expressed during virus-induced neuroinflammation. J. Neuroinflammation 2019, 16, 152. [Google Scholar] [CrossRef] [PubMed]
- Rangaraju, S.; Raza, S.A.; Li, N.X.; Betarbet, R.; Dammer, E.B.; Duong, D.; Lah, J.J.; Seyfried, N.T.; Levey, A.I. Differential Phagocytic Properties of CD45(low) Microglia and CD45(high) Brain Mononuclear Phagocytes-Activation and Age-Related Effects. Front. Immunol. 2018, 9, 405. [Google Scholar] [CrossRef] [PubMed]
- Testa, U.; Pelosi, E.; Castelli, G.; Labbaye, C. miR-146 and miR-155: Two key modulators of immune response and tumor development. Non-Coding RNA 2017, 3, 22. [Google Scholar] [CrossRef]
- O’Connell, R.M.; Taganov, K.D.; Boldin, M.P.; Cheng, G.; Baltimore, D. MicroRNA-155 is induced during the macrophage inflammatory response. Proc. Natl. Acad. Sci. USA 2007, 104, 1604–1609. [Google Scholar] [CrossRef]
- Wang, M.; Mungur, R.; Lan, P.; Wang, P.; Wan, S. MicroRNA-21 and microRNA-146a negatively regulate the secondary inflammatory response of microglia after intracerebral hemorrhage. Int. J. Clin. Exp. Pathol. 2018, 11, 3348–3356. [Google Scholar] [PubMed]
- Sun, L.; Sun, M.; Ma, K.; Liu, J. Let-7d-5p suppresses inflammatory response in neonatal rats with necrotizing enterocolitis via LGALS3-mediated TLR4/NF-kappaB signaling pathway. Am. J. Physiol. Cell Physiol. 2020, 319, C967–C979. [Google Scholar] [CrossRef]
- Melendez, R.I. Intermittent (every-other-day) drinking induces rapid escalation of ethanol intake and preference in adolescent and adult C57BL/6J mice. Alcohol. Clin. Exp. Res. 2011, 35, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Hwa, L.S.; Chu, A.; Levinson, S.A.; Kayyali, T.M.; DeBold, J.F.; Miczek, K.A. Persistent escalation of alcohol drinking in C57BL/6J mice with intermittent access to 20% ethanol. Alcohol. Clin. Exp. Res. 2011, 35, 1938–1947. [Google Scholar] [CrossRef] [PubMed]
- Osterndorff-Kahanek, E.; Ponomarev, I.; Blednov, Y.A.; Harris, R.A. Gene expression in brain and liver produced by three different regimens of alcohol consumption in mice: Comparison with immune activation. PLoS ONE 2013, 8, e59870. [Google Scholar] [CrossRef]
- Erickson, E.K.; Farris, S.P.; Blednov, Y.A.; Mayfield, R.D.; Harris, R.A. Astrocyte-specific transcriptome responses to chronic ethanol consumption. Pharmacogenomics J. 2018, 18, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla, M.E.; Morales, P.; Santapau, D.; Avila, A.; Ponce, C.; Berrios-Carcamo, P.; Olivares, B.; Gallardo, J.; Ezquer, M.; Herrera-Marschitz, M.; et al. Chronic Voluntary Morphine Intake Is Associated with Changes in Brain Structures Involved in Drug Dependence in a Rat Model of Polydrug Use. Int. J. Mol. Sci. 2023, 24, 17081. [Google Scholar] [CrossRef] [PubMed]
- National Institute of Alcohol Abuse and Alcoholism (NIAAA). Understanding Binge Drinking. Available online: https://www.niaaa.nih.gov/publications/brochures-and-fact-sheets/binge-drinking (accessed on 20 February 2024).
- Montesinos, J.; Pascual, M.; Rodriguez-Arias, M.; Minarro, J.; Guerri, C. Involvement of TLR4 in the long-term epigenetic changes, rewarding and anxiety effects induced by intermittent ethanol treatment in adolescence. Brain Behav. Immun. 2016, 53, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Alavez, M.; Nguyen, W.; Mori, S.; Wills, D.N.; Otero, D.; Ehlers, C.L.; Conti, B. Time course of microglia activation and brain and blood cytokine/chemokine levels following chronic ethanol exposure and protracted withdrawal in rats. Alcohol 2019, 76, 37–45. [Google Scholar] [CrossRef]
- Marshall, S.A.; McClain, J.A.; Kelso, M.L.; Hopkins, D.M.; Pauly, J.R.; Nixon, K. Microglial activation is not equivalent to neuroinflammation in alcohol-induced neurodegeneration: The importance of microglia phenotype. Neurobiol. Dis. 2013, 54, 239–251. [Google Scholar] [CrossRef]
- Blednov, Y.A.; Bergeson, S.E.; Walker, D.; Ferreira, V.M.; Kuziel, W.A.; Harris, R.A. Perturbation of chemokine networks by gene deletion alters the reinforcing actions of ethanol. Behav. Brain Res. 2005, 165, 110–125. [Google Scholar] [CrossRef]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.S.; Knapp, D.J.; Crews, F.T. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Nio-Kobayashi, J.; Kudo, M.; Sakuragi, N.; Kimura, S.; Iwanaga, T.; Duncan, W.C. Regulated C-C motif ligand 2 (CCL2) in luteal cells contributes to macrophage infiltration into the human corpus luteum during luteolysis. Mol. Hum. Reprod. 2015, 21, 645–654. [Google Scholar] [CrossRef]
- Dar, M.S. Ethanol-Induced Cerebellar Ataxia: Cellular and Molecular Mechanisms. Cerebellum 2015, 14, 447–465. [Google Scholar] [CrossRef] [PubMed]
- Mitoma, H.; Manto, M.; Shaikh, A.G. Mechanisms of Ethanol-Induced Cerebellar Ataxia: Underpinnings of Neuronal Death in the Cerebellum. Int. J. Environ. Res. Public Health 2021, 18, 8678. [Google Scholar] [CrossRef] [PubMed]
- Haubek, A.; Lee, K. Computed tomography in alcoholic cerebellar atrophy. Neuroradiology 1979, 18, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Koeppen, A.H. The neuropathology of the adult cerebellum. Handb. Clin. Neurol. 2018, 154, 129–149. [Google Scholar] [CrossRef] [PubMed]
- Del Brutto, O.H.; Mera, R.M.; King, N.R.; Zambrano, M.; Sullivan, L.J. Years of Drinking but Not the Amount of Alcohol Intake Contribute to the Association Between Alcoholic Cerebellar Degeneration and Worse Cognitive Performance. A Population-Based Study. Cerebellum 2017, 16, 612–614. [Google Scholar] [CrossRef] [PubMed]
- Del Brutto, O.H.; Mera, R.M.; Sullivan, L.J.; Zambrano, M.; King, N.R. Population-based study of alcoholic cerebellar degeneration: The Atahualpa Project. J. Neurol. Sci. 2016, 367, 356–360. [Google Scholar] [CrossRef]
- Dadsetan, S.; Balzano, T.; Forteza, J.; Agusti, A.; Cabrera-Pastor, A.; Taoro-Gonzalez, L.; Hernandez-Rabaza, V.; Gomez-Gimenez, B.; ElMlili, N.; Llansola, M.; et al. Infliximab reduces peripheral inflammation, neuroinflammation, and extracellular GABA in the cerebellum and improves learning and motor coordination in rats with hepatic encephalopathy. J. Neuroinflammation 2016, 13, 245. [Google Scholar] [CrossRef]
- Agusti, A.; Hernandez-Rabaza, V.; Balzano, T.; Taoro-Gonzalez, L.; Ibanez-Grau, A.; Cabrera-Pastor, A.; Fustero, S.; Llansola, M.; Montoliu, C.; Felipo, V. Sildenafil reduces neuroinflammation in cerebellum, restores GABAergic tone, and improves motor in-coordination in rats with hepatic encephalopathy. CNS Neurosci. Ther. 2017, 23, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Chiva-Blanch, G.; Badimon, L. Benefits and risks of moderate alcohol consumption on cardiovascular disease: Current findings and controversies. Nutrients 2019, 12, 108. [Google Scholar] [CrossRef] [PubMed]
- Middaugh, L.D.; Kelley, B.M.; Bandy, A.-L.E.; McGroarty, K.K. Ethanol consumption by C57BL/6 mice: Influence of gender and procedural variables. Alcohol 1999, 17, 175–183. [Google Scholar] [CrossRef]
- Zhou, Y.; Schwartz, B.I.; Giza, J.; Gross, S.S.; Lee, F.S.; Kreek, M.J. Blockade of alcohol escalation and “relapse” drinking by pharmacological FAAH inhibition in male and female C57BL/6J mice. Psychopharmacology 2017, 234, 2955–2970. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Wan, J. Methodological comparison of FACS and MACS isolation of enriched microglia and astrocytes from mouse brain. J. Immunol. Methods 2020, 486, 112834. [Google Scholar] [CrossRef] [PubMed]
- Benes, V.; Collier, P.; Kordes, C.; Stolte, J.; Rausch, T.; Muckentaler, M.U.; Häussinger, D.; Castoldi, M. Identification of cytokine-induced modulation of microRNA expression and secretion as measured by a novel microRNA specific qPCR assay. Sci. Rep. 2015, 5, 11590. [Google Scholar] [CrossRef]
- Motulsky, H.J.; Brown, R.E. Detecting outliers when fitting data with nonlinear regression—A new method based on robust nonlinear regression and the false discovery rate. BMC Bioinform. 2006, 7, 123. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berríos-Cárcamo, P.; Núñez, S.; Castañeda, J.; Gallardo, J.; Bono, M.R.; Ezquer, F. Two-Month Voluntary Ethanol Consumption Promotes Mild Neuroinflammation in the Cerebellum but Not in the Prefrontal Cortex, Hippocampus, or Striatum of Mice. Int. J. Mol. Sci. 2024, 25, 4173. https://doi.org/10.3390/ijms25084173
Berríos-Cárcamo P, Núñez S, Castañeda J, Gallardo J, Bono MR, Ezquer F. Two-Month Voluntary Ethanol Consumption Promotes Mild Neuroinflammation in the Cerebellum but Not in the Prefrontal Cortex, Hippocampus, or Striatum of Mice. International Journal of Molecular Sciences. 2024; 25(8):4173. https://doi.org/10.3390/ijms25084173
Chicago/Turabian StyleBerríos-Cárcamo, Pablo, Sarah Núñez, Justine Castañeda, Javiera Gallardo, María Rosa Bono, and Fernando Ezquer. 2024. "Two-Month Voluntary Ethanol Consumption Promotes Mild Neuroinflammation in the Cerebellum but Not in the Prefrontal Cortex, Hippocampus, or Striatum of Mice" International Journal of Molecular Sciences 25, no. 8: 4173. https://doi.org/10.3390/ijms25084173
APA StyleBerríos-Cárcamo, P., Núñez, S., Castañeda, J., Gallardo, J., Bono, M. R., & Ezquer, F. (2024). Two-Month Voluntary Ethanol Consumption Promotes Mild Neuroinflammation in the Cerebellum but Not in the Prefrontal Cortex, Hippocampus, or Striatum of Mice. International Journal of Molecular Sciences, 25(8), 4173. https://doi.org/10.3390/ijms25084173