
Genome-Wide Identification of Glutathione S-Transferase Genes in Eggplant (Solanum melongena L.) Reveals Their Potential Role in Anthocyanin Accumulation on the Fruit Peel

Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification and Analysis of GST Family Genes in Eggplant
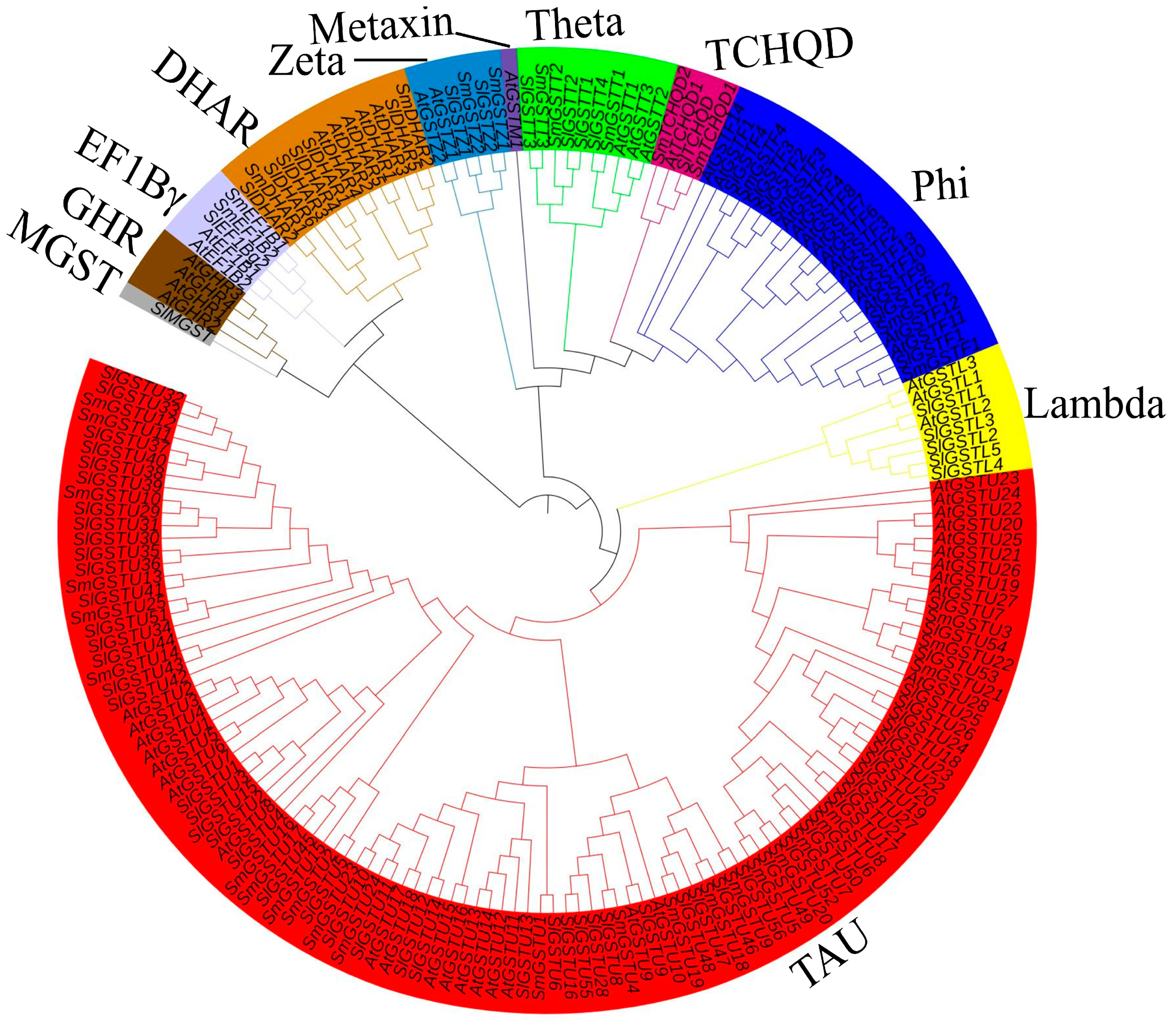
2.2. Phylogenetic Analysis of the GST Family Genes
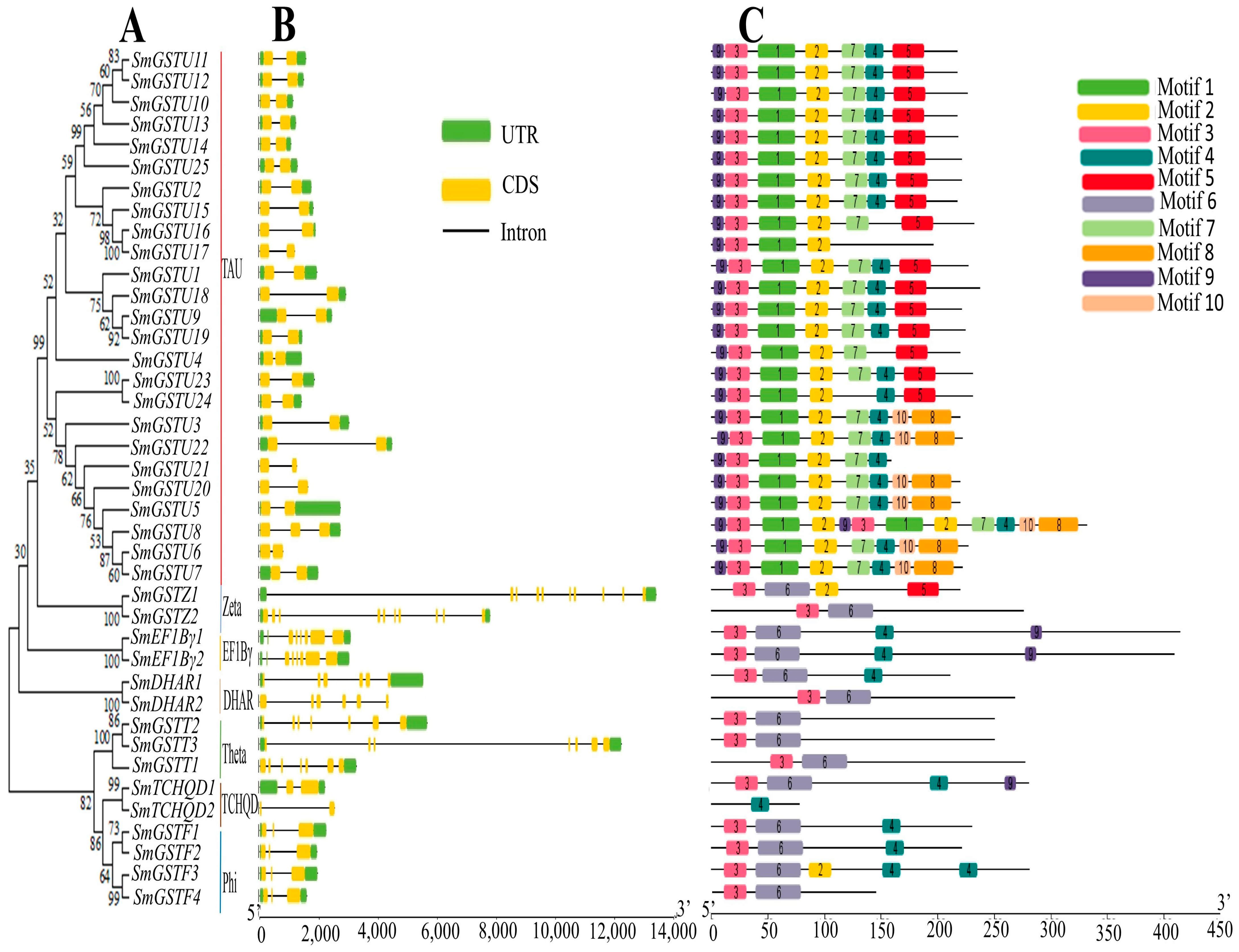
2.3. The Phylogeny, Gene Structure and Motif Composition of the SmGST Genes
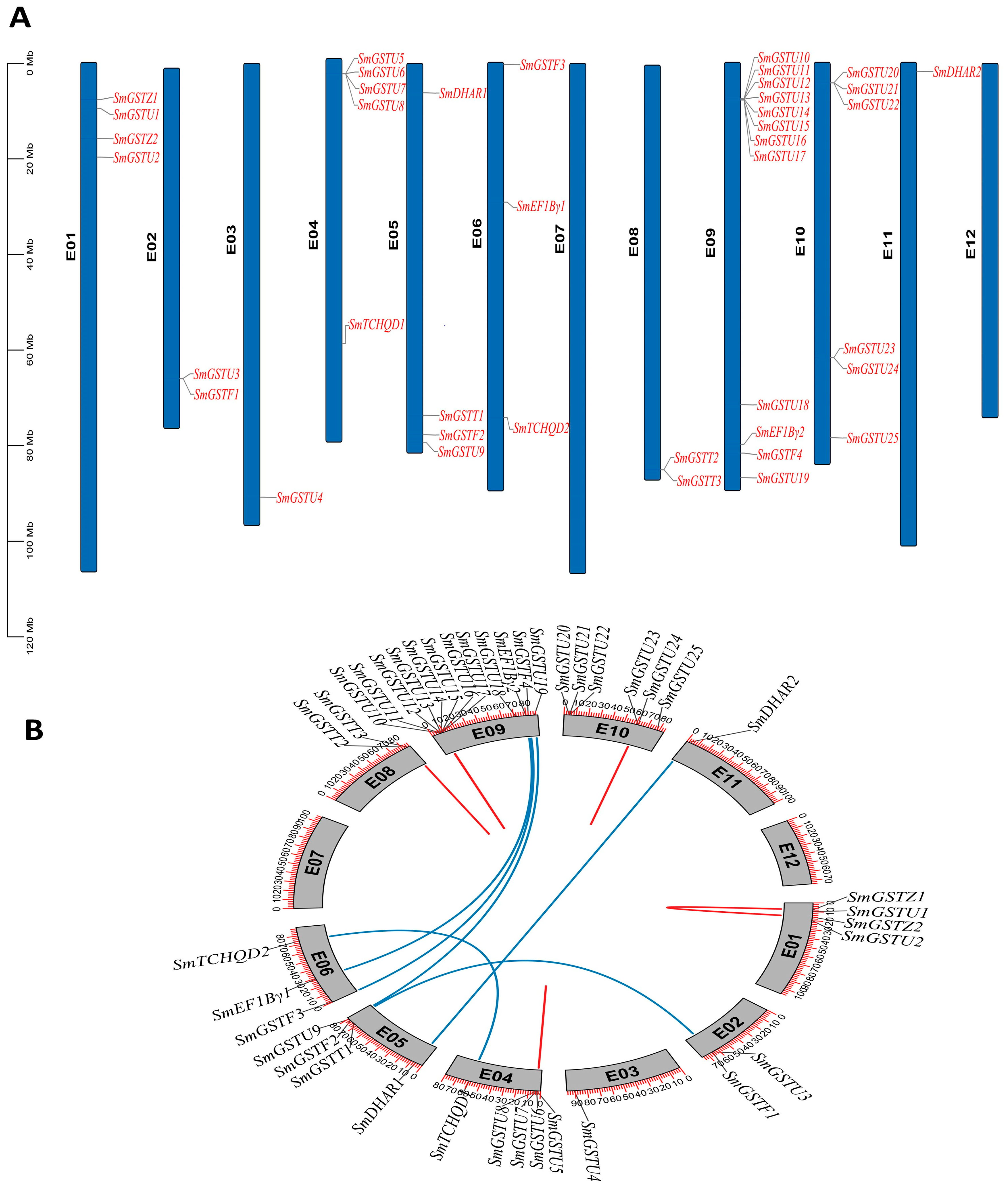
2.4. Distribution of SmGST Genes on the Chromosomes and Their Duplication
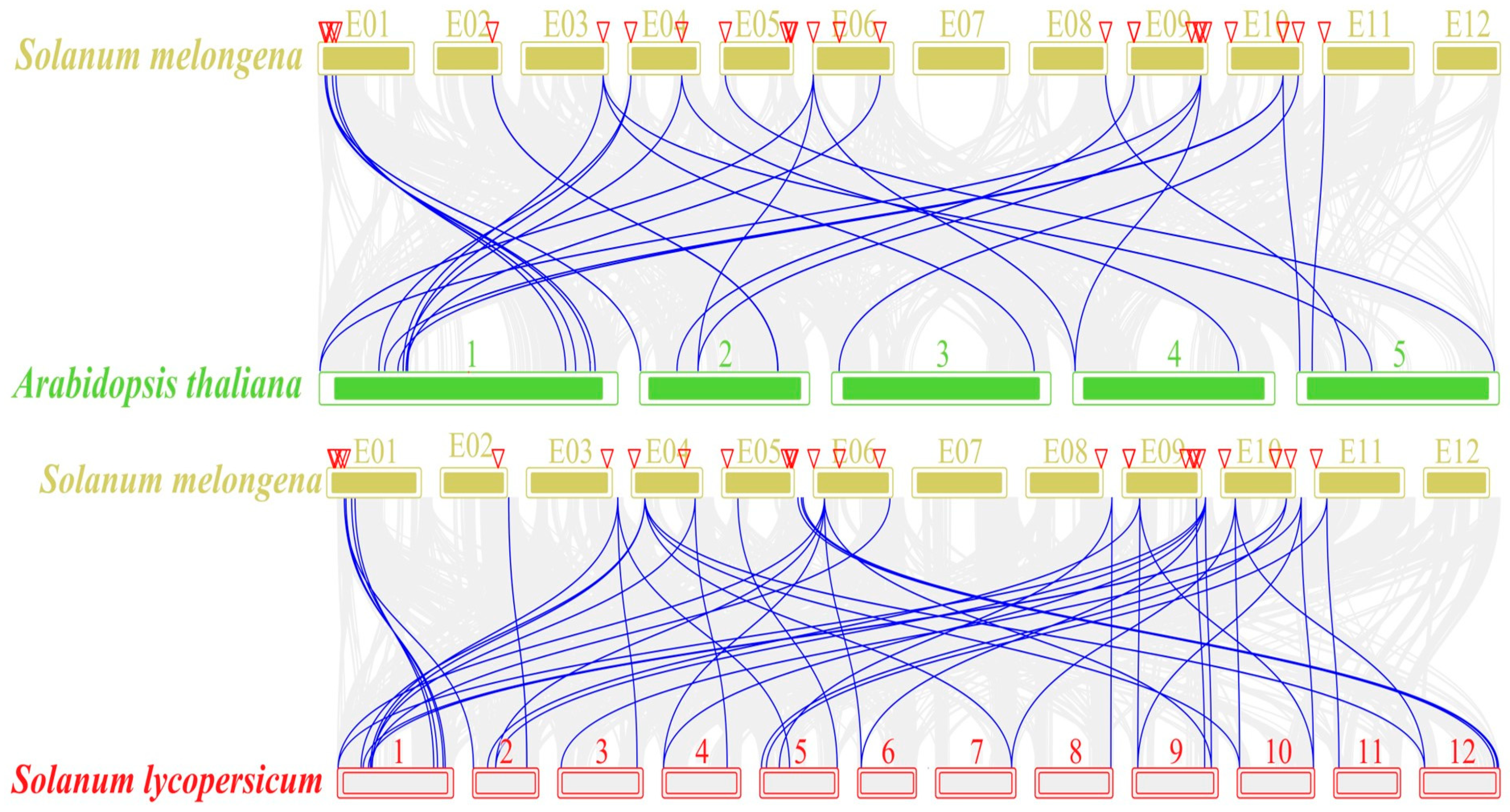
2.5. Collinearity Analysis of the SmGST Genes in Eggplant
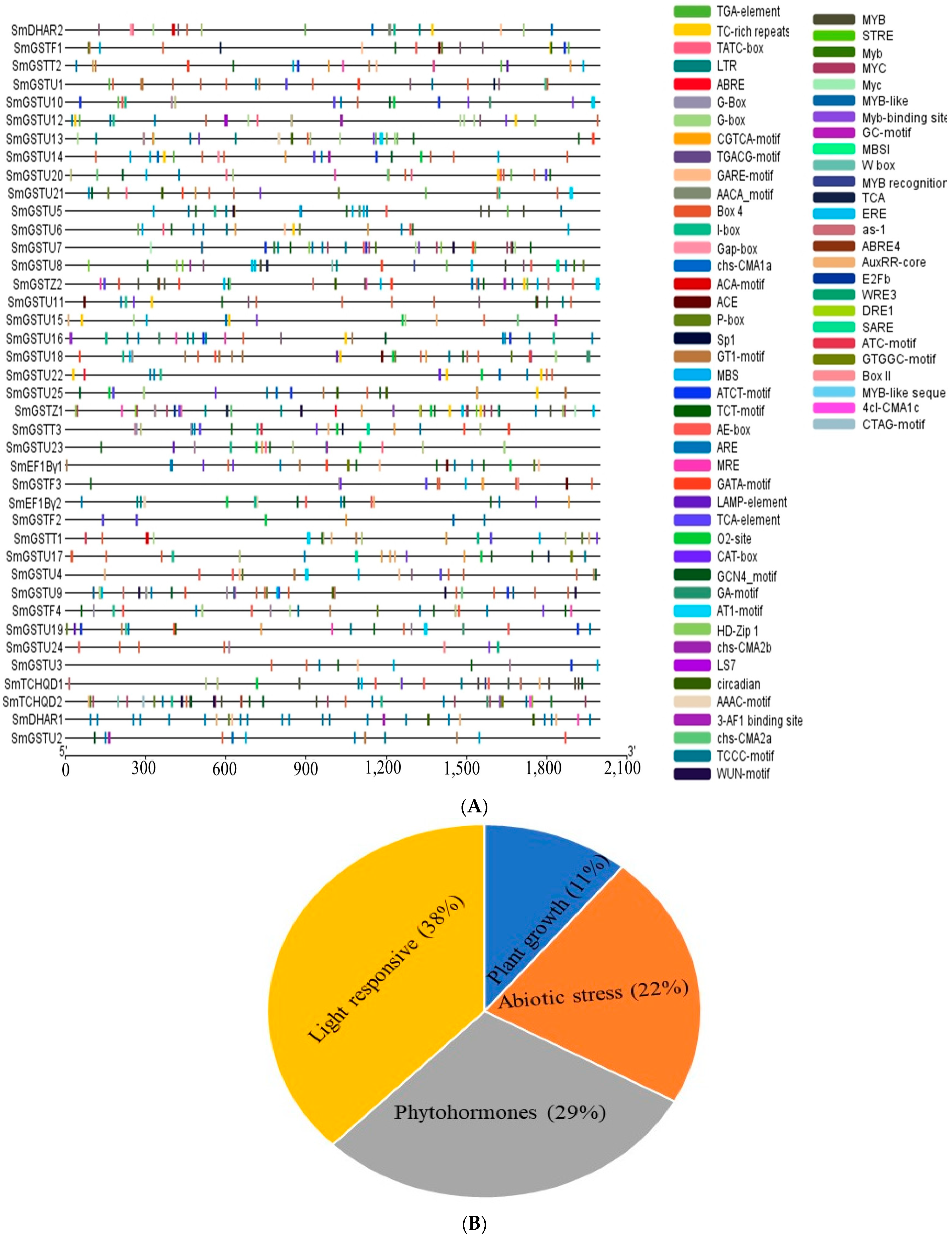
2.6. Analysis of Cis-Acting Elements in the Promoters of SmGST Genes
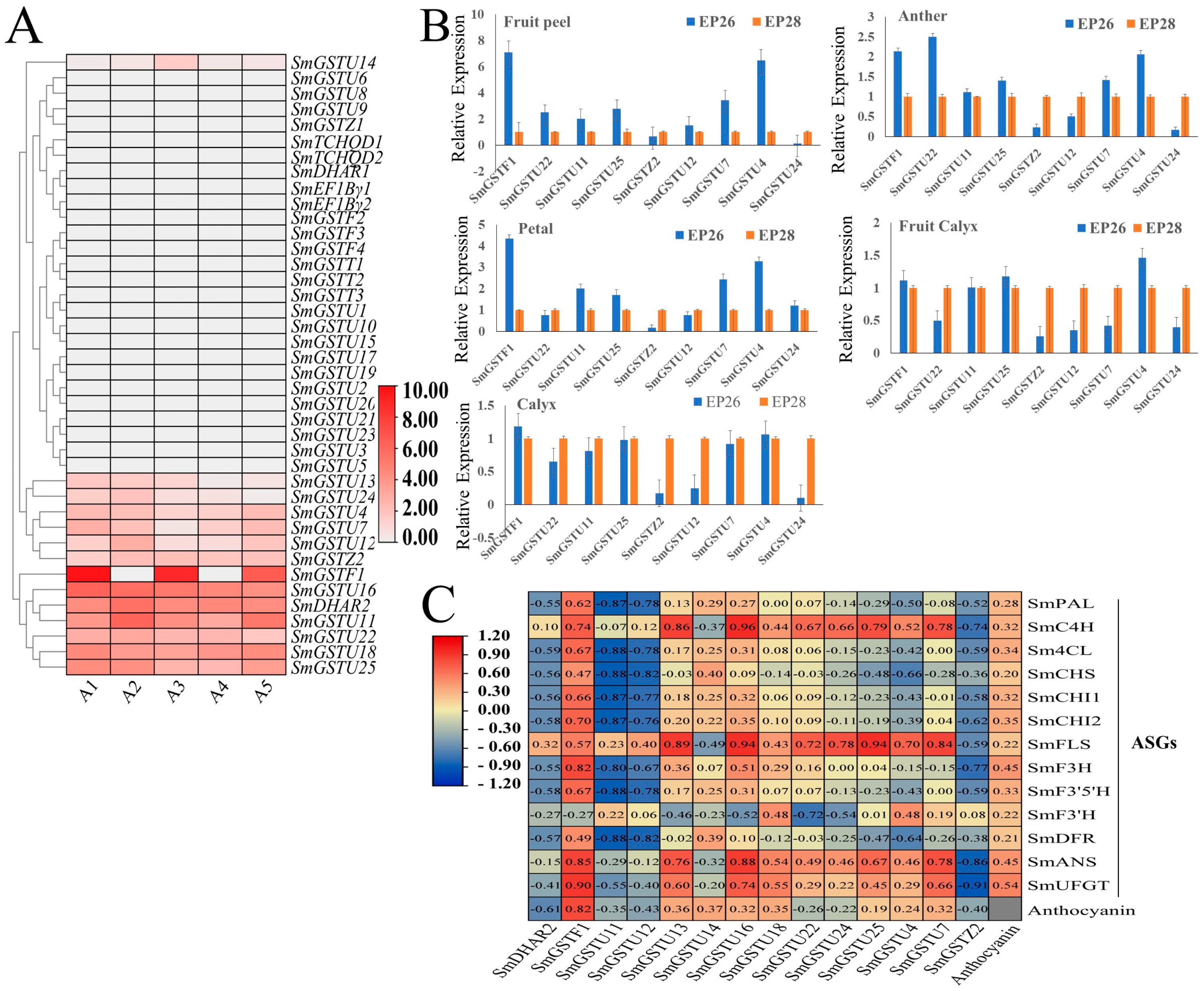
2.7. Expression Patterns of SmGST Genes in the Peel of Different Eggplant Varieties
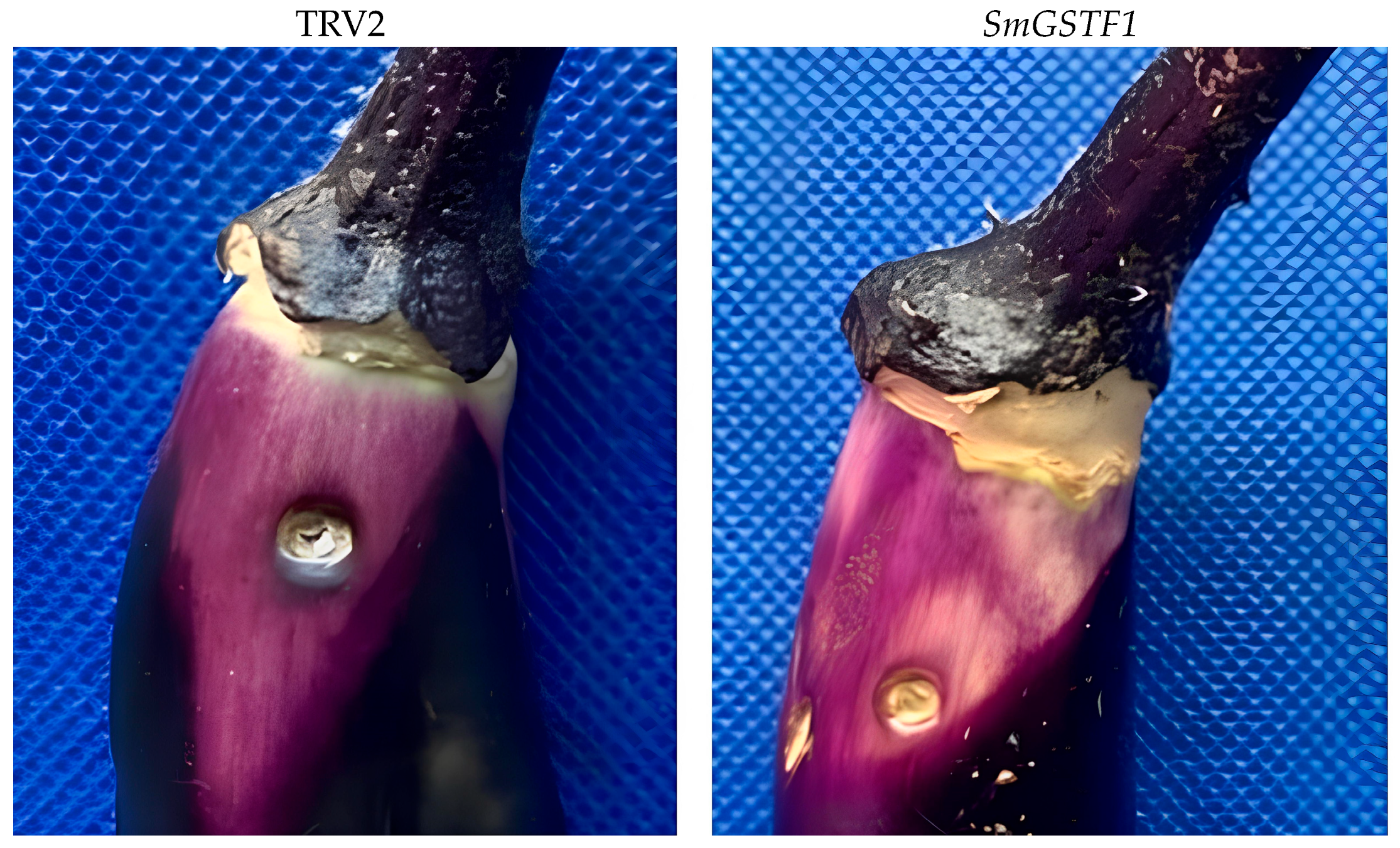
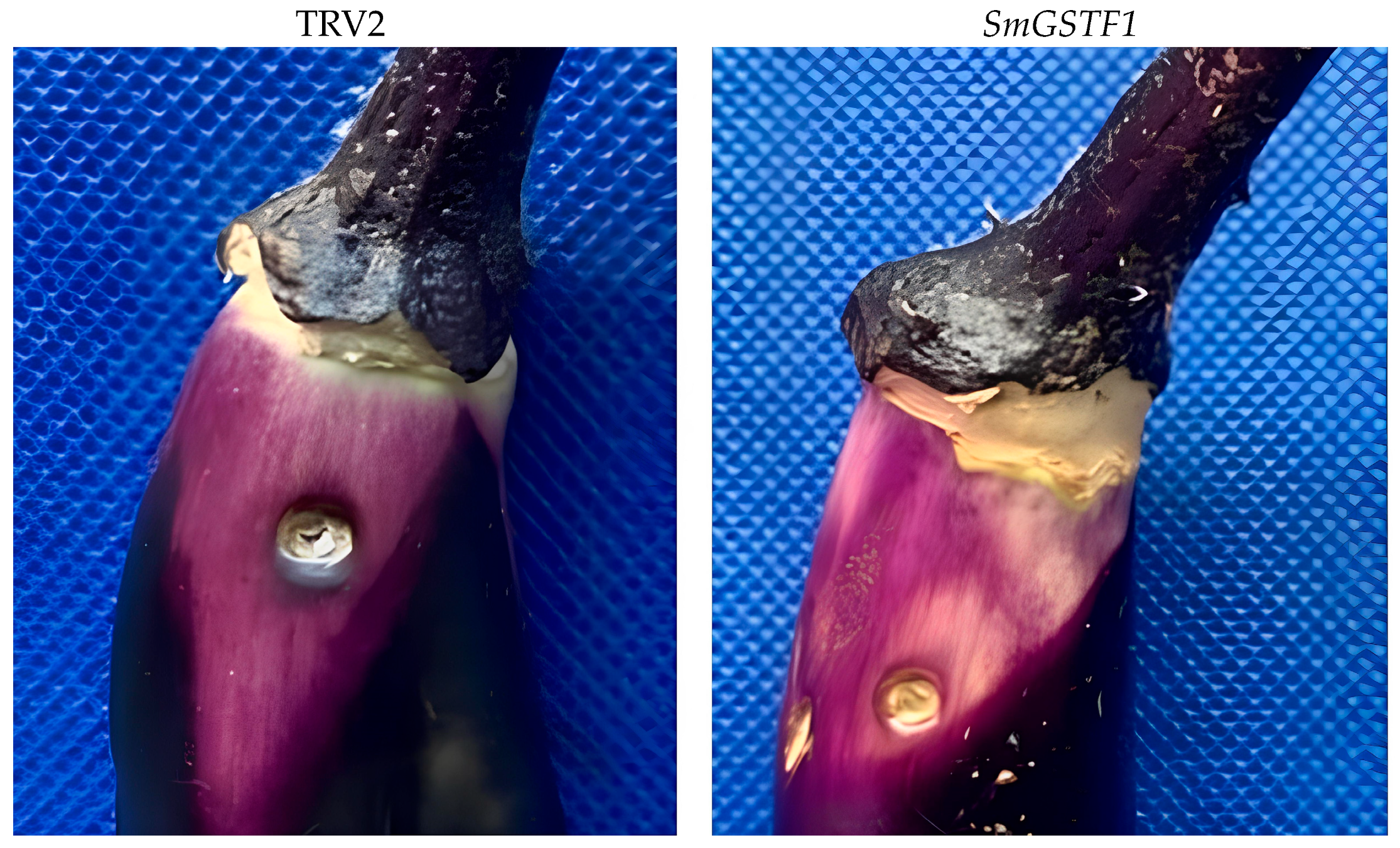
2.8. Effects of Silencing SmGSTF1 on the Eggplant Peel Color
3. Discussion
4. Materials and Methods
4.1. Identification and Characterization of GST Family Genes in Eggplant
4.2. Phylogenetic, Motif and Protein Structure Analyses of GST Genes
4.3. Chromosomal Localization, Gene Duplication and Collinearity Analyses of GST Genes
4.4. Analysis of Cis-Acting Element of SmGST Genes
4.5. Expression Analysis Using Transcriptomic Data
4.6. Total Anthocyanin Analysis
4.7. Vector Construction and Transformation of Eggplant
4.8. Quantitative Real-Time PCR Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, M.; Opoku, K.N.; Bissah, N.A.B.; Su, T. Solanum aethiopicum: The nutrient-rich vegetable crop with great economic, genetic biodiversity and pharmaceutical potential. Horticulturae 2021, 7, 126. [Google Scholar] [CrossRef]
- FAOSTAT. FAO Statistical Database. 2022. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 10 October 2023).
- Sang, Z.; Zuo, J.; Wang, Q.; Fu, A.; Zheng, Y.; Ge, Y.; Qian, Z.; Cui, Y. Determining the effects of light on the fruit peel quality of photosensitive and non-photosensitive eggplant. Plants 2022, 11, 2095. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Wang, P.; Wang, W.; Peng, J.; Zheng, J.; Zhu, G.; Zhong, C.; Yu, W. Fine mapping and identification of SmAPRR2 regulating rindcolor in eggplant (Solanum melongena L.). Int. J. Mol. Sci. 2023, 24, 3059. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, M.A.; Sabir, I.A.; Shah, I.H.; Riaz, M.W.; Rehman, S.; Song, C.; Li, G.; Malik, M.S.; Ashraf, G.A.; Haider, M.S.; et al. Flavonoids: A review on biosynthesis and transportation mechanism in plants. Funct. Integr. Genom. 2023, 23, 212. [Google Scholar] [CrossRef]
- Cerqueira, J.V.A.; de Andrade, M.T.; Rafael, D.D.; Zhu, F.; Martins, S.V.C.; Nunes-Nesi, A.; Benedito, V.; Fernie, A.R.; Zsögön, A. Anthocyanins and reactive oxygen species: A team of rivals regulating plant development? Plant Mol. Biol. 2023, 112, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Paul, I.; Sarkar, M.P.; Bhadoria, P.B.S. Floral secondary metabolites in context of biotic and abiotic stress factors. Chemoecology 2021, 32, 49–68. [Google Scholar] [CrossRef]
- Kaur, S.; Tiwari, V.; Kumari, A.; Chaudhary, E.; Sharma, A.; Ali, U.; Garg, M. Protective and defensive role of anthocyanins under plant abiotic and biotic stresses: An emerging application in sustainable agriculture. J. Biotechnol. 2023, 361, 12–29. [Google Scholar] [CrossRef]
- Zhuang, W.-B.; Li, Y.-H.; Shu, X.-C.; Pu, Y.-T.; Wang, X.-J.; Wang, T.; Wang, Z. The classification, molecular structure and biological biosynthesis of flavonoids and their roles in biotic and abiotic stresses. Molecules 2023, 28, 3599. [Google Scholar] [CrossRef]
- Rahaman, M.; Hossain, R.; Herrera-Bravo, J.; Islam, M.T.; Atolani, O.; Adeyemi, O.S.; Owolodun, O.A.; Kambizi, L.; Daştan, S.D.; Calina, D.; et al. Natural antioxidants from some fruits, seeds, foods, natural products and associated health benefits: An update. Food Sci. Nutr. 2023, 11, 1657–1670. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, B.; Wu, T.; Yang, Y.; Fan, L.; Wen, M.; Sui, J. Transcriptomic profiling of two Pak Choi varieties with contrasting anthocyanin contents provides an insight into structural and regulatory genes in anthocyanin biosynthetic pathway. BMC Genom. 2017, 18, 288. [Google Scholar] [CrossRef]
- Naing, A.H.; Kil Kim, C. Roles of R2R3-MYB transcription factors in transcriptional regulation of anthocyanin biosynthesis in horticultural plants. Plant Mol. Biol. 2018, 98, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Silva, S.; dos Santos, A.L.; Chalfun-Júnior, A.; Zhao, J.; Peres, L.E.; Benedito, V.A. Understanding the genetic regulation of anthocyanin biosynthesis in plants—Tools for breeding purple varieties of fruits and vegetables. Phytochemistry 2018, 153, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.-D.; Wang, C.-K.; Hu, D.-G.; Hao, Y.-J. How do anthocyanins paint our horticultural products? Sci. Hortic. 2019, 249, 257–262. [Google Scholar] [CrossRef]
- Rahim, A.; Zhang, X.; Busatto, N. Editorial: Phenylpropanoid biosynthesis in plants. Front. Plant Sci. 2023, 14, 1230664. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Song, Y.; Lin, J.; Dixon, R.A. The complexities of proanthocyanidin biosynthesis and its regulation in plants. Plant Commun. 2023, 4, 100498. [Google Scholar] [CrossRef]
- Chen, L.; Hu, B.; Qin, Y.; Hu, G.; Zhao, J. Advance of the negative regulation of anthocyanin biosynthesis by MYB transcription factors. Plant Physiol. Biochem. 2019, 136, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Gomez, C.; Conejero, G.; Torregrosa, L.; Cheynier, V.; Terrier, N.; Ageorges, A. In vivo grapevine anthocyanin transport involves vesicle-mediated trafficking and the contribution of anthoMATE transporters and GST. Plant J. 2011, 67, 960–970. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J. Flavonoid transport mechanisms: How to go and with whom. Trends Plant Sci. 2015, 20, 576–585. [Google Scholar] [CrossRef]
- Kaur, S.; Sharma, N.; Kapoor, P.; Chunduri, V.; Pandey, A.K.; Garg, M. Spotlight on the overlapping routes and partners for anthocyanin transport in plants. Physiol. Plant. 2021, 171, 868–881. [Google Scholar] [CrossRef]
- Moons, A. regulatory and functional interactions of plant growth regulators and plant glutathione S-transferases (GSTs). Vitam. Horm. 2005, 72, 155–202. [Google Scholar] [CrossRef]
- Vaish, S.; Gupta, D.; Mehrotra, R.; Mehrotra, S.; Basantani, M.K. Glutathione S-transferase: A versatile protein family. 3 Biotech 2020, 10, 321. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Zhou, F.; Shan, C.; Zhang, Q.; Ning, M.; Liu, X.; Zhao, X.; Cai, W.; Yang, X.; Hao, G.; et al. Identification of glutathione S-transferase genes in hami melon (Cucumis melo var. saccharinus) and their expression analysis under cold stress. Front. Plant Sci. 2021, 12, 672017. [Google Scholar] [CrossRef]
- Shao, D.; Li, Y.; Zhu, Q.; Zhang, X.; Liu, F.; Xue, F.; Sun, J. GhGSTF12, a glutathione S-transferase gene, is essential for anthocyanin accumulation in cotton (Gossypium hirsutum L.). Plant Sci. 2021, 305, 110827. [Google Scholar] [CrossRef] [PubMed]
- Nianiou-Obeidat, I.; Madesis, P.; Kissoudis, C.; Voulgari, G.; Chronopoulou, E.; Tsaftaris, A.; Labrou, N.E. Plant glutathione transferase-mediated stress tolerance: Functions and biotechnological applications. Plant Cell Rep. 2017, 36, 791–805. [Google Scholar] [CrossRef]
- Mano, J.; Kanameda, S.; Kuramitsu, R.; Matsuura, N.; Yamauchi, Y. Detoxification of reactive carbonyl species by glutathione transferase tau isozymes. Front. Plant Sci. 2019, 10, 487. [Google Scholar] [CrossRef] [PubMed]
- Sylvestre-Gonon, E.; Schwartz, M.; Girardet, J.-M.; Hecker, A.; Rouhier, N. Is there a role for tau glutathione transferases in tetrapyrrole metabolism and retrograde signalling in plants? Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190404. [Google Scholar] [CrossRef] [PubMed]
- Holm, P.J.; Bhakat, P.; Jegerschöld, C.; Gyobu, N.; Mitsuoka, K.; Fujiyoshi, Y.; Morgenstern, R.; Hebert, H. Structural basis for detoxification and oxidative stress protection in membranes. J. Mol. Biol. 2006, 360, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Basantani, M.; Srivastava, A. Plant glutathione transferases—A decade falls short. Can. J. Bot. 2007, 85, 443–456. [Google Scholar] [CrossRef]
- Dixon, D.P.; Lapthorn, A.; Edwards, R. Plant glutathione transferases. Genome Biol. 2002, 3, Reviews3004.1. [Google Scholar] [CrossRef]
- Lallement, P.-A.; Brouwer, B.; Keech, O.; Hecker, A.; Rouhier, N. The still mysterious roles of cysteine-containing glutathione transferases in plants. Front. Pharmacol. 2014, 5, 192. [Google Scholar] [CrossRef]
- Sylvestre-Gonon, E.; Law, S.R.; Schwartz, M.; Robe, K.; Keech, O.; Didierjean, C.; Dubos, C.; Rouhier, N.; Hecker, A. Functional, structural and biochemical features of plant serinyl-glutathione transferases. Front. Plant Sci. 2019, 10, 608. [Google Scholar] [CrossRef] [PubMed]
- Vaish, S.; Parveen, R.; Gupta, D.; Basantani, M.K. Genome-wide identification and characterization of glutathione S-transferase gene family in Musa acuminata L. AAA group and gaining an insight to their role in banana fruit development. J. Appl. Genet. 2022, 63, 609–631. [Google Scholar] [CrossRef] [PubMed]
- Vaish, S.; Parveen, R.; Rajneesh; Singh, N.; Gupta, D.; Basantani, M.K. Computational insights into diverse aspects of glutathione S-transferase gene family in Papaver somniferum. J. Plant Res. 2022, 135, 823–852. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, H.; Huang, J.-R. Arabidopsis TT19 functions as a carrier to transport anthocyanin from the cytosol to tonoplasts. Mol. Plant 2012, 5, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Díaz, R.; Madrid-Espinoza, J.; Salinas-Cornejo, J.; González-Villanueva, E.; Ruiz-Lara, S. Differential roles for VviGST1, VviGST3, and VviGST4 in proanthocyanidin and anthocyanin transport in Vitis vinífera. Front. Plant Sci. 2016, 7, 1166. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Zhao, J.; Lai, B.; Qin, Y.; Wang, H.; Hu, G. LcGST4 is an anthocyanin-related glutathione S-transferase gene in Litchi chinensis Sonn. Plant Cell Rep. 2016, 35, 831–843. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-W.; Wang, C.-K.; Huang, X.-Y.; Hu, D.-G. Genome-wide analysis of the glutathione S-transferase (GST) genes and functional identification of MdGSTU12 reveals the involvement in the regulation of anthocyanin accumulation in apple. Genes 2021, 12, 1733. [Google Scholar] [CrossRef]
- Jiang, S.; Chen, M.; He, N.; Chen, X.; Wang, N.; Sun, Q.; Zhang, T.; Xu, H.; Fang, H.; Wang, Y.; et al. MdGSTF6, activated by MdMYB1, plays an essential role in anthocyanin accumulation in apple. Hortic. Res. 2019, 6, 40. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qi, Y.; Zhang, A.; Wu, H.; Liu, Z.; Ren, X. Molecular cloning and functional characterization of AcGST1, an anthocyanin-related glutathione S-transferase gene in kiwifruit (Actinidia chinensis). Plant Mol. Biol. 2019, 100, 451–465. [Google Scholar] [CrossRef]
- Han, L.; Zhou, L.; Zou, H.; Yuan, M.; Wang, Y. PsGSTF3, an anthocyanin-related glutathione s-transferase gene, is essential for petal coloration in tree peony. Int. J. Mol. Sci. 2022, 23, 1423. [Google Scholar] [CrossRef]
- Wei, K.; Wang, L.; Zhang, Y.; Ruan, L.; Li, H.; Wu, L.; Xu, L.; Zhang, C.; Zhou, X.; Cheng, H.; et al. A coupled role for CsMYB75 and CsGSTF1 in anthocyanin hyperaccumulation in purple tea. Plant J. 2018, 97, 825–840. [Google Scholar] [CrossRef]
- Li, B.; Zhang, X.; Duan, R.; Han, C.; Yang, J.; Wang, L.; Wang, S.; Su, Y.; Wang, L.; Dong, Y.; et al. Genomic analysis of the glutathione S-transferase family in pear (Pyrus communis) and functional identification of PcGST57 in anthocyanin accumulation. Int. J. Mol. Sci. 2022, 23, 746. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Liu, S.; Yang, Y.; Liu, J.; Zhuang, Y. Integrated metabolome and transcriptome analysis reveals a regulatory network of fruit peel pigmentation in eggplant (Solanum melongena L.). Int. J. Mol. Sci. 2022, 23, 13475. [Google Scholar] [CrossRef] [PubMed]
- Monticolo, F.; Colantuono, C.; Chiusano, M.L. Shaping the evolutionary tree of green plants: Evidence from the GST family. Sci. Rep. 2017, 7, 14363. [Google Scholar] [CrossRef]
- Sappl, P.G.; Carroll, A.J.; Clifton, R.; Lister, R.; Whelan, J.; Millar, A.H.; Singh, K.B. The Arabidopsis glutathione transferase gene family displays complex stress regulation and co-silencing multiple genes results in altered metabolic sensitivity to oxidative stress. Plant J. 2009, 58, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Chen, B.; Lin, H.; Liu, Y.; Wei, Y.; Chen, F.; Li, W. Identification and characterization of the glutathione S-Transferase (GST) family in radish reveals a likely role in anthocyanin biosynthesis and heavy metal stress tolerance. Gene 2020, 743, 144484. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Choudhury, M.; Majlish, A.-N.K.; Islam, T.; Ghosh, A. Comprehensive genome-wide analysis of Glutathione S-transferase gene family in potato (Solanum tuberosum L.) and their expression profiling in various anatomical tissues and perturbation conditions. Gene 2018, 639, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Rahman, I.A.; Islam, T.; Ghosh, A. Genome-wide identification and expression analysis of glutathione S-transferase gene family in tomato: Gaining an insight to their physiological and stress-specific roles. PLoS ONE 2017, 12, e0187504. [Google Scholar] [CrossRef]
- Ahmad, M.Z.; Nasir, J.A.; Ahmed, S.; Ahmad, B.; Sana, A.; Salman, S.; Shah, Z.; Yang, C. Genome-wide analysis of glutathione S-transferase gene family in G. max. Biologia 2020, 75, 1691–1705. [Google Scholar] [CrossRef]
- Islam, S.; Sajib, S.D.; Jui, Z.S.; Arabia, S.; Islam, T.; Ghosh, A. Genome-wide identification of glutathione S-transferase gene family in pepper, its classification, and expression profiling under different anatomical and environmental conditions. Sci. Rep. 2019, 9, 9101. [Google Scholar] [CrossRef]
- Li, H.; Yang, Y.; Li, H.; Wang, W.; Zheng, H.; Tao, J. Genome-wide identification of glutathione s-transferase and expression analysis in response to anthocyanin transport in the flesh of the new teinturier grape germplasm ‘Zhongshan-HongYu’. Int. J. Mol. Sci. 2022, 23, 7717. [Google Scholar] [CrossRef] [PubMed]
- Mo, Z.; Huang, Y.; Pu, T.; Duan, L.; Pi, K.; Luo, J.; Long, B.; Lu, A.; Liu, R. Genome-wide identification and characterization of glutathione S-transferases (GSTs) and their expression profile under abiotic stresses in tobacco (Nicotiana tabacum L.). BMC Genom. 2023, 24, 341. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.S.; Raes, J. Duplication and Divergence: The evolution of new genes and old ideas. Annu. Rev. Genet. 2004, 38, 615–643. [Google Scholar] [CrossRef]
- Sasan, M.; Maryam, E.; Fateme, M.; Maryam, S.; Babak, S.; Hassan, M. Plant glutathione S-transferase classification, structure and evolution. Afr. J. Biotechnol. 2011, 10, 8160–8165. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, Y.; Zhang, W.; Yang, S.; Chen, J.-Q.; Tian, D. Patterns of exon-intron architecture variation of genes in eukaryotic genomes. BMC Genom. 2009, 10, 47. [Google Scholar] [CrossRef]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exon–intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef]
- Singh, P.; Ahi, E.P. The importance of alternative splicing in adaptive evolution. Mol. Ecol. 2022, 31, 1928–1938. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, Z.; Chu, G.; Huang, C.; Tian, S.; Zhao, Z.; Chen, G. Anthocyanin accumulation and molecular analysis of anthocyanin biosynthesis-associated genes in eggplant (Solanum melongena L.). J. Agric. Food Chem. 2014, 62, 2906–2912. [Google Scholar] [CrossRef]
- Li, S.; Zuo, D.; Cheng, H.; Ali, M.; Wu, C.; Ashraf, J.; Zhang, Y.; Feng, X.; Lin, Z.; Wang, Q.; et al. Glutathione S-transferases GhGSTF1 and GhGSTF2 involved in the anthocyanin accumulation in Gossypium hirsutum L. Int. J. Biol. Macromol. 2020, 165, 2565–2575. [Google Scholar] [CrossRef]
- Niu, M.; Bao, C.; Chen, J.; Zhou, W.; Zhang, Y.; Zhang, X.; Su, N.; Cui, J. RsGSTF12 Contributes to anthocyanin sequestration in radish (Raphanus sativus L.). Front. Plant Sci. 2022, 13, 870202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, Y.; Zhang, M.; Cai, F.; Qin, J.; Wang, Q.; Liu, C.; Wang, G.; Xu, L.; Yang, L.; et al. Molecular cloning and functional characterization of a glutathione S-transferase involved in both anthocyanin and proanthocyanidin accumulation in Camelina sativa (Brassicaceae). Evolution 2012, 11, 4711–4719. [Google Scholar] [CrossRef]
- Zhang, Z.; Tian, C.; Zhang, Y.; Li, C.; Li, X.; Yu, Q.; Wang, S.; Wang, X.; Chen, X.; Feng, S. Transcriptomic and metabolomic analysis provides insights into anthocyanin and procyanidin accumulation in pear. BMC Plant Biol. 2020, 20, 129. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, w202–w208. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Koch, M.A.; Haubold, B.; Mitchell-Olds, T. Comparative evolutionary analysis of chalcone synthase and alcohol dehydrogenase loci in Arabidopsis, Arabis and related genera (Brassicaceae). Mol. Biol. Evol. 2000, 17, 1483–1498. [Google Scholar] [CrossRef]
- He, Q.; Zhang, Z.F.; Zhang, L.G. Anthocyanin accumulation, antioxidant ability and stability and a transcriptional analysis of anthocyanin biosynthesis in purple heading chinese cabbage (Brassica rapa L. ssp. pekinensis). J. Agric. Food Chem. 2016, 64, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Romero, I.; Tikunov, Y.; Bovy, A. Virus-induced gene silencing in detached tomatoes and biochemical effects of phytoene desaturase gene silencing. J. Plant Physiol. 2011, 168, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Gene Name | Gene Id | Chr | Start-End | Cds | AA | mW (kDA) | pI | Sub. Loc. |
|---|---|---|---|---|---|---|---|---|---|
| TAU | SmGSTU1 | Smechr0101012 | E01 | 9,617,214–9,619,141 | 1928 | 227 | 25.664 | 5.18 | cytoplasm |
| SmGSTU2 | Smechr0101935 | E01 | 19,836,231–19,837,973 | 1743 | 221 | 25.269 | 6.46 | cytoplasm | |
| SmGSTU3 | Smechr0201984 | E02 | 64,921,863–64,924,878 | 3016 | 220 | 25.402 | 5.3 | cytoplasm | |
| SmGSTU4 | Smechr0303071 | E03 | 90,785,487–90,786,908 | 1422 | 220 | 25.326 | 5.52 | cytoplasm | |
| SmGSTU5 | Smechr0400284 | E04 | 3,173,220–3,175,948 | 2729 | 220 | 26.052 | 6.23 | cytoplasm | |
| SmGSTU6 | Smechr0400285 | E04 | 3,176,679–3,177,460 | 782 | 227 | 26.695 | 5.33 | cytoplasm | |
| SmGSTU7 | Smechr0400287 | E04 | 3,180,873–3,182,848 | 1976 | 222 | 25.764 | 5.91 | cytoplasm | |
| SmGSTU8 | Smechr0400288 | E04 | 3,182,978–3,185,708 | 2731 | 332 | 38.905 | 5.67 | cytoplasm | |
| SmGSTU9 | Smechr0502567 | E05 | 79,384,784–79,387,225 | 2442 | 221 | 25.895 | 5.51 | chloroplast | |
| SmGSTU10 | Smechr0900514 | E09 | 7,607,904–7,609,024 | 1121 | 226 | 26.289 | 7.6 | chloroplast | |
| SmGSTU11 | Smechr0900520 | E09 | 7,650,862–7,652,420 | 1559 | 217 | 25.204 | 5.59 | cytoplasm | |
| SmGSTU12 | Smechr0900521 | E09 | 7,666,984–7,668,465 | 1482 | 217 | 25.287 | 5.17 | chloroplast | |
| SmGSTU13 | Smechr0900522 | E09 | 7,693,565–7,694,619 | 1055 | 218 | 24.828 | 5.58 | cytoplasm | |
| SmGSTU14 | Smechr0900523 | E09 | 7,693,565–7,694,619 | 1055 | 218 | 24.828 | 5.58 | cytoplasm | |
| SmGSTU15 | Smechr0900526 | E09 | 7,849,254–7,851,059 | 1806 | 217 | 25.070 | 6.91 | cytoplasm | |
| SmGSTU16 | Smechr0900527 | E09 | 7,866,824–7,868,709 | 1886 | 232 | 26.968 | 5.18 | cytoplasm | |
| SmGSTU17 | Smechr0900529 | E09 | 7,890,677–7,891,855 | 1179 | 196 | 22.930 | 7.64 | nucleus | |
| SmGSTU18 | Smechr0901584 | E09 | 71,621,469–71,624,378 | 2910 | 237 | 27.918 | 5.6 | cytoplasm | |
| SmGSTU19 | Smechr0902464 | E09 | 86,899,097–86,900,533 | 1437 | 224 | 26.242 | 5.34 | cytoplasm | |
| SmGSTU20 | Smechr1000365 | E10 | 4,291,432–4,293,058 | 1627 | 220 | 25.588 | 5.55 | cytoplasm | |
| SmGSTU21 | Smechr1000366 | E10 | 4,296,416–4,297,666 | 1251 | 159 | 18.369 | 5.52 | nucleus | |
| SmGSTU22 | Smechr1000367 | E10 | 4,304,841–4,309,315 | 4475 | 222 | 26.047 | 5.85 | nucleus | |
| SmGSTU23 | Smechr1001526 | E10 | 61,779,912–61,781,751 | 1840 | 231 | 25.955 | 6.18 | cytoplasm | |
| SmGSTU24 | Smechr1001527 | E10 | 61,782,434–61,783,844 | 1411 | 231 | 25.832 | 5.2 | cytoplasm | |
| SmGSTU25 | Smechr1002369 | E10 | 78,540,197–78,541,469 | 1273 | 221 | 25.308 | 5.46 | cytoplasm | |
| Phi | SmGSTF1 | Smechr0201996 | E02 | 64,999,172–65,001,420 | 2249 | 230 | 26.330 | 5.87 | chloroplast |
| SmGSTF2 | Smechr0502407 | E05 | 77,720,819–77,722,753 | 1935 | 221 | 25.204 | 5.57 | chloroplast | |
| SmGSTF3 | Smechr0600022 | E06 | 434,013–435,961 | 1949 | 281 | 31.636 | 7.81 | chloroplast | |
| SmGSTF4 | Smechr0902065 | E09 | 81,756,012–81,757,597 | 1586 | 145 | 16.400 | 4.92 | chloroplast | |
| Theta | SmGSTT1 | Smechr0502113 | E05 | 73,646,479–73,649,739 | 3261 | 277 | 31.134 | 6.63 | cytoplasm |
| SmGSTT2 | Smechr0802345 | E08 | 84,625,796–84,631,454 | 5659 | 250 | 28.457 | 9.35 | cytoplasm | |
| SmGSTT3 | Smechr0802346 | E08 | 84,631,786–84,644,013 | 12,228 | 250 | 28.676 | 9.45 | cytoplasm | |
| EF1Bγ | SmEF1Bγ1 | Smechr0600694 | E06 | 29,261,081–29,264,158 | 3078 | 414 | 47.144 | 5.93 | cytoplasm |
| SmEF1Bγ2 | Smechr0901946 | E09 | 79,912,280–79,915,304 | 3025 | 409 | 46.476 | 6.09 | chloroplast | |
| DHAR | SmDHAR1 | Smechr0500548 | E05 | 6,181,924–6,187,450 | 5527 | 211 | 23.544 | 6.6 | cytoplasm |
| SmDHAR2 | Smechr1100162 | E11 | 1,888,515–1,892,865 | 4351 | 268 | 30.058 | 6.14 | chloroplast | |
| TCHQD | SmTCHQD1 | Smechr0401375 | E04 | 59,614,117–59,616,319 | 2203 | 280 | 32.854 | 8.99 | cytoplasm |
| SmTCHQD2 | Smechr0601694 | E06 | 74,293,706–74,296,233 | 2528 | 78 | 9.166 | 4.4 | chloroplast | |
| Zeta | SmGSTZ1 | Smechr0100825 | E01 | 7,795,166–7,808,572 | 13,407 | 220 | 24.872 | 6.32 | cytoplasm |
| SmGSTZ2 | Smechr0101620 | E01 | 15,928,247–15,936,036 | 7790 | 276 | 31.585 | 6.61 | chloroplast |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obel, H.O.; Zhou, X.; Liu, S.; Yang, Y.; Liu, J.; Zhuang, Y. Genome-Wide Identification of Glutathione S-Transferase Genes in Eggplant (Solanum melongena L.) Reveals Their Potential Role in Anthocyanin Accumulation on the Fruit Peel. Int. J. Mol. Sci. 2024, 25, 4260. https://doi.org/10.3390/ijms25084260
Obel HO, Zhou X, Liu S, Yang Y, Liu J, Zhuang Y. Genome-Wide Identification of Glutathione S-Transferase Genes in Eggplant (Solanum melongena L.) Reveals Their Potential Role in Anthocyanin Accumulation on the Fruit Peel. International Journal of Molecular Sciences. 2024; 25(8):4260. https://doi.org/10.3390/ijms25084260
Chicago/Turabian StyleObel, Hesbon Ochieng, Xiaohui Zhou, Songyu Liu, Yan Yang, Jun Liu, and Yong Zhuang. 2024. "Genome-Wide Identification of Glutathione S-Transferase Genes in Eggplant (Solanum melongena L.) Reveals Their Potential Role in Anthocyanin Accumulation on the Fruit Peel" International Journal of Molecular Sciences 25, no. 8: 4260. https://doi.org/10.3390/ijms25084260