
Cardioprotection and Suppression of Fibrosis by Diverse Cancer and Non-Cancer Cell Lines in a Murine Model of Duchenne Muscular Dystrophy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
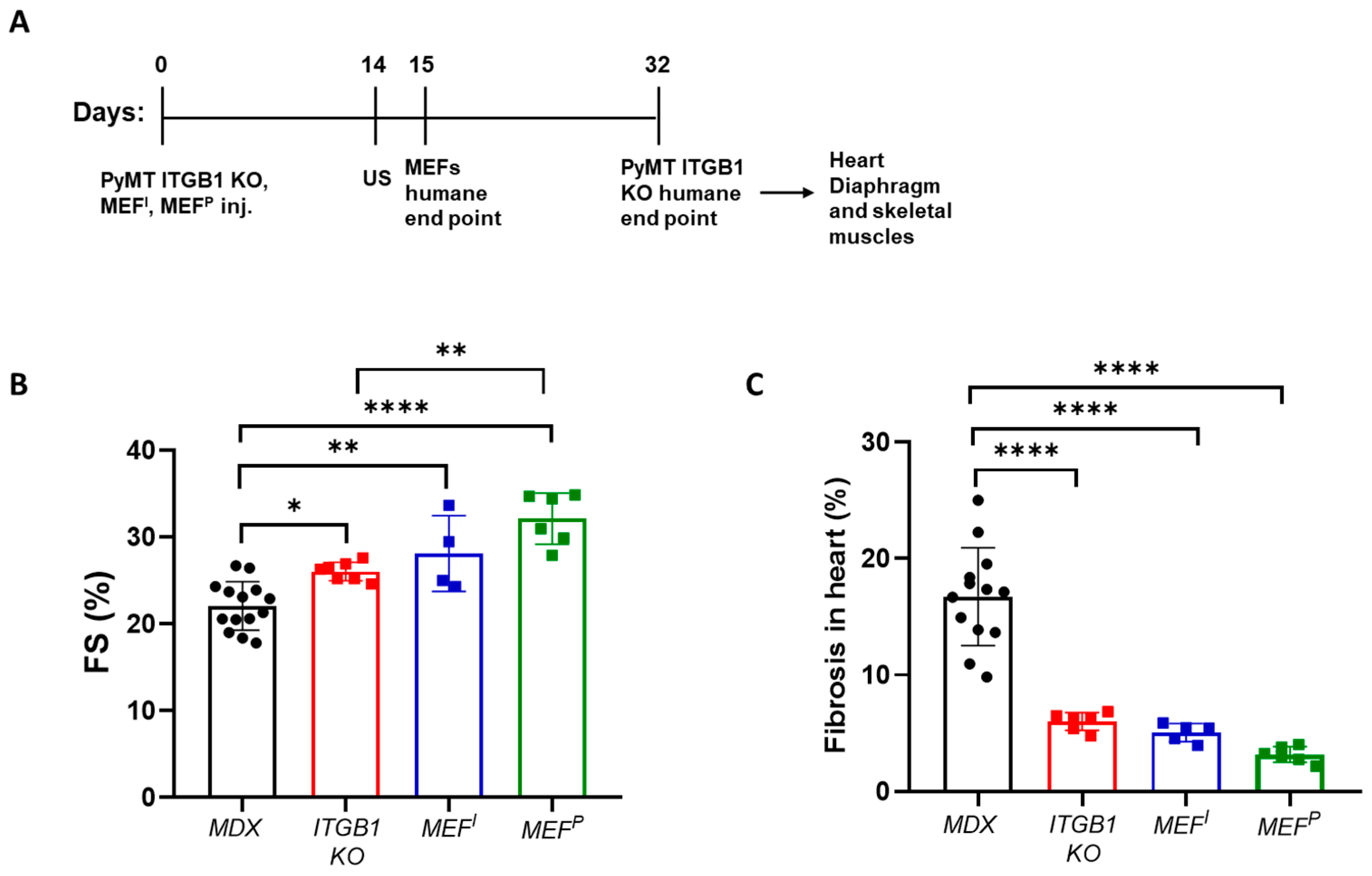
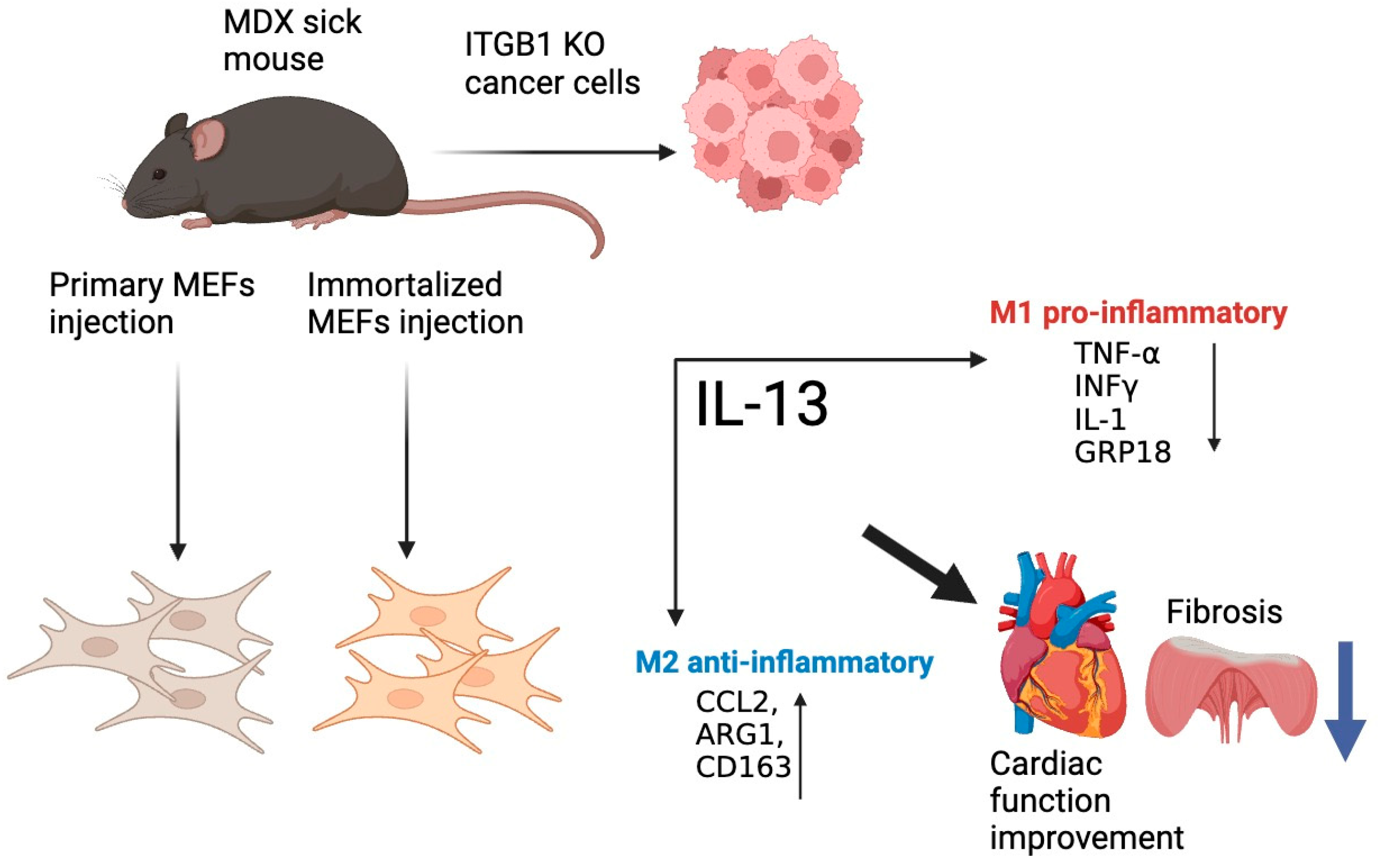
2.1. PyMT ITGB1 KO, MEFI, and MEFP Cells Ameliorate Heart Contractile Dysfunction and Suppress Fibrosis in the MDX Mice
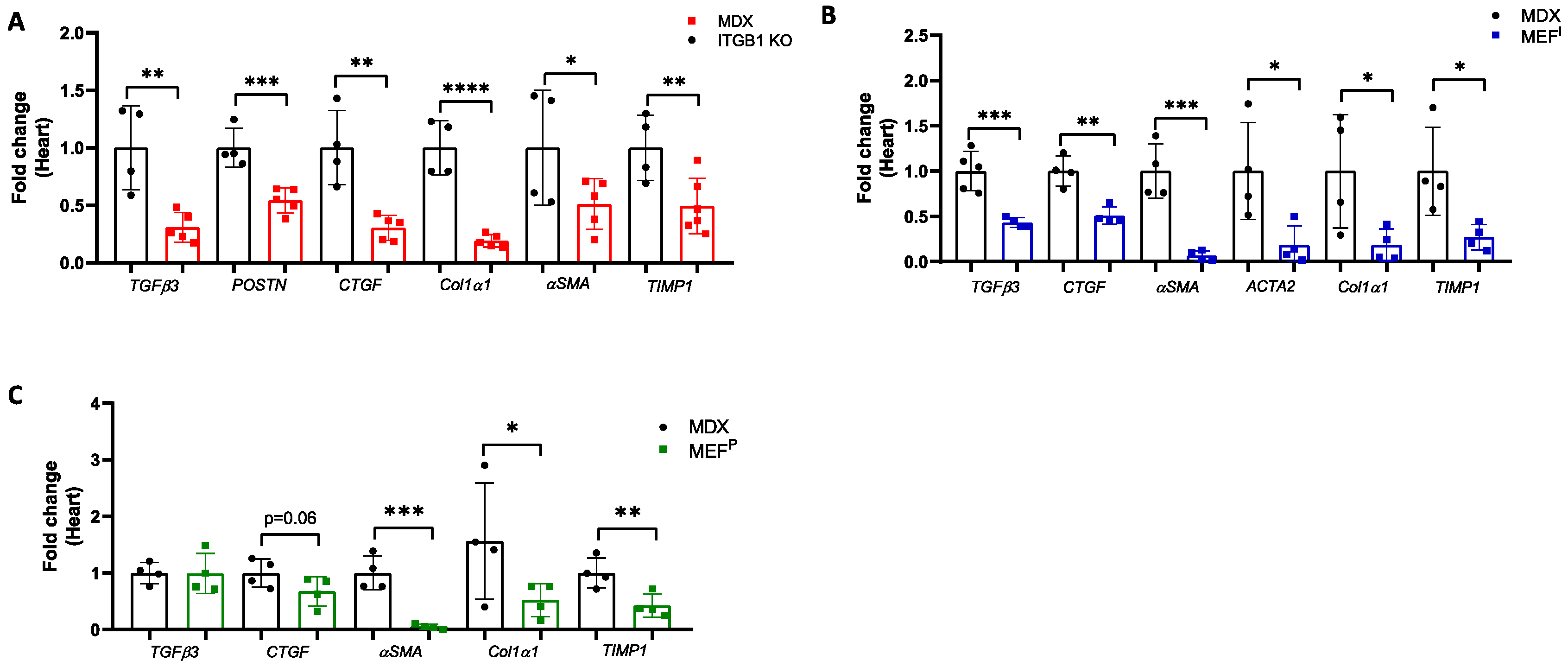
2.2. Improved Contractile Function of the Heart Is Accompanied by a Reduction in Fibrosis Hallmark GENE Markers
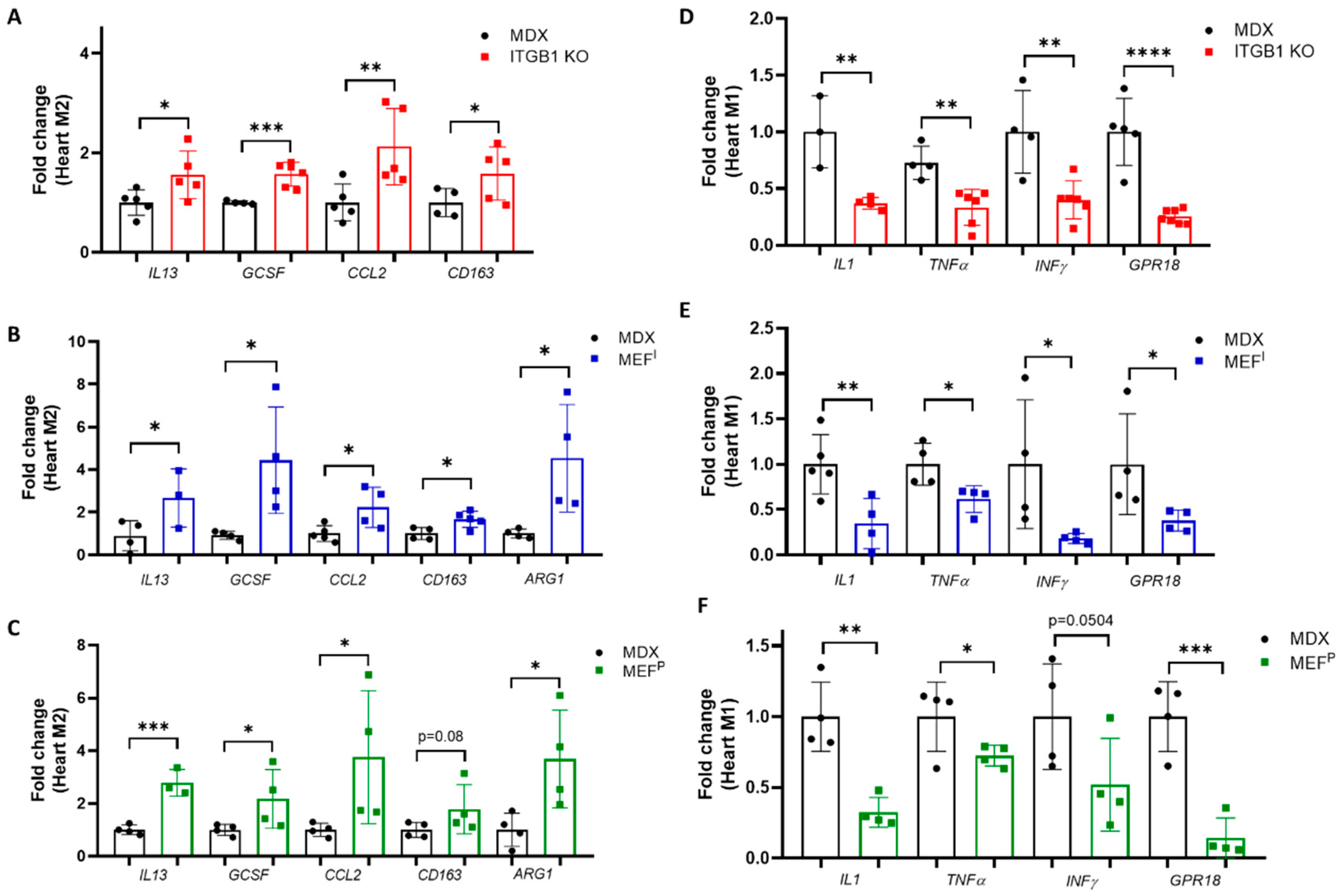
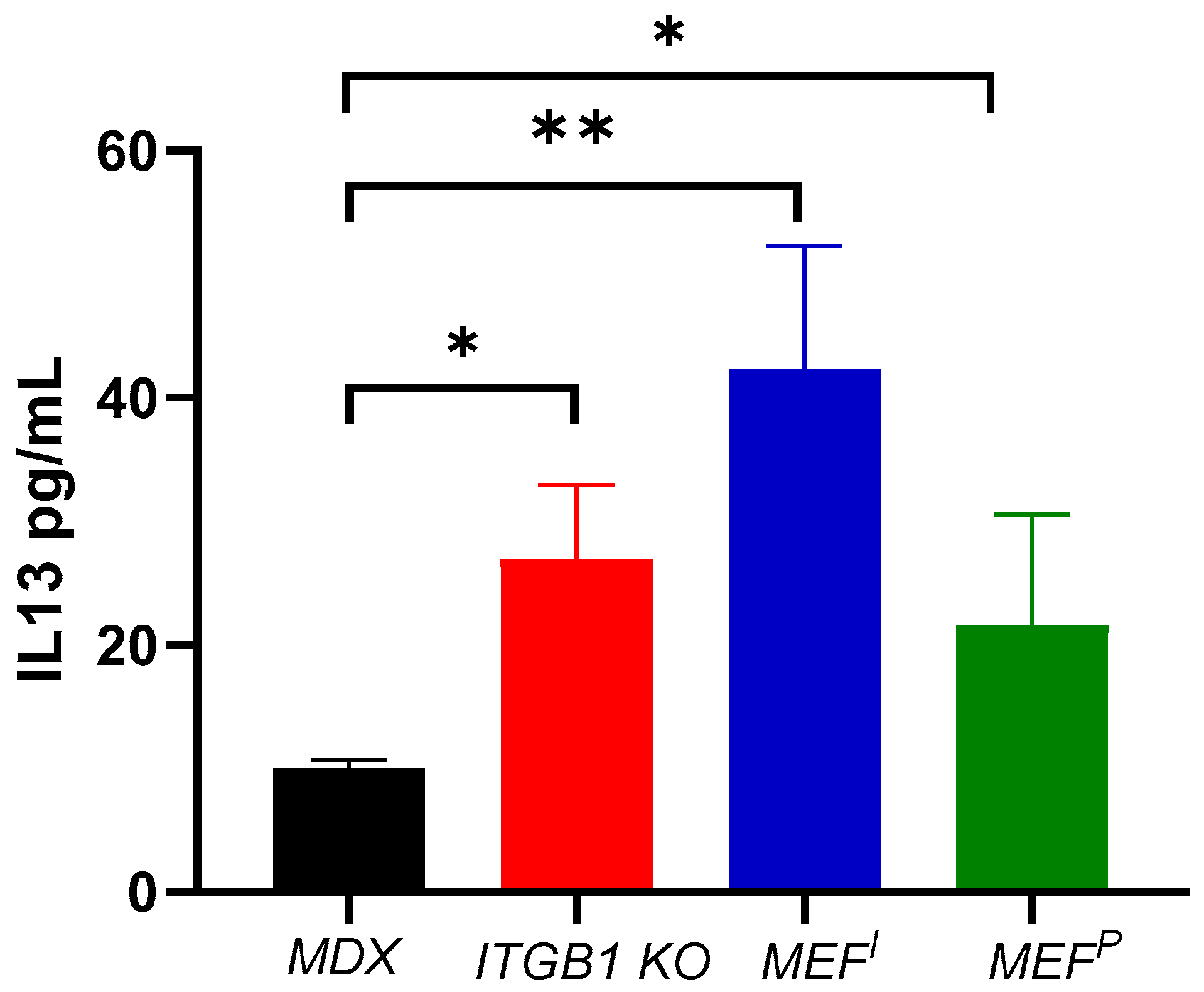
2.3. M2 Macrophage Gene Expression in the Heart
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Lines
4.3. Cell Implantation
4.4. Echocardiography
4.5. RNA Extraction
4.6. Quantitative Real-Time PCR
4.7. Fibrosis Staining
4.8. ELISA
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murtha, L.A.; Schuliga, M.J.; Mabotuwana, N.S.; Hardy, S.A.; Waters, D.W.; Burgess, J.K.; Knight, D.A.; Boyle, A.J. The processes and mechanisms of cardiac and pulmonary fibrosis. Front. Physiol. 2017, 8, 777. [Google Scholar] [CrossRef] [PubMed]
- Awwad, L.; Shofti, R.; Haas, T.; Aronheim, A. Tumor growth ameliorates cardiac dysfunction. Cells 2023, 12, 1853. [Google Scholar] [CrossRef] [PubMed]
- Awwad, L.; Aronheim, A. Tumor Progression Reverses Cardiac Hypertrophy and Fibrosis in a Tetracycline-Regulated ATF3 Transgenic Mouse Model. Cells 2023, 12, 2289. [Google Scholar] [CrossRef] [PubMed]
- Giovarelli, M.; Arnaboldi, F.; Zecchini, S.; Cornaghi, L.B.; Nava, A.; Sommariva, M.; Clementi, E.G.I.; Gagliano, N. Characterisation of progressive skeletal muscle fibrosis in the Mdx mouse model of duchenne muscular dystrophy: An in vivo and in vitro study. Int. J. Mol. Sci. 2022, 23, 8735. [Google Scholar] [CrossRef]
- Achlaug, L.; Awwad, L.; Langier Goncalves, I.; Goldenberg, T.; Aronheim, A. Tumor Growth Ameliorates Cardiac Dysfunction and Suppresses Fibrosis in a Mouse Model for Duchenne Muscular Dystrophy. Int. J. Mol. Sci. 2023, 24, 12595. [Google Scholar] [CrossRef]
- Wolford, C.C.; McConoughey, S.J.; Jalgaonkar, S.P.; Leon, M.; Merchant, A.S.; Dominick, J.L.; Yin, X.; Chang, Y.; Zmuda, E.J.; O’Toole, S.A.; et al. Transcription factor ATF3 links host adaptive response to breast cancer metastasis. J. Clin. Investig. 2013, 123, 2893–2906. [Google Scholar] [CrossRef] [PubMed]
- Bertram, J.S.; Janik, P. Establishment of a cloned line of Lewis Lung Carcinoma cells adapted to cell culture. Cancer Lett. 1980, 11, 63–73. [Google Scholar] [CrossRef]
- Humphries, J.D.; Byron, A.; Humphries, M.J. Integrin ligands at a glance. J. Cell Sci. 2006, 119 Pt 19, 3901–3903. [Google Scholar] [CrossRef]
- Kreidberg, J.A.; Symons, J.M. Integrins in kidney development, function, and disease. Am. J. Physiol.-Ren. Physiol. 2000, 279, F233–F242. [Google Scholar] [CrossRef]
- Hamidi, H.; Ivaska, J. Every step of the way: Integrins in cancer progression and metastasis. Nat. Rev. Cancer 2018, 18, 533–548. [Google Scholar] [CrossRef]
- Hou, S.; Isaji, T.; Hang, Q.; Im, S.; Fukuda, T.; Gu, J. Distinct effects of β1 integrin on cell proliferation and cellular signaling in MDA-MB-231 breast cancer cells. Sci. Rep. 2016, 6, 18430. [Google Scholar] [CrossRef] [PubMed]
- Langier Goncalves, I.; Awwad, L.; Aviram, S.; Izraeli, T.; Achlaug, L.; Aronheim, A. Heart Failure Promotes Cancer Progression in an Integrin β1-Dependent Manner. Int. J. Mol. Sci. 2023, 24, 17367. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Okita, K.; Nakagawa, M.; Yamanaka, S. Induction of pluripotent stem cells from fibroblast cultures. Nat. Protoc. 2007, 2, 3081–3089. [Google Scholar] [CrossRef] [PubMed]
- Choupani, F.; Assadollahi, V.; Vahabzadeh, Z.; Daneshi, E.; Abouzaripour, M.; Soleimani, F.; Bahrami, S.; Fathi, F. Feeding role of mouse embryonic fibroblast cells is influenced by genetic background, cell passage and day of isolation. Zygote 2022, 30, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Brocke, J.v.; Schmeiser, H.H.; Reinbold, M.; Hollstein, M. MEF immortalization to investigate the ins and outs of mutagenesis. Carcinogenesis 2006, 27, 2141–2147. [Google Scholar] [CrossRef][Green Version]
- Kim, Y.; Nurakhayev, S.; Nurkesh, A.; Zharkinbekov, Z.; Saparov, A. Macrophage polarization in cardiac tissue repair following myocardial infarction. Int. J. Mol. Sci. 2021, 22, 2715. [Google Scholar] [CrossRef] [PubMed]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage polarization: Different gene signatures in M1 (LPS+) vs. classically and M2 (LPS–) vs. alternatively activated macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Chen, Y.; Gao, C. Interleukin-13 reduces cardiac injury and prevents heart dysfunction in viral myocarditis via enhanced M2 macrophage polarization. Oncotarget 2017, 8, 99495. [Google Scholar] [CrossRef] [PubMed]
- Hinderer, S.; Schenke-Layland, K. Cardiac fibrosis–A short review of causes and therapeutic strategies. Adv. Drug Deliv. Rev. 2019, 146, 77–82. [Google Scholar] [CrossRef]
- Duan, D.; Goemans, N.; Takeda, S.i.; Mercuri, E.; Aartsma-Rus, A. Duchenne muscular dystrophy. Nat. Rev. Dis. Primers 2021, 7, 13. [Google Scholar] [CrossRef]
- Verhaart, I.E.; Aartsma-Rus, A. Therapeutic developments for Duchenne muscular dystrophy. Nat. Rev. Neurol. 2019, 15, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.; Evangelista, M.; Simili, M.; Mariani, L.; Pitto, L.; Rainaldi, G. Immortalization of MEF is characterized by the deregulation of specific miRNAs with potential tumor suppressor activity. Aging (Albany NY) 2011, 3, 665. [Google Scholar] [CrossRef] [PubMed]
- Saeed, H.; Taipaleenmäki, H.; Aldahmash, A.M.; Abdallah, B.M.; Kassem, M. Mouse embryonic fibroblasts (MEF) exhibit a similar but not identical phenotype to bone marrow stromal stem cells (BMSC). Stem Cell Rev. Rep. 2012, 8, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, D.; Gutiérrez, J.; Cabello-Verrugio, C.; Morales, M.G.; Mezzano, S.; Fadic, R.; Casar, J.C.; Hancke, J.L.; Brandan, E. Andrographolide attenuates skeletal muscle dystrophy in mdx mice and increases efficiency of cell therapy by reducing fibrosis. Skelet. Muscle 2014, 4, 6. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.G.; Gutierrez, J.; Cabello-Verrugio, C.; Cabrera, D.; Lipson, K.E.; Goldschmeding, R.; Brandan, E. Reducing CTGF/CCN2 slows down mdx muscle dystrophy and improves cell therapy. Hum. Mol. Genet. 2013, 22, 4938–4951. [Google Scholar] [CrossRef] [PubMed]
- Tabebordbar, M.; Zhu, K.; Cheng, J.K.; Chew, W.L.; Widrick, J.J.; Yan, W.X.; Maesner, C.; Wu, E.Y.; Xiao, R.; Ran, F.A. In vivo gene editing in dystrophic mouse muscle and muscle stem cells. Science 2016, 351, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Dubinin, M.V.; Mikheeva, I.B.; Stepanova, A.E.; Igoshkina, A.D.; Cherepanova, A.A.; Semenova, A.A.; Sharapov, V.A.; Kireev, I.I.; Belosludtsev, K.N. Mitochondrial Transplantation Therapy Ameliorates Muscular Dystrophy in mdx Mouse Model. Biomolecules 2024, 14, 316. [Google Scholar] [CrossRef] [PubMed]
- Kourakis, S.; Timpani, C.A.; de Haan, J.B.; Gueven, N.; Fischer, D.; Rybalka, E. Targeting Nrf2 for the treatment of Duchenne muscular dystrophy. Redox Biol. 2021, 38, 101803. [Google Scholar] [CrossRef]
- Wynn, T.A.; Vannella, K.M. Macrophages in tissue repair, regeneration, and fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef]
- Morimoto, H.; Takahashi, M.; Izawa, A.; Ise, H.; Hongo, M.; Kolattukudy, P.E.; Ikeda, U. Cardiac overexpression of monocyte chemoattractant protein-1 in transgenic mice prevents cardiac dysfunction and remodeling after myocardial infarction. Circ. Res. 2006, 99, 891–899. [Google Scholar] [CrossRef]
- Ley, K. M1 means kill; M2 means heal. J. Immunol. 2017, 199, 2191–2193. [Google Scholar] [CrossRef] [PubMed]
- Honkisz-Orzechowska, E.; Łażewska, D.; Baran, G.; Kieć-Kononowicz, K. Uncovering the Power of GPR18 Signalling: How RvD2 and Other Ligands Could Have the Potential to Modulate and Resolve Inflammation in Various Health Disorders. Molecules 2024, 29, 1258. [Google Scholar] [CrossRef] [PubMed]
- Juhas, U.; Ryba-Stanisławowska, M.; Szargiej, P.; Myśliwska, J. Different pathways of macrophage activation and polarization. Adv. Hyg. Exp. Med. 2015, 69, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Xu, J. Preparation, culture, and immortalization of mouse embryonic fibroblasts. Curr. Protoc. Mol. Biol. 2005, 70, 28.1.1–28.1.8. [Google Scholar] [CrossRef] [PubMed]
- Darlyuk-Saadon, I.; Weidenfeld-Baranboim, K.; Yokoyama, K.K.; Hai, T.; Aronheim, A. The bZIP repressor proteins, c-Jun dimerization protein 2 and activating transcription factor 3, recruit multiple HDAC members to the ATF3 promoter. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2012, 1819, 1142–1153. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Achlaug, L.; Langier Goncalves, I.; Aronheim, A. Cardioprotection and Suppression of Fibrosis by Diverse Cancer and Non-Cancer Cell Lines in a Murine Model of Duchenne Muscular Dystrophy. Int. J. Mol. Sci. 2024, 25, 4273. https://doi.org/10.3390/ijms25084273
Achlaug L, Langier Goncalves I, Aronheim A. Cardioprotection and Suppression of Fibrosis by Diverse Cancer and Non-Cancer Cell Lines in a Murine Model of Duchenne Muscular Dystrophy. International Journal of Molecular Sciences. 2024; 25(8):4273. https://doi.org/10.3390/ijms25084273
Chicago/Turabian StyleAchlaug, Laris, Irina Langier Goncalves, and Ami Aronheim. 2024. "Cardioprotection and Suppression of Fibrosis by Diverse Cancer and Non-Cancer Cell Lines in a Murine Model of Duchenne Muscular Dystrophy" International Journal of Molecular Sciences 25, no. 8: 4273. https://doi.org/10.3390/ijms25084273
APA StyleAchlaug, L., Langier Goncalves, I., & Aronheim, A. (2024). Cardioprotection and Suppression of Fibrosis by Diverse Cancer and Non-Cancer Cell Lines in a Murine Model of Duchenne Muscular Dystrophy. International Journal of Molecular Sciences, 25(8), 4273. https://doi.org/10.3390/ijms25084273