
Identification of Plasma Lipid Alterations Associated with Melanoma Metastasis


 , ,
, , 
Abstract
1. Introduction
2. Results
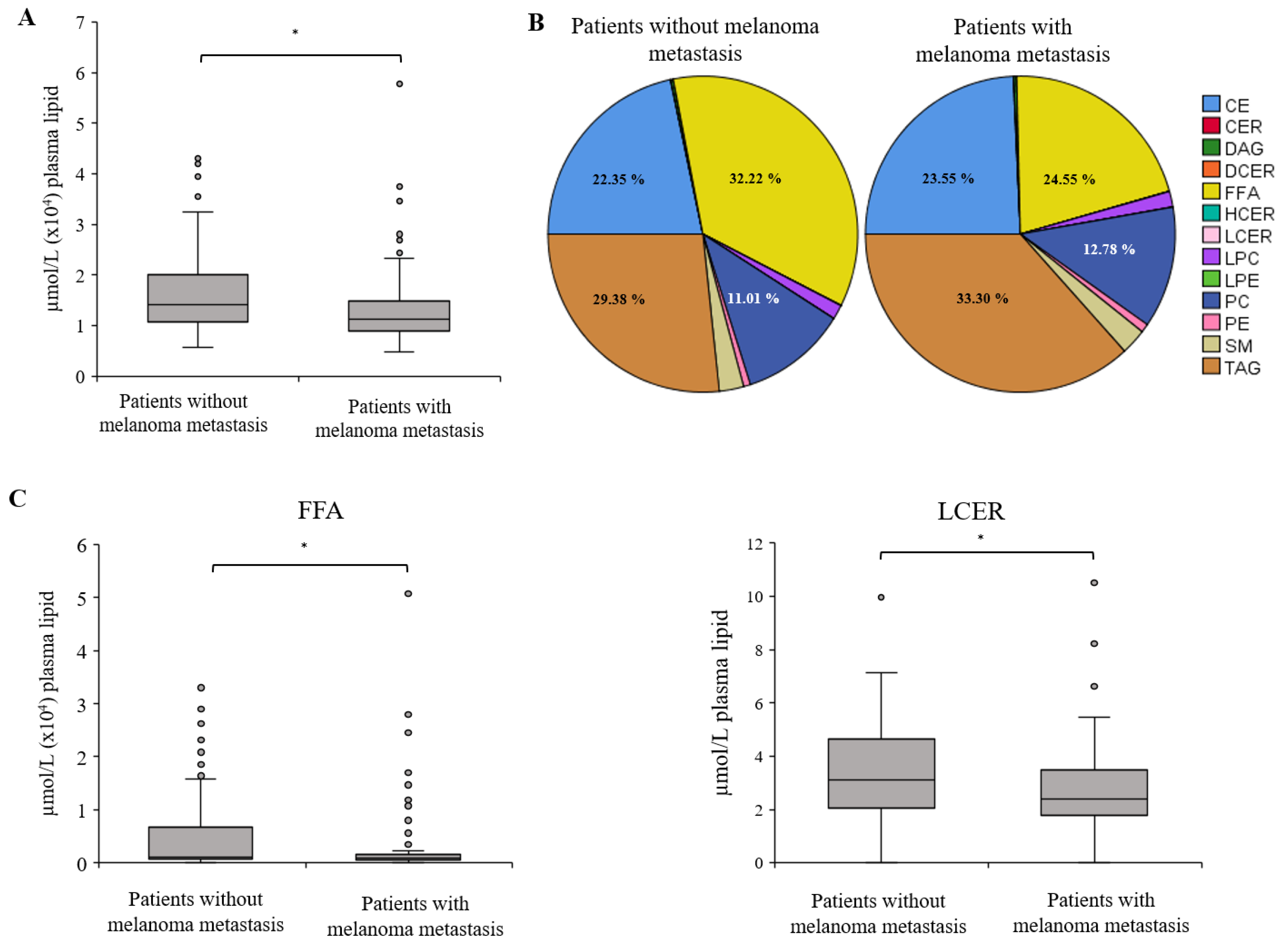
2.1. Distribution of Lipid Classes between Melanoma Patients without and with Metastasis
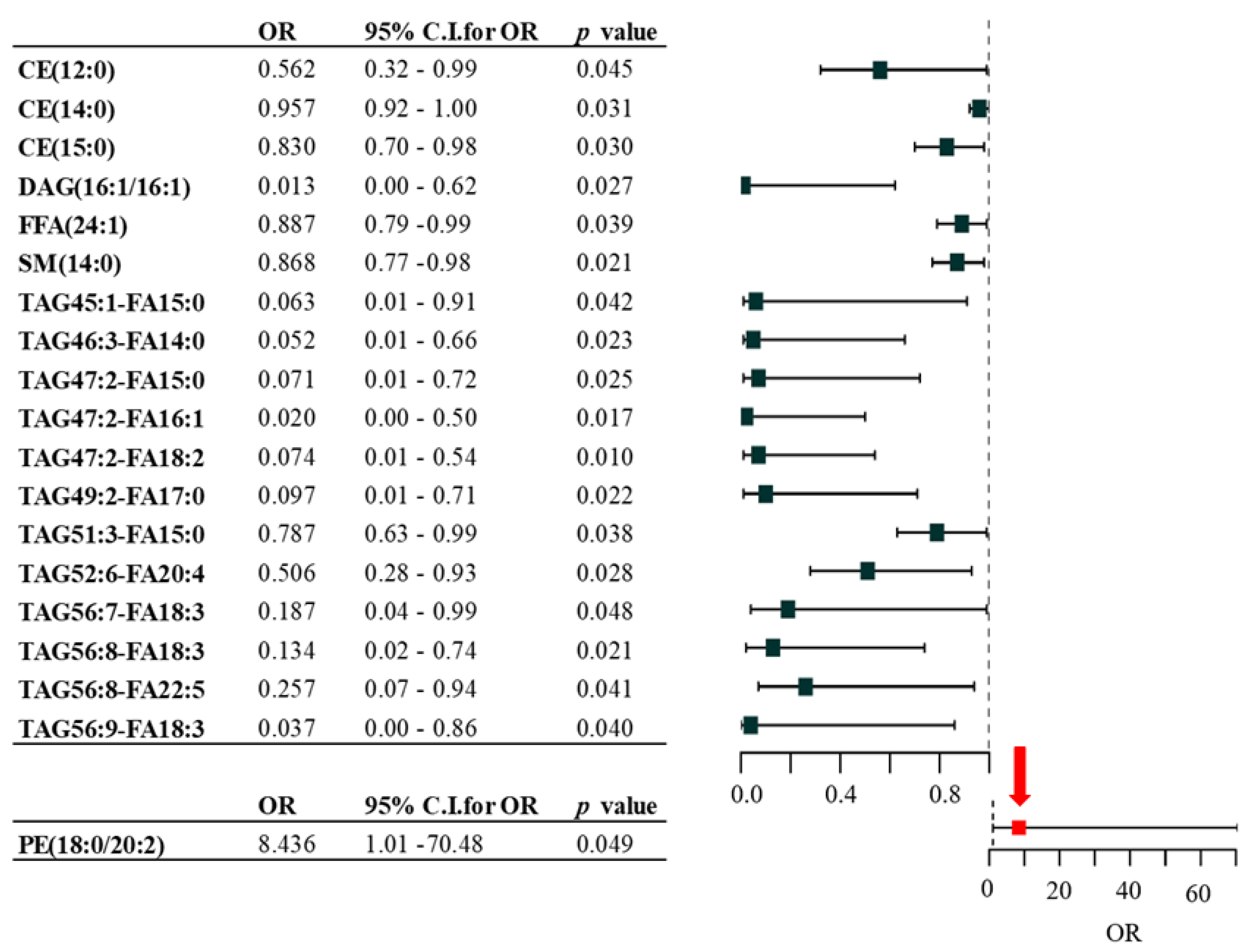
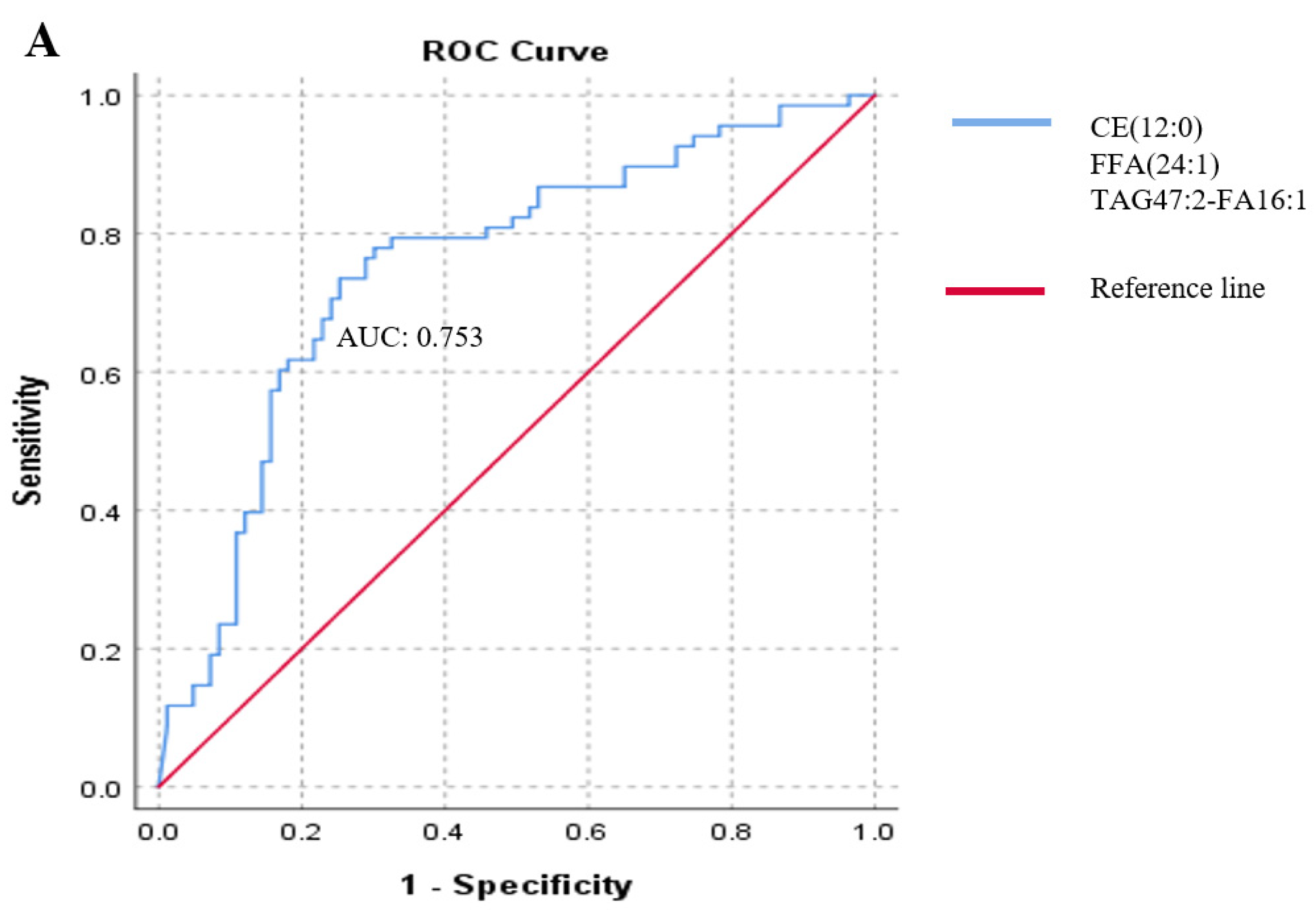
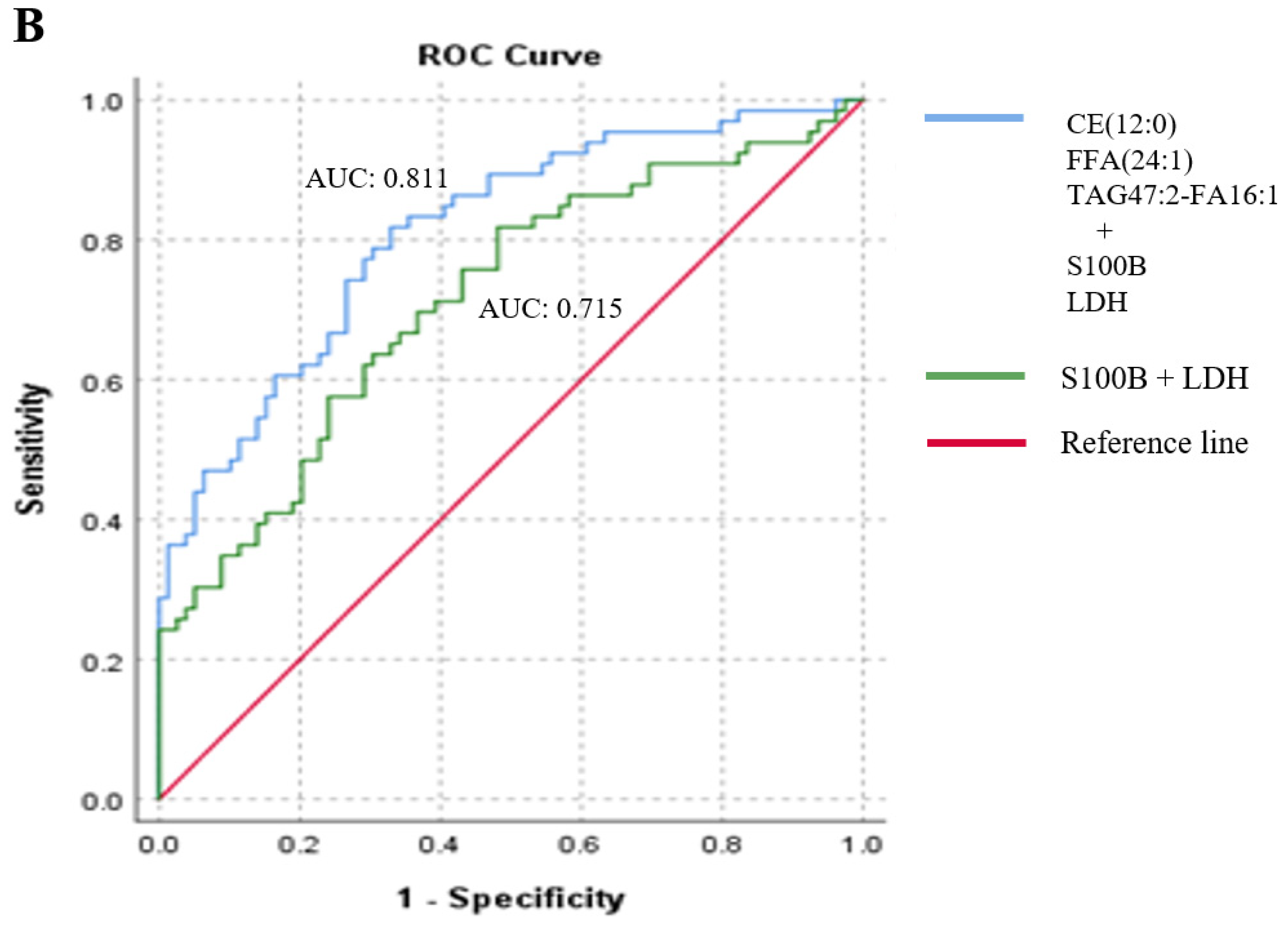
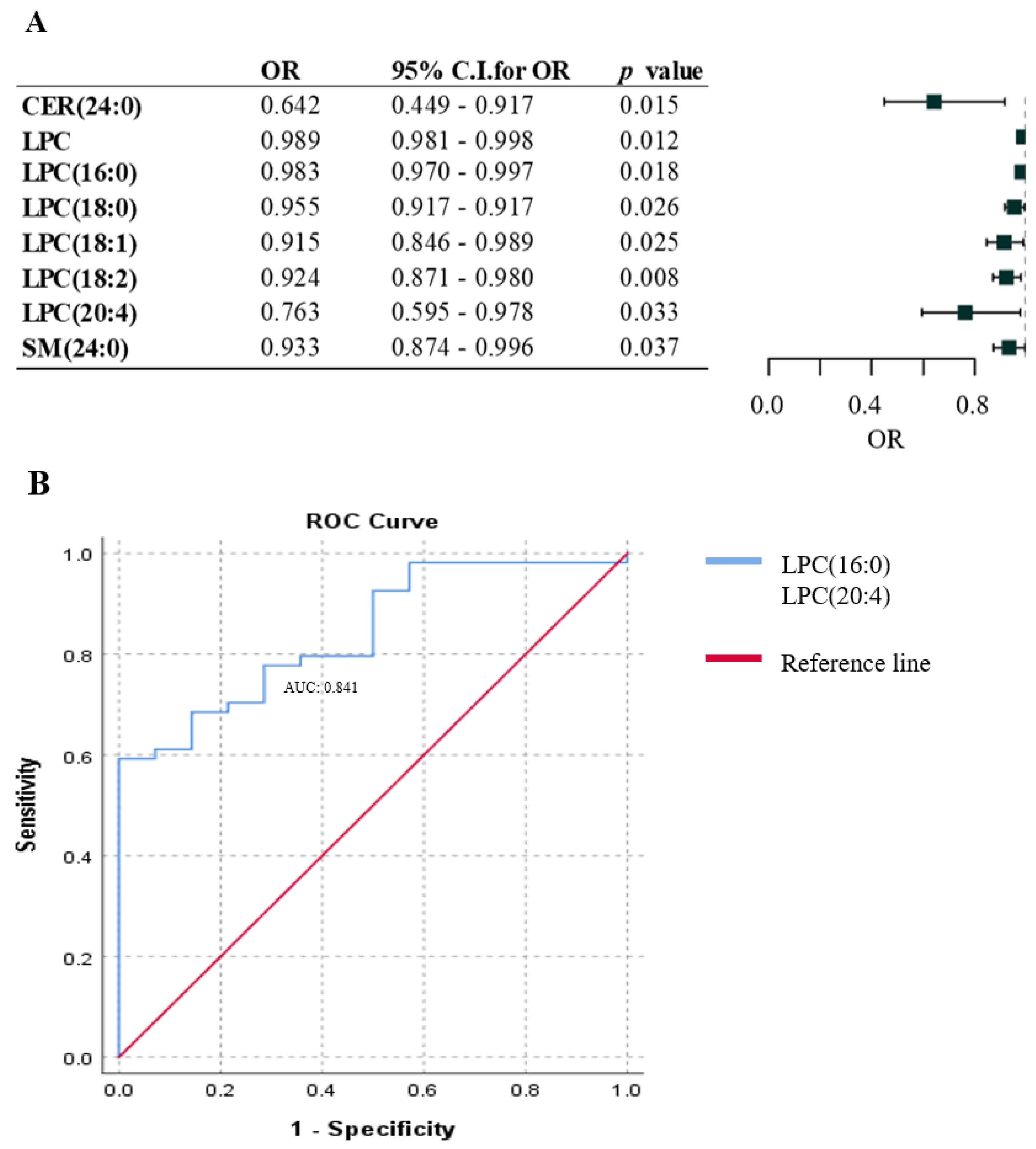
2.2. Association between Lipid Species and the Presence of Metastasis in Melanoma Patients
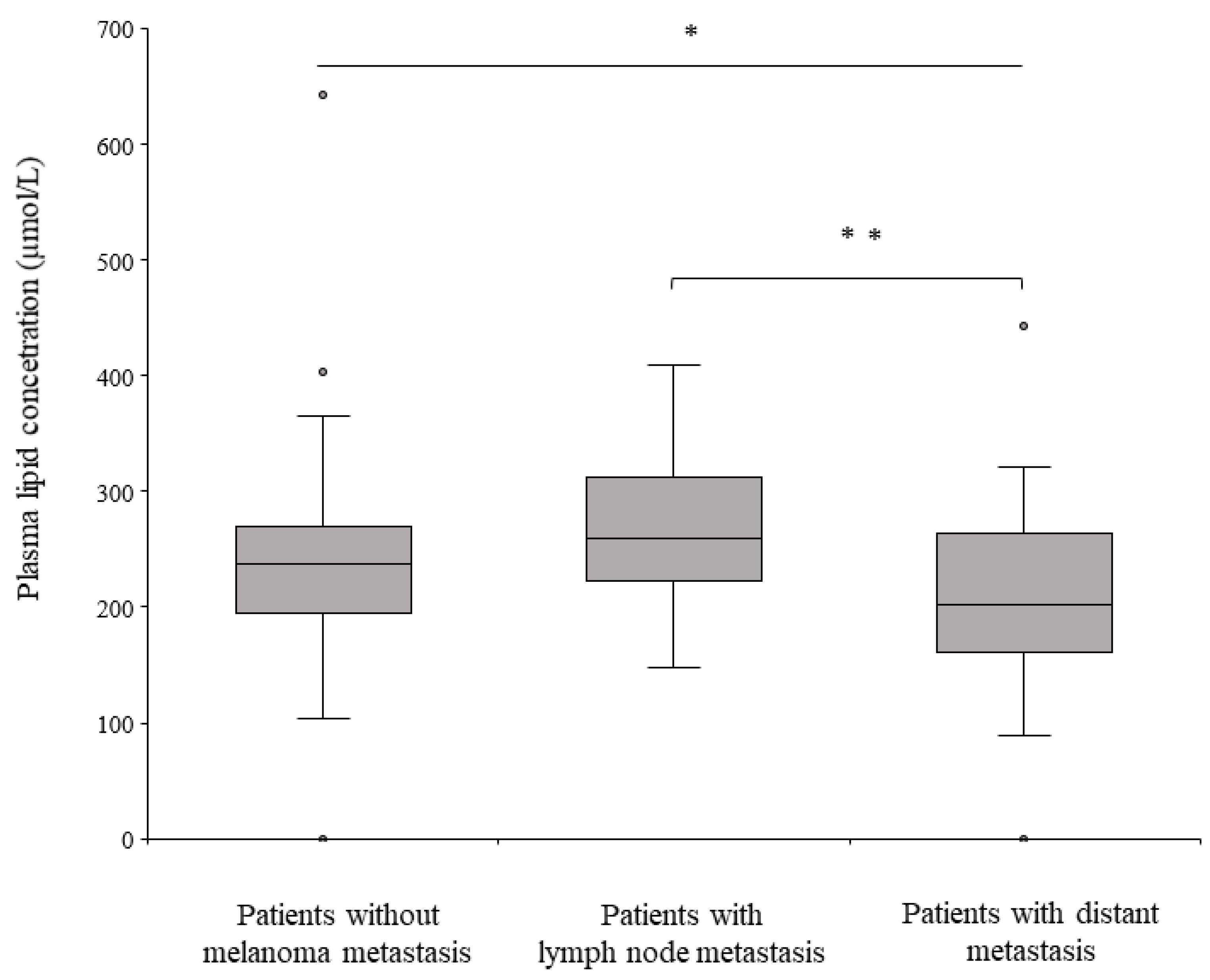
2.3. Lipid Species Associated with the Lymphatic and Hematogenous Pathways of Melanoma Metastasis
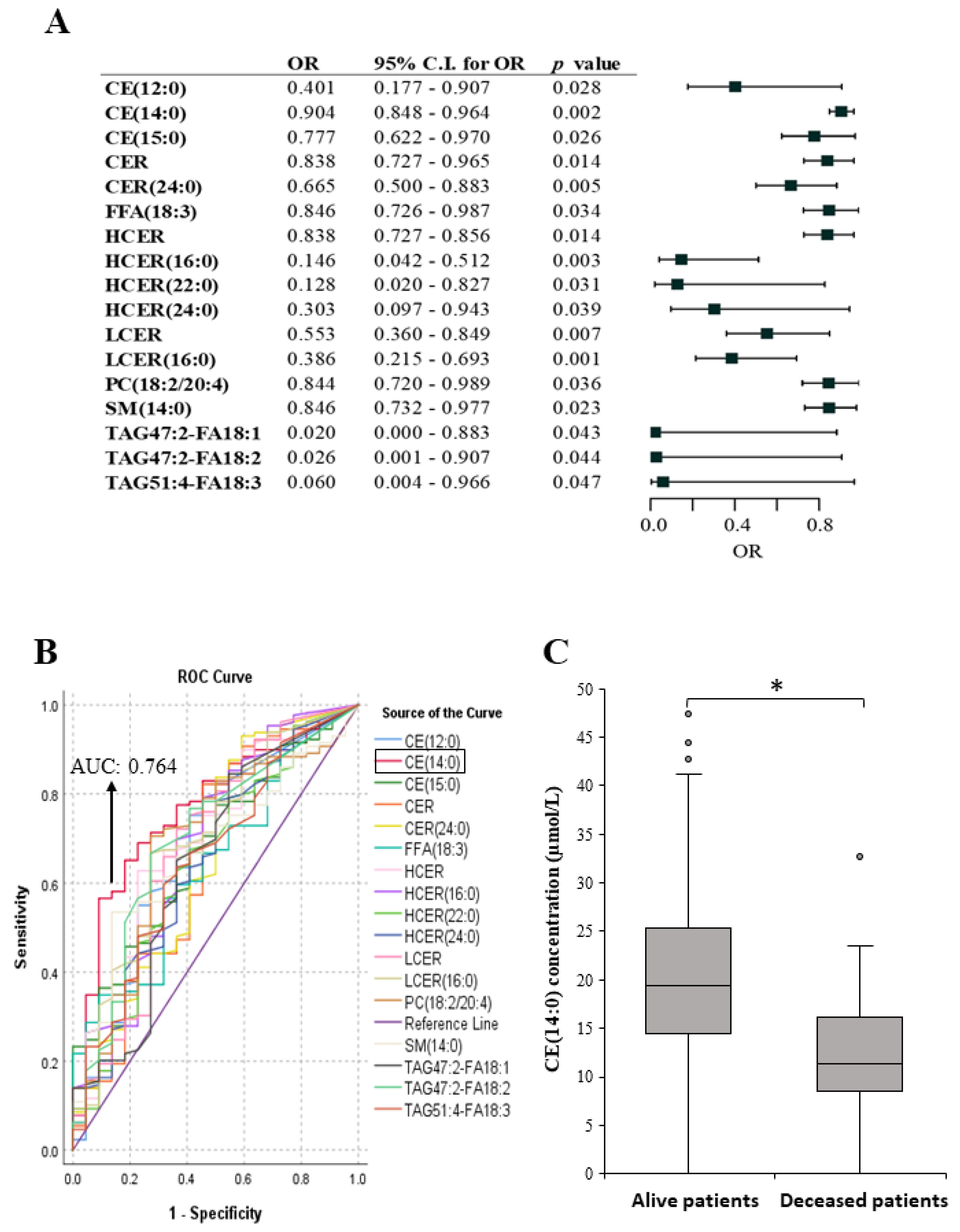
2.4. Lipid Species Associated with Melanoma Patient’s Survival
3. Discussion
4. Materials and Methods
4.1. Characteristics of Patients and Melanoma Samples Included in the Study

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Therapy at the Time of Blood Sampling | |||||
|---|---|---|---|---|---|
| Number of Patients (%) | Metastases Peresent (%) | None (%) | Immuno-Therapy a (%) | Targeted Therapy b (%) | |
| All patients | 151 (100) | 68 (45.0) | 64 (43.4) | 64 (43.4) | 23 (15.2) |
| Gender | |||||
| Female | 67 (44.4) | 23 (34.3) | 31 (46.2) | 27 (40.3) | 9 (13.4) |
| Male | 84 (55.6) | 45 (53.6) | 33 (33.3) | 37 (44.0) | 14 (16.7) |
| Age (years) (average age: 61.91) | |||||
| 20–50 | 28 (18.5) | 8 (28.6) | 14 (50.0) | 9 (32.1) | 5 (17.9) |
| ≥50 | 123 (81.5) | 60 (48.8) | 50 (40.7) | 55(44.7) | 18 (14.6) |
| Metastasis | |||||
| Absent | 83 (55.0) | 83 (55.0) | 53 (63.9) | 22 (26.5) | 8 (9.6) |
| Present | 68 (45.0) | 68 (45.0) | 11 (16.2) | 42 (61.8) | 15 (22.1) |
| Pathway of metastasis | |||||
| Lymphatic (only lymph node) | 19 (12.6) | ||||
| Hematogenous (distant and/or cutan) | 49 (32.5) | ||||
| Patient’s survival (with a 3-year follow-up period) | |||||
| Alive | 129 (85.4) | ||||
| Deceased | 22 (14.6) | ||||
4.2. Blood Samples
4.3. Standards and Extraction of Lipids
4.4. Lipidomic Analysis and Data Processing
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ning, B.; Liu, Y.; Wang, M.; Li, Y.; Xu, T.; We, Y. The Predictive Value of Tumor Mutation Burden on Clinical Efficacy of Immune Checkpoint Inhibitors in Melanoma: A Systematic Review and Meta-Analysis. Front. Pharmacol. 2022, 13, 748674. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O.; Wind, F.; Negelein, E. The Metabolism of Tumors in the Body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Swinnen, J.V.; Brusselmans, K.; Verhoeven, G. Increased lipogenesis in cancer cells: New players, novel targets. Curr. Opin. Clin. Nutr. Metab. Care 2006, 9, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Spector, A.A.; Steinberg, D. Relationship between fatty acid and glucose utilization in Ehrlich ascites tumor cells. J. Lipid Res. 1966, 7, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Tator, C.H.; Evans, J.R.; Olszewski, J. Tracers for the detection of brain tumors. Evaluation of radioiodinated human serum albumin and radioiodinated fatty acid. Neurology 1966, 16, 650–661. [Google Scholar] [CrossRef] [PubMed]
- Jensen, V.; Ladekarl, M.; Holm-Nielsen, P.; Melsen, F.; Soerensen, F.B. The prognostic value of oncogenic antigen 519 (OA-519) expression and proliferative activity detected by antibody MIB-1 in node-negative breast cancer. J. Pathol. 1995, 176, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Butler, L.M.; Perone, Y.; Dehairs, J.; Lupien, L.E.; de Laat, V.; Talebi, A.; Loda, M.; Kinlaw, W.B.; Swinnen, J.V. Lipids and cancer: Emerging roles in pathogenesis, diagnosis and therapeutic intervention. Adv. Drug Deliv. Rev. 2020, 159, 245–293. [Google Scholar] [CrossRef]
- Hao, Y.; Li, D.; Xu, Y.; Ouyang, J.; Wang, Y.; Zhang, Y.; Li, B.; Xie, L.; Qin, G. Investigation of lipid metabolism dysregulation and the effects on immune microenvironments in pan-cancer using multiple omics data. BMC Bioinform. 2019, 20, 195. [Google Scholar] [CrossRef] [PubMed]
- Peck, B.; Schulze, A. Lipid Metabolism at the Nexus of Diet and Tumor Microenvironment. Trends Cancer 2019, 5, 693–703. [Google Scholar] [CrossRef]
- Nagarajan, S.R.; Butler, L.M.; Hoy, A.J. The diversity and breadth of cancer cell fatty acid metabolism. Cancer Metab. 2021, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Riscal, R.; Skuli, N.; Simon, M.C. Even Cancer Cells Watch Their Cholesterol! Mol. Cell 2019, 76, 220–231. [Google Scholar] [CrossRef]
- Mollinedo, F.; Gajate, C. Lipid rafts as signaling hubs in cancer cell survival/death and invasion: Implications in tumor progression and therapy: Thematic Review Series: Biology of Lipid Rafts. J. Lipid Res. 2020, 61, 611–635. [Google Scholar] [CrossRef]
- Kim, H.Y.; Lee, H.; Kim, S.H.; Jin, H.; Bae, J.; Choi, H.K. Discovery of potential biomarkers in human melanoma cells with different metastatic potential by metabolic and lipidomic profiling. Sci. Rep. 2017, 7, 8864. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, X.; Qiu, L.; Qin, X.; Liu, H.; Wang, Y.; Li, F.; Wang, X.; Chen, G.; Song, G.; et al. Probing gender-specific lipid metabolites and diagnostic biomarkers for lung cancer using Fourier transform ion cyclotron resonance mass spectrometry. Clin. Chim. Acta 2012, 414, 135–141. [Google Scholar] [CrossRef]
- Patel, N.; Vogel, R.; Chandra-Kuntal, K.; Glasgow, W.; Kelavkar, U. A novel three serum phospholipid panel differentiates normal individuals from those with prostate cancer. PLoS ONE 2014, 9, e88841. [Google Scholar] [CrossRef] [PubMed]
- Cas, M.D.; Ciniselli, C.M.; Vergani, E.; Ciusani, E.; Aloisi, M.; Duroni, V.; Verderio, P.; Ghidoni, R.; Paroni, R.; Perego, P.; et al. Alterations in Plasma Lipid Profiles Associated with Melanoma and Therapy Resistance. Int. J. Mol. Sci. 2024, 25, 1558. [Google Scholar] [CrossRef]
- Fu, Y.; Zou, T.; Shen, X.; Nelson, P.J.; Li, J.; Wu, C.; Yang, J.; Zheng, Y.; Bruns, C.; Zhao, Y.; et al. Lipid metabolism in cancer progression and therapeutic strategies. MedComm 2021, 2, 27–59. [Google Scholar] [CrossRef]
- Aloia, A.; Müllhaupt, D.; Chabbert, C.D.; Eberhart, T.; Flückiger-Mangual, S.; Vukolic, A.; Eichhoff, O.; Irmisch, A.; Alexander, L.T.; Scibona, E.; et al. A Fatty Acid Oxidation-dependent Metabolic Shift Regulates the Adaptation of BRAF-mutated Melanoma to MAPK Inhibitors. Clin. Cancer Res. 2019, 25, 6852–6867. [Google Scholar] [CrossRef]
- Rysman, E.; Brusselmans, K.; Scheys, K.; Timmermans, L.; Derua, R.; Munck, S.; Van Veldhoven, P.P.; Waltregny, D.; Daniëls, V.W.; Machiels, J.; et al. De novo lipogenesis protects cancer cells from free radicals and chemotherapeutics by promoting membrane lipid saturation. Cancer Res. 2010, 70, 8117–8126. [Google Scholar] [CrossRef]
- Wang, H.; Franco, F.; Tsui, Y.C.; Xie, X.; Trefny, M.P.; Zappasodi, R.; Mohmood, S.R.; Fernández-García, J.; Tsai, C.H.; Schulze, I.; et al. CD36-mediated metabolic adaptation supports regulatory T cell survival and function in tumors. Nat. Immunol. 2020, 21, 298–308. [Google Scholar] [CrossRef]
- Stephenson, D.J.; Hoeferlin, L.A.; Chalfant, C.E. Lipidomics in translational research and the clinical significance of lipid-based biomarkers. Transl. Res. 2017, 189, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Perrotti, F.; Rosa, C.; Cicalini, I.; Sacchetta, P.; Del Boccio, P.; Genovesi, D.; Pieragostino, D. Advances in Lipidomics for Cancer Biomarkers Discovery. Int. J. Mol. Sci. 2016, 17, 1992. [Google Scholar] [CrossRef]
- Cvetkovic, Z.; Cvetkovic, B.; Petrovic, M.; Ranic, M.; Debeljak-Martarcic, J.; Vucic, V.; Glibetic, M. Lipid profile as a prognostic factor in cancer patients. J BUON 2009, 14, 501–506. [Google Scholar]
- Muntoni, S.; Atzori, L.; Mereu, R.; Satta, G.; Macis, M.D.; Congia, M.; Tedde, A.; Desogus, A.; Muntoni, S. Serum lipoproteins and cancer. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Ghahremanfard, F.; Mirmohammadkhani, M.; Shahnazari, B.; Gholami, G.; Mehdizadeh, J. The Valuable Role of Measuring Serum Lipid Profile in Cancer Progression. Oman Med. J. 2015, 30, 353–357. [Google Scholar] [CrossRef]
- Li, R.; Liu, B.; Liu, Y.; Liu, Y.; He, Y.; Wang, D.; Sun, Y.; Xu, Y.; Yu, Q. Elevated serum lipid level can serve as early signal for metastasis for Non-Small Cell Lung Cancer patients: A retrospective nested case-control study. J. Cancer 2020, 11, 7023–7031. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Han, L.; He, J.; Lv, J.; Pan, R.; Lv, T. A high serum-free fatty acid level is associated with cancer. J. Cancer Res. Clin. Oncol. 2020, 146, 705–710. [Google Scholar] [CrossRef]
- Fan, L.; Lin, Q.; Huang, X.; Fu, D.; Huang, H. Prognostic significance of pretreatment serum free fatty acid in patients with diffuse large B-cell lymphoma in the rituximab era: A retrospective analysis. BMC Cancer 2021, 21, 1255. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, L.; Liu, N.; He, C.; Li, Z. Decreased serum levels of free fatty acids are associated with breast cancer. Clin. Chim. Acta 2014, 437, 31–37. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef]
- Faedo, R.R.; da Silva, G.; da Silva, R.M.; Ushida, T.R.; da Silva, R.R.; Lacchini, R.; Matos, L.L.; Kowalski, L.P.; Lopes, N.P.; Leopoldino, A.M. Sphingolipids signature in plasma and tissue as diagnostic and prognostic tools in oral squamous cell carcinoma. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2022, 1867, 159057. [Google Scholar] [CrossRef] [PubMed]
- Nath, A.; Li, I.; Roberts, L.R.; Chan, C. Elevated free fatty acid uptake via CD36 promotes epithelial-mesenchymal transition in hepatocellular carcinoma. Sci. Rep. 2015, 5, 14752. [Google Scholar] [CrossRef] [PubMed]
- Nath, A.; Chan, C. Genetic alterations in fatty acid transport and metabolism genes are associated with metastatic progression and poor prognosis of human cancers. Sci. Rep. 2016, 6, 18669. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V.; Vijayakumar, M.V.; Ajay, A.K.; Malvi, P.; Bhat, M.K. Diet-induced obesity increases melanoma progression: Involvement of Cav-1 and FASN. Int. J. Cancer 2012, 30, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Ueno, G.; Iwagami, Y.; Kobayashi, S.; Mitsufuji, S.; Yamada, D.; Tomimaru, Y.; Akita, H.; Asaoka, T.; Noda, T.; Gotoh, K.; et al. CAT-1-Regulated Cholesteryl Ester Accumulation Modulates Gemcitabine Resistance in Biliary Tract Cancer. Ann. Surg. Oncol. 2022, 29, 2899–2909. [Google Scholar] [CrossRef]
- Chang, T.Y.; Chang, C.C.; Cheng, D. Acyl-coenzyme A:cholesterol acyltransferase. Annu. Rev. Biochem. 1997, 66, 613–638. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.C. Molecular mechanisms of cellular cholesterol efflux. J. Biol. Chem. 2014, 289, 24020–24029. [Google Scholar] [CrossRef]
- Hartmann, P.; Trufa, D.I.; Hohenberger, K.; Tausche, P.; Trump, S.; Mittler, S.; Geppert, C.I.; Rieker, R.J.; Schieweck, O.; Sirbu, H.; et al. Contribution of serum lipids and cholesterol cellular metabolism in lung cancer development and progression. Sci. Rep. 2023, 13, 5662. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Nilsson-Ehle, P.; Xu, N. Influence of liver cancer on lipid and lipoprotein metabolism. Lipids Health Dis. 2006, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Motta, M.; Giugno, I.; Ruello, P.; Pistone, G.; Di Fazio, I.; Malaguarnera, M. Lipoprotein (a) behaviour in patients with hepatocellular carcinoma. Minerva Med. 2001, 92, 301–305. [Google Scholar] [PubMed]
- Argiles, J.M.; Lopez-Soriano, F.J.; Evans, R.D.; Williamson, D.H. Interleukin-1 and lipid metabolism in the rat. Biochem. J. 1989, 259, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Kwong, L.K.; Ridinger, D.N.; Bandhauer, M.; Ward, J.H.; Samlowski, W.E.; Iverius, P.H.; Pritchard, H.; Wilson, D.E. Acute dyslipoproteinemia induced by interleukin-2: Lecithin: Cholesteryl acyltransferase, lipoprotein lipase, and hepatic lipase deficiencies. J. Clin. Endocrinol. Metab. 1997, 82, 1572–1581. [Google Scholar] [CrossRef][Green Version]
- Zimmerman, W.F.; Keys, S. Lysophospholipase and the metabolism of lysophosphatidylcholine in isolated bovine rod outer segments. Exp. Eye Res. 1989, 48, 69–76. [Google Scholar] [CrossRef]
- Liu, P.; Zhu, W.; Chen, C.; Yan, B.; Zhu, L.; Chen, X.; Peng, C. The mechanisms of lysophosphatidylcholine in the development of diseases. Life Sci. 2020, 247, 117443. [Google Scholar] [CrossRef]
- Kuliszkiewicz-Janus, M.; Tuz, M.A.; Baczynski, S. Application of 31P MRS to the analysis of phospholipid changes in plasma of patients with acute leukemia. Biochim. Biophys. Acta 2005, 1737, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.A.; Wilke, M.S.; Perrine, M.; Pawlowicz, M.; Mourtzakis, M.; Lieffers, J.R.; Maneshgar, M.; Bruera, E.; Clandinin, M.T.; Baracos, V.E.; et al. Loss of adipose tissue and plasma phospholipids: Relationship to survival in advanced cancer patients. Clin. Nutr. 2010, 29, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Süllentrop, F.; Moka, D.; Neubauer, S.; Haupt, G.; Engelmann, U.; Hahn, J.; Schicha, H. 31P NMR spectroscopy of blood plasma: Determination and quantification of phospholipid classes in patients with renal cell carcinoma. NMR Biomed. 2002, 15, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.A.; Bureyko, T.F.; Mourtzakis, M.; Chu, Q.S.; Clandinin, M.T.; Reiman, T.; Mazurak, V.C. Aberrations in plasma phospholipid fatty acids in lung cancer patients. Lipids 2012, 47, 363–369. [Google Scholar] [CrossRef]
- Zhao, Z.; Xiao, Y.; Elson, P.; Tan, H.; Plummer, S.J.; Berk, M.; Aung, P.P.; Lavery, I.C.; Achkar, J.P.; Li, L.; et al. Plasma lysophosphatidylcholine levels: Potential biomarkers for colorectal cancer. J. Clin. Oncol. 2007, 25, 2696–2701. [Google Scholar] [CrossRef] [PubMed]
- Ojala, P.J.; Hirvonen, T.E.; Hermansson, M.; Somerharju, P.; Parkkinen, J. Acyl chain-dependent effect of lysophosphatidyl-choline on human neutrophils. J. Leukoc. Biol. 2007, 82, 1501–1509. [Google Scholar] [CrossRef] [PubMed]
- Brkic, L.; Riederer, M.; Graier, W.F.; Malli, R.; Frank, S. Acyl chain-dependent effect of lysophosphatidylcholine on cyclooxygenase (COX)-2 expression in endothelial cells. Atherosclerosis 2012, 224, 348–354. [Google Scholar] [CrossRef]
- Kang, S.; Kim, M.H.; Park, I.A.; Kim, J.W.; Park, N.H.; Kang, D.; Yoo, K.Y.; Kang, S.B.; Lee, H.P.; Song, Y.S. Elevation of cyclooxygenase-2 is related to lymph node metastasis in adenocarcinoma of uterine cervix. Cancer Lett. 2006, 237, 305–311. [Google Scholar] [CrossRef]
- Kyzas, P.A.; Stefanou, D.; Agnantis, N.J. COX-2 expression correlates with VEGF-C and lymph node metastases in patients with head and neck squamous cell carcinoma. Mod. Pathol. 2005, 18, 153–160. [Google Scholar] [CrossRef]
- Crofford, L.J. COX-1 and COX-2 tissue expression: Implications and predictions. J. Rheumatology. Suppl. 1997, 49, 15–19. [Google Scholar]
- Denkert, C.; Köbel, M.; Berger, S.; Siegert, A.; Leclere, A.; Trefzer, U.; Hauptmann, S. Expression of cyclooxygenase 2 in human malignant melanoma. Cancer Res. 2001, 61, 303–308. [Google Scholar]
- Zhou, P.; Qin, J.; Li, Y.; Li, G.; Wang, Y.; Zhang, N.; Chen, P.; Li, C. Combination therapy of PKCzeta and COX-2 inhibitors synergistically suppress melanoma metastasis. J. Exp. Clin. Cancer Res. 2017, 36, 115. [Google Scholar] [CrossRef] [PubMed]
- Tudor, D.V.; Bâldea, I.; Lupu, M.; Kacso, T.; Kutasi, E.; Hopârtean, A.; Stretea, R.; Gabriela Filip, A. COX-2 as a potential biomarker and therapeutic target in melanoma. Cancer Biol. Med. 2020, 17, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Gerl, M.J.; Vaz, W.L.C.; Domingues, N.; Klose, C.; Surma, M.A.; Sampaio, J.L.; Almeida, M.S.; Rodrigues, G.; Araújo-Gonçalves, P.; Ferreira, J.; et al. Cholesterol is Inefficiently Converted to Cholesteryl Esters in the Blood of Cardiovascular Disease Patients. Sci. Rep. 2018, 8, 14764. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gomez, C.R.; Espinoza, I.; Le, T.P.T.; Shenoy, V.; Zhou, X. Correlation of cholesteryl ester metabolism to pathogenesis, progression and disparities in colorectal Cancer. Lipids Health Dis. 2022, 21, 22. [Google Scholar] [CrossRef]
- Tisdale, M.J. Cachexia in cancer patients. Nat. Rev. Cancer 2002, 2, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Zwickl, H.; Hackner, K.; Köfeler, H.; Krzizek, E.C.; Muqaku, B.; Pils, D.; Scharnagl, H.; Solheim, T.S.; Zwickl-Traxler, E.; Pecherstorfer, M. Reduced LDL-Cholesterol and Reduced Total Cholesterol as Potential Indicators of Early Cancer in Male Treatment-Naive Cancer Patients With Pre-cachexia and Cachexia. Front. Oncol. 2020, 10, 1262. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, X.W.; Liu, D.B.; Han, C.Z.; Du, L.L.; Jing, J.X.; Wang, Y. Lipid levels in serum and cancerous tissues of colorectal cancer patients. World J. Gastroenterol. 2014, 20, 8646–8652. [Google Scholar] [CrossRef] [PubMed]
- Pikó, P.; Pál, L.; Szűcs, S.; Kósa, Z.; Sándor, J.; Ádány, R. Obesity-Related Changes in Human Plasma Lipidome Determined by the Lipidyzer Platform. Biomolecules 2021, 11, 326. [Google Scholar] [CrossRef] [PubMed]
- Franko, A.; Merkel, D.; Kovarova, M.; Hoene, M.; Jaghutriz, B.A.; Heni, M.; Königsrainer, A.; Papan, C.; Lehr, S.; Häring, H.U.; et al. Dissociation of Fatty Liver and Insulin Resistance in I148M PNPLA3 Carriers: Differences in Diacylglycerol (DAG) FA18:1 Lipid Species as a Possible Explanation. Nutrients 2018, 10, 1314. [Google Scholar] [CrossRef] [PubMed]
- Ubhi, B.K. Direct Infusion-Tandem Mass Spectrometry (DI-MS/MS) Analysis of Complex Lipids in Human Plasma and Serum Using the Lipidyzer Platform. Clin. Metabolomics Methods Protoc. 2018, 1730, 227–236. [Google Scholar] [CrossRef]
- Liebisch, G.; Fahy, E.; Aoki, J.; Dennis, E.A.; Durand, T.; Ejsing, C.S.; Fedorova, M.; Feussner, I.; Griffiths, W.J.; Köfeler, H.; et al. Update on LIPID MAPS classification, nomenclature, and shorthand notation for MS-derived lipid structures. J. Lipid Res. 2020, 61, 1539–1555. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szász, I.; Koroknai, V.; Várvölgyi, T.; Pál, L.; Szűcs, S.; Pikó, P.; Emri, G.; Janka, E.; Szabó, I.L.; Ádány, R.; et al. Identification of Plasma Lipid Alterations Associated with Melanoma Metastasis. Int. J. Mol. Sci. 2024, 25, 4251. https://doi.org/10.3390/ijms25084251
Szász I, Koroknai V, Várvölgyi T, Pál L, Szűcs S, Pikó P, Emri G, Janka E, Szabó IL, Ádány R, et al. Identification of Plasma Lipid Alterations Associated with Melanoma Metastasis. International Journal of Molecular Sciences. 2024; 25(8):4251. https://doi.org/10.3390/ijms25084251
Chicago/Turabian StyleSzász, István, Viktória Koroknai, Tünde Várvölgyi, László Pál, Sándor Szűcs, Péter Pikó, Gabriella Emri, Eszter Janka, Imre Lőrinc Szabó, Róza Ádány, and et al. 2024. "Identification of Plasma Lipid Alterations Associated with Melanoma Metastasis" International Journal of Molecular Sciences 25, no. 8: 4251. https://doi.org/10.3390/ijms25084251
APA StyleSzász, I., Koroknai, V., Várvölgyi, T., Pál, L., Szűcs, S., Pikó, P., Emri, G., Janka, E., Szabó, I. L., Ádány, R., & Balázs, M. (2024). Identification of Plasma Lipid Alterations Associated with Melanoma Metastasis. International Journal of Molecular Sciences, 25(8), 4251. https://doi.org/10.3390/ijms25084251