
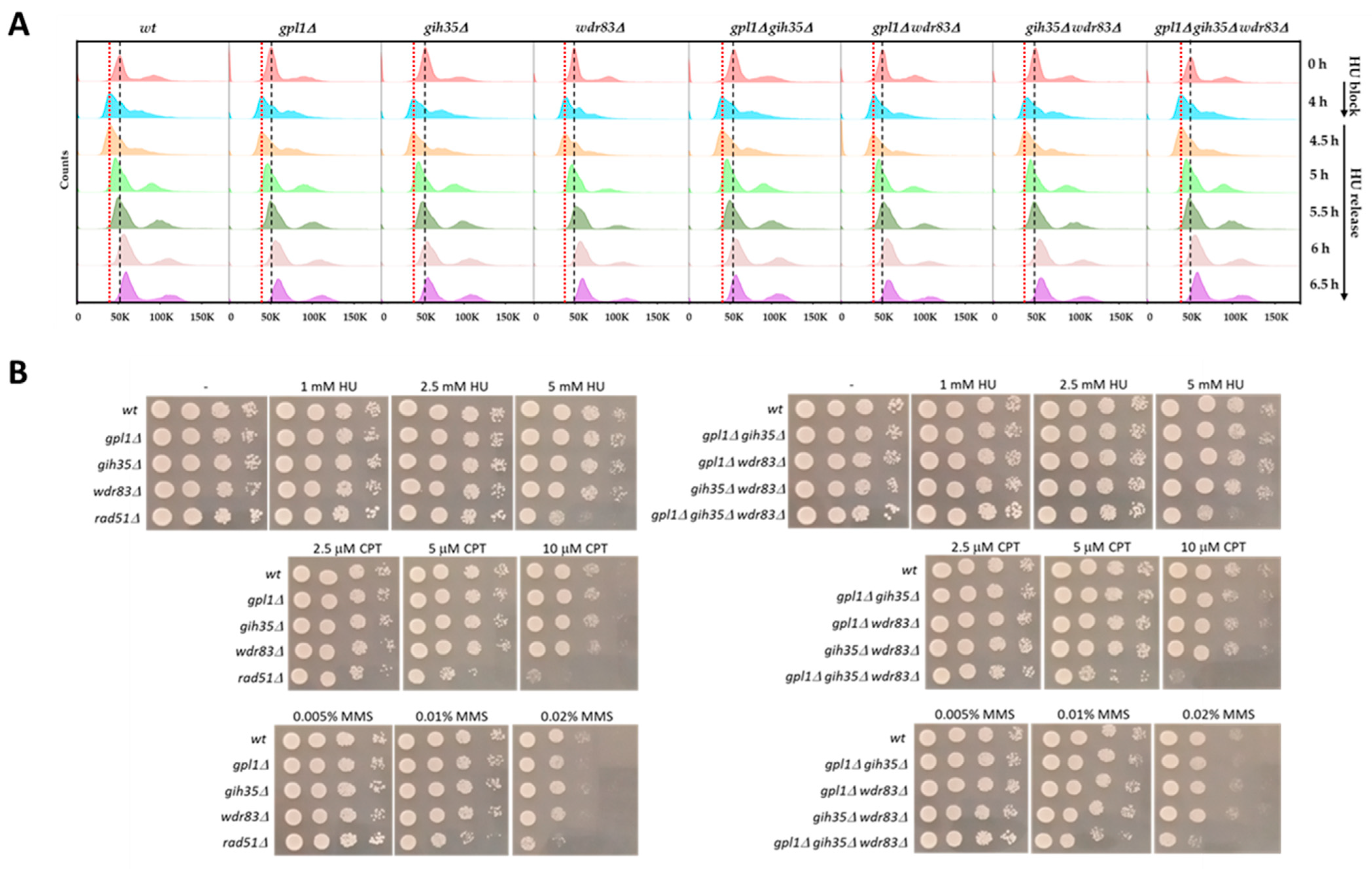
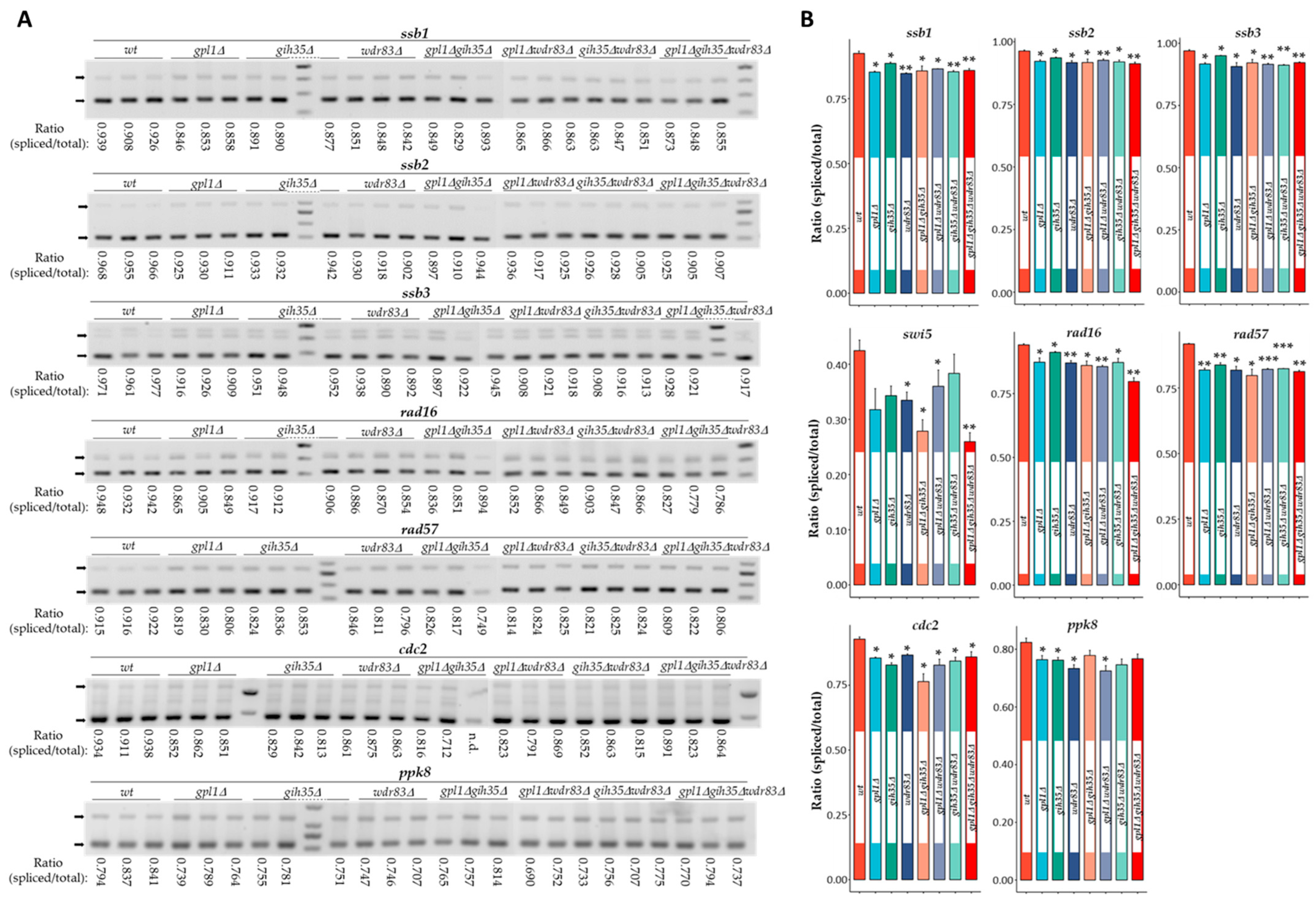
Dysfunction of Gpl1–Gih35–Wdr83 Complex in S. pombe Affects the Splicing of DNA Damage Repair Factors Resulting in Increased Sensitivity to DNA Damage

 ,
, {kind=link}
{kind=link}
Abstract
Share and Cite
Cipakova, I.; Jurcik, M.; Selicky, T.; Lalakova, L.O.; Jakubikova, J.; Cipak, L. Dysfunction of Gpl1–Gih35–Wdr83 Complex in S. pombe Affects the Splicing of DNA Damage Repair Factors Resulting in Increased Sensitivity to DNA Damage. Int. J. Mol. Sci. 2024, 25, 4192. https://doi.org/10.3390/ijms25084192
Cipakova I, Jurcik M, Selicky T, Lalakova LO, Jakubikova J, Cipak L. Dysfunction of Gpl1–Gih35–Wdr83 Complex in S. pombe Affects the Splicing of DNA Damage Repair Factors Resulting in Increased Sensitivity to DNA Damage. International Journal of Molecular Sciences. 2024; 25(8):4192. https://doi.org/10.3390/ijms25084192
Chicago/Turabian StyleCipakova, Ingrid, Matus Jurcik, Tomas Selicky, Laura Olivia Lalakova, Jana Jakubikova, and Lubos Cipak. 2024. "Dysfunction of Gpl1–Gih35–Wdr83 Complex in S. pombe Affects the Splicing of DNA Damage Repair Factors Resulting in Increased Sensitivity to DNA Damage" International Journal of Molecular Sciences 25, no. 8: 4192. https://doi.org/10.3390/ijms25084192
APA StyleCipakova, I., Jurcik, M., Selicky, T., Lalakova, L. O., Jakubikova, J., & Cipak, L. (2024). Dysfunction of Gpl1–Gih35–Wdr83 Complex in S. pombe Affects the Splicing of DNA Damage Repair Factors Resulting in Increased Sensitivity to DNA Damage. International Journal of Molecular Sciences, 25(8), 4192. https://doi.org/10.3390/ijms25084192