
Therapeutic Potential of 4-Hexylresorcinol in Preserving Testicular Function in Streptozotocin-Induced Diabetic Rats





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
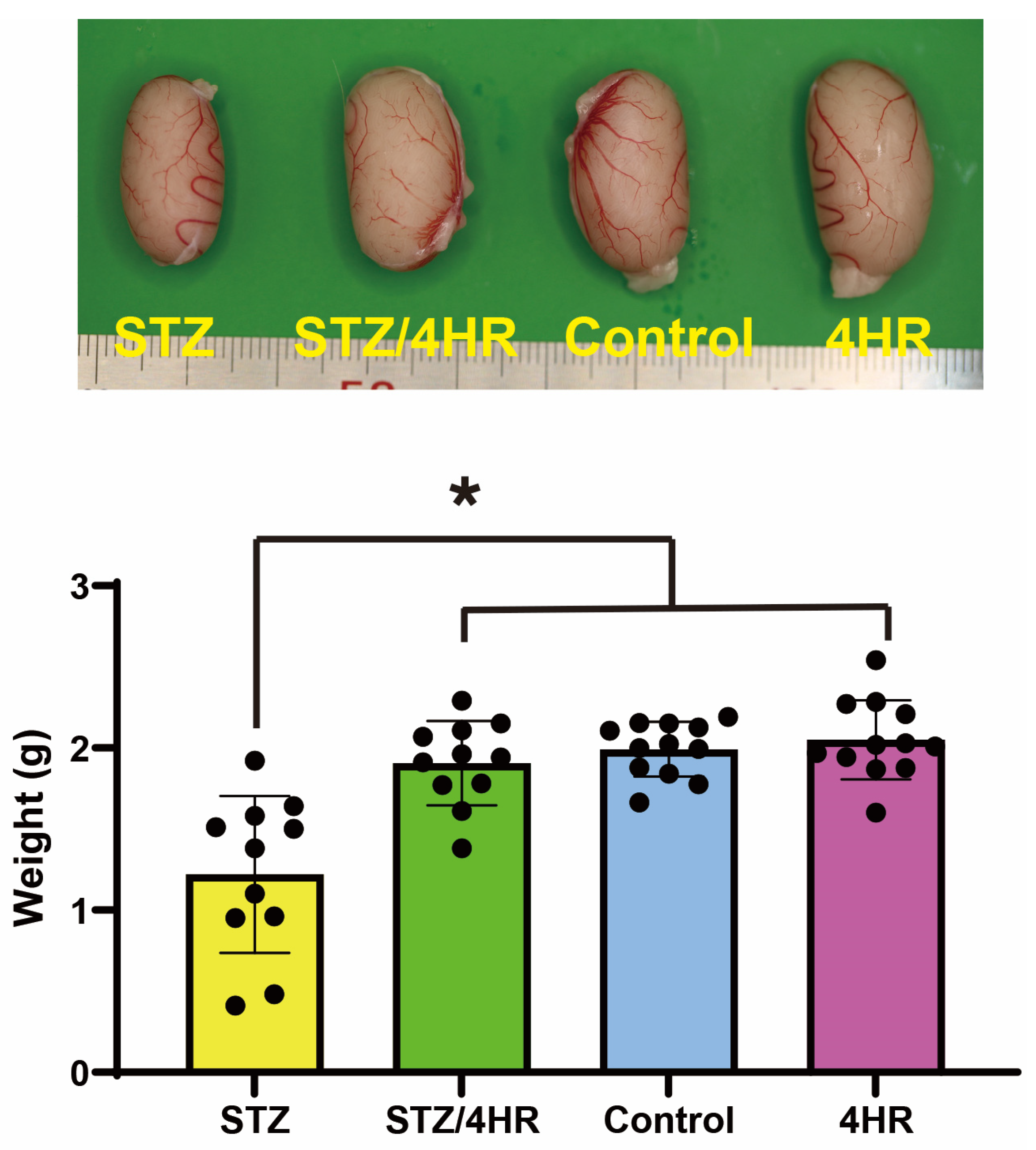
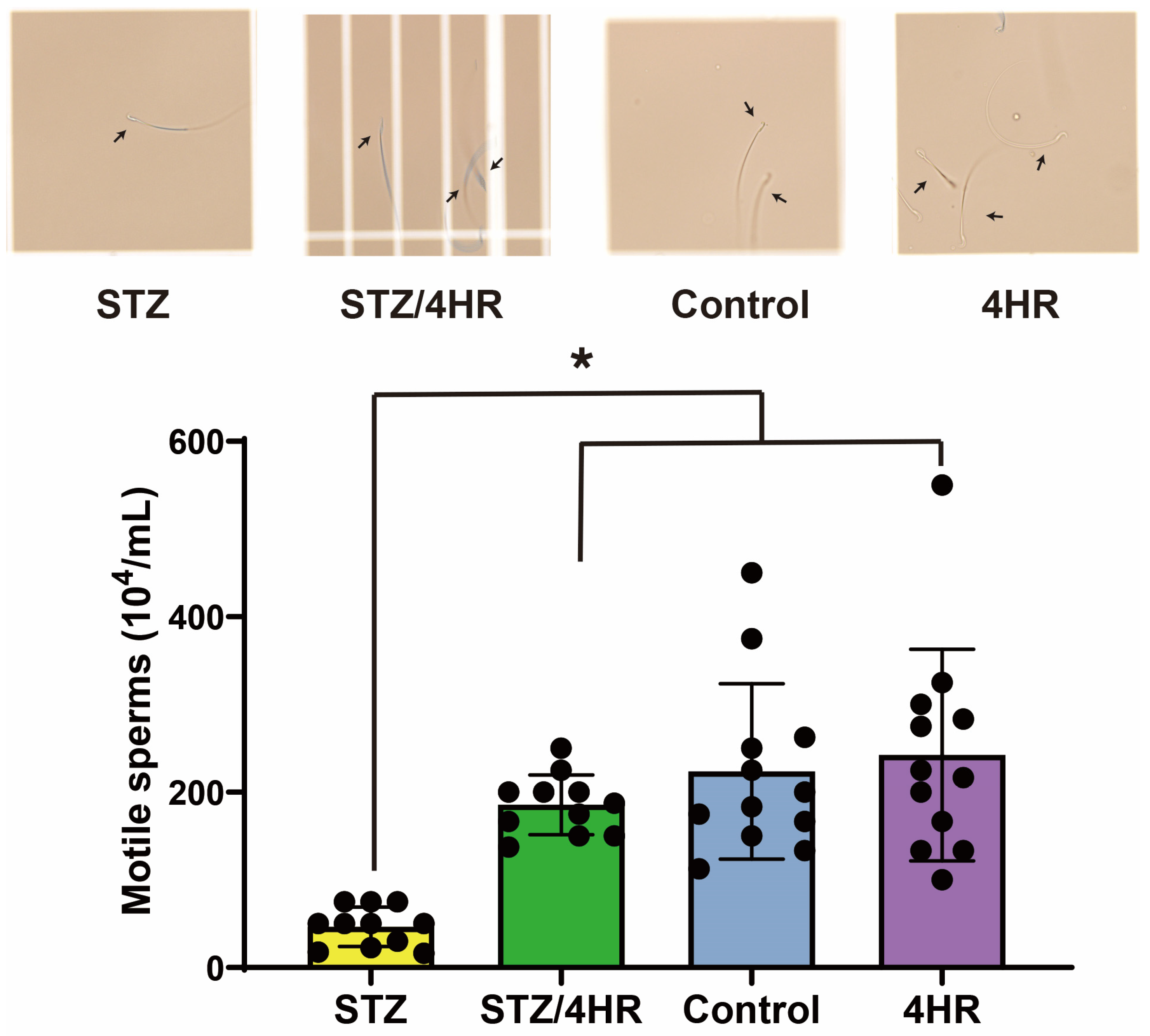
2.1. Effect of STZ and 4HR on Testis Weight and Sperm Motility in Diabetic and Non-Diabetic Rats
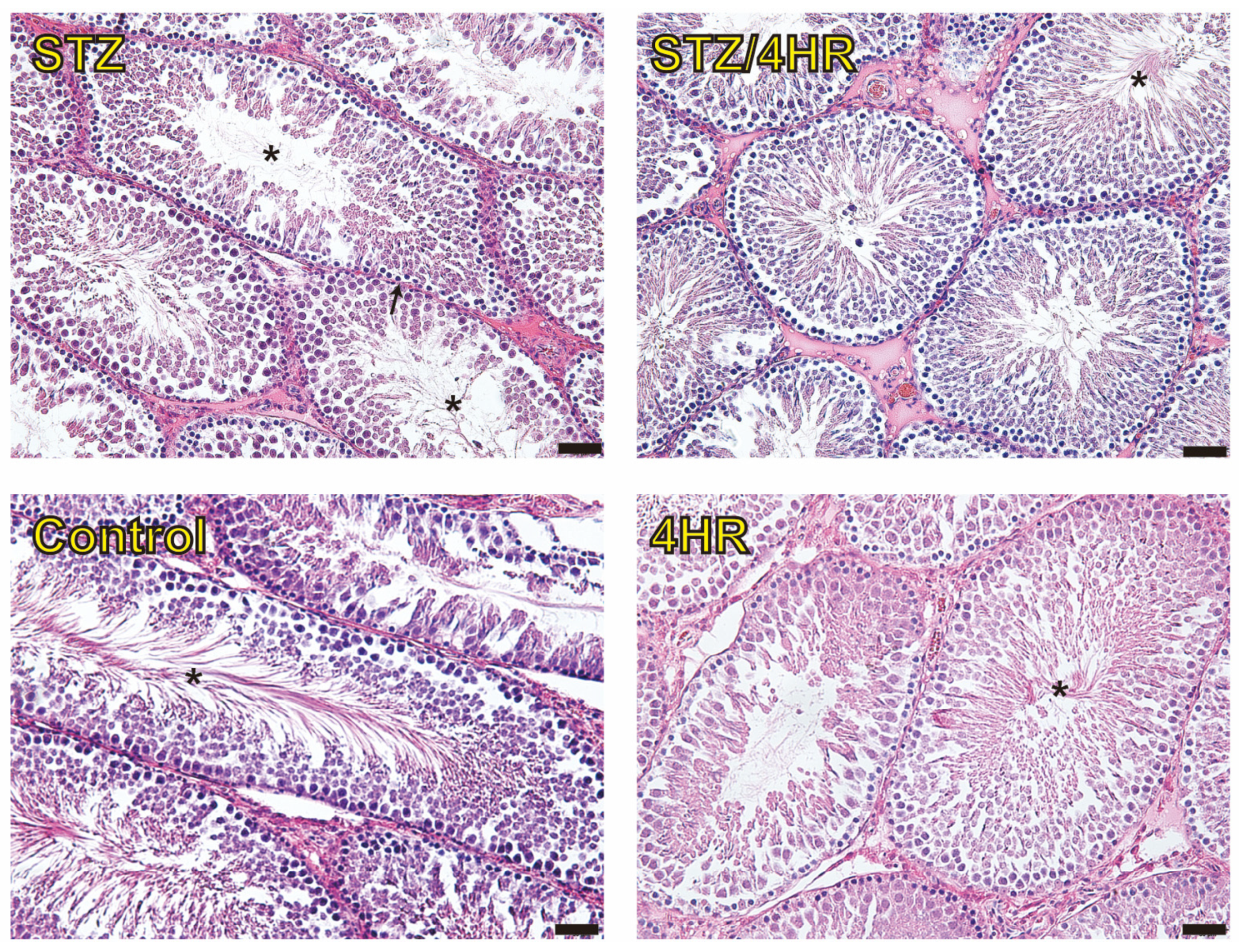
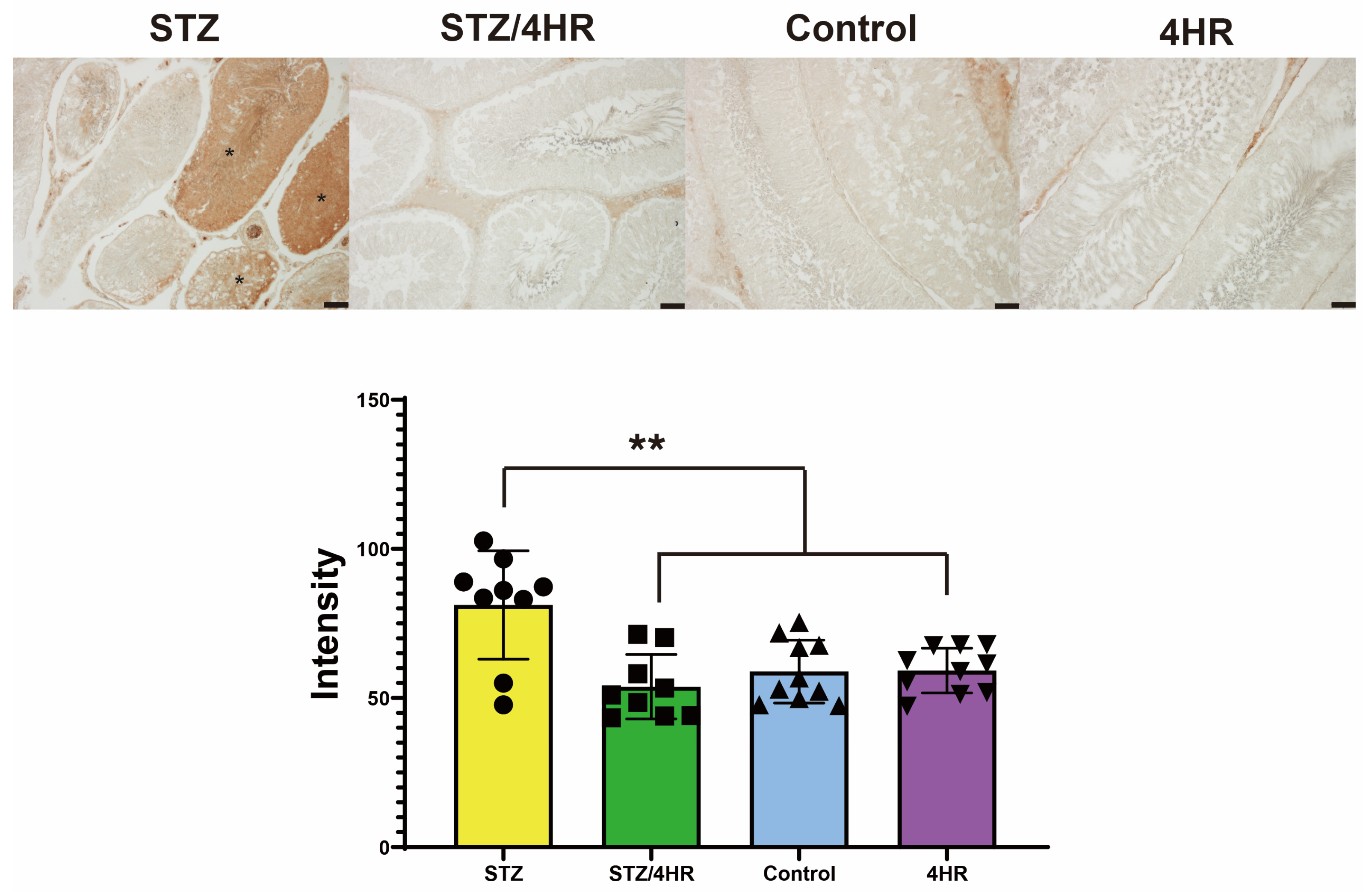
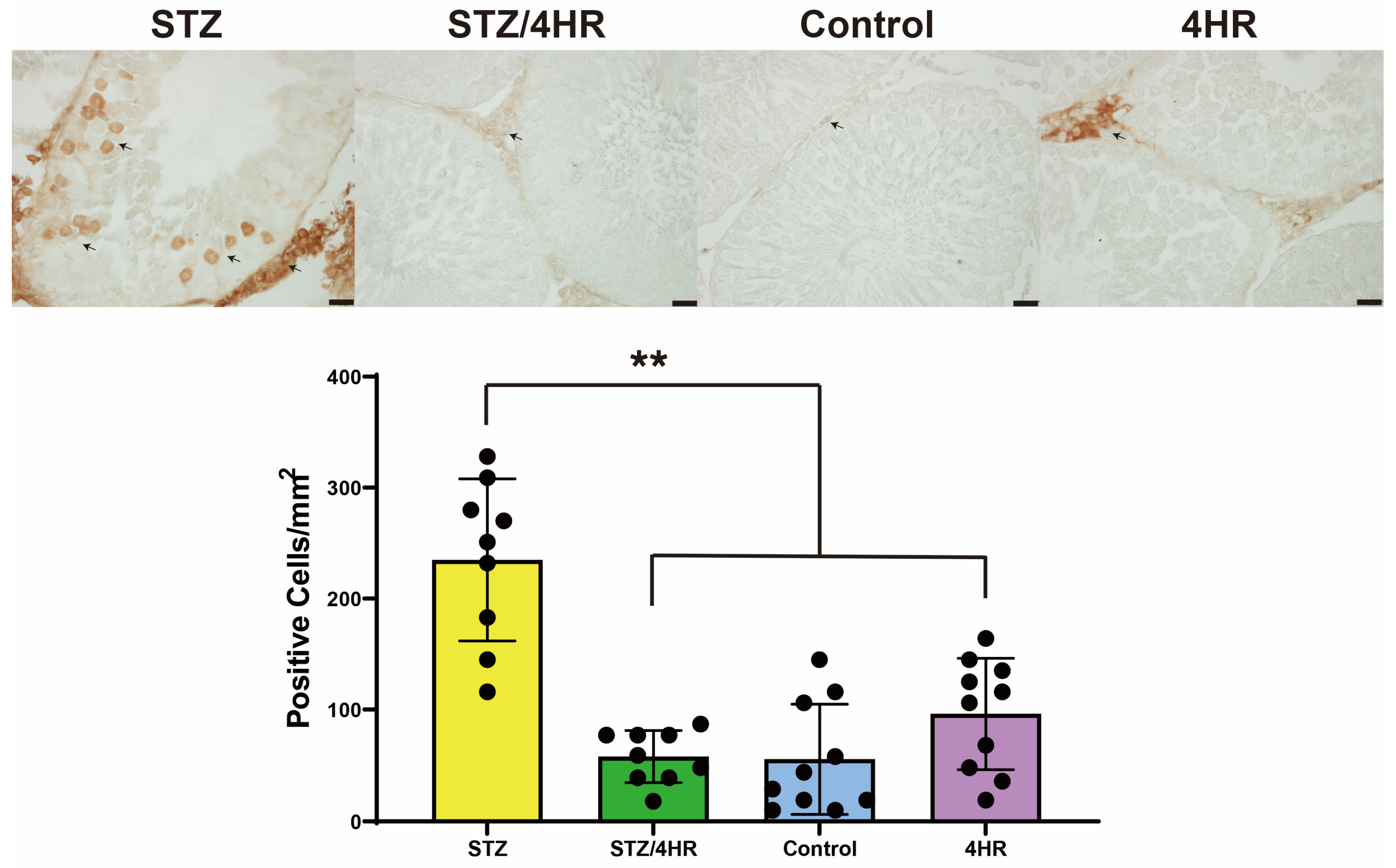
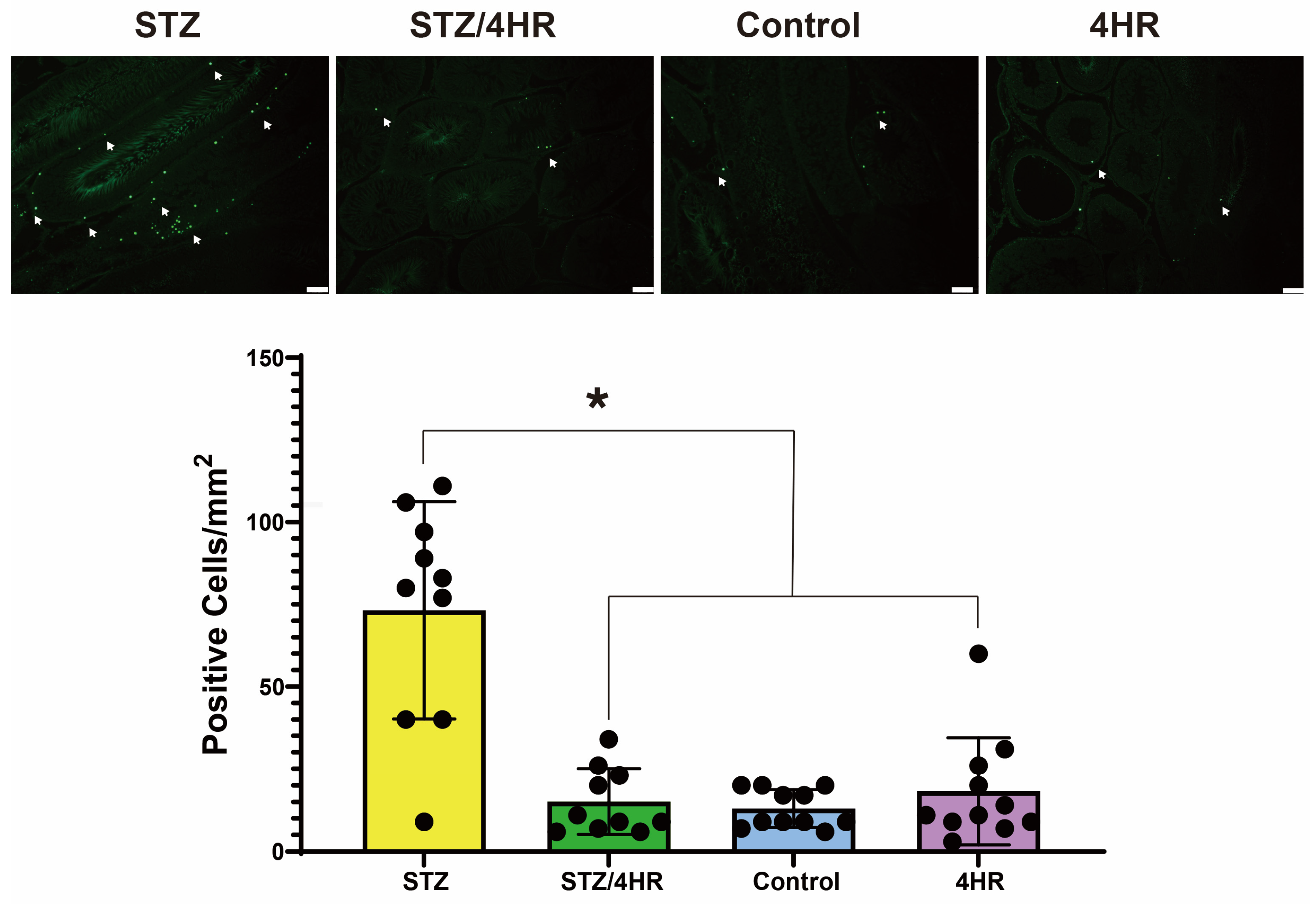
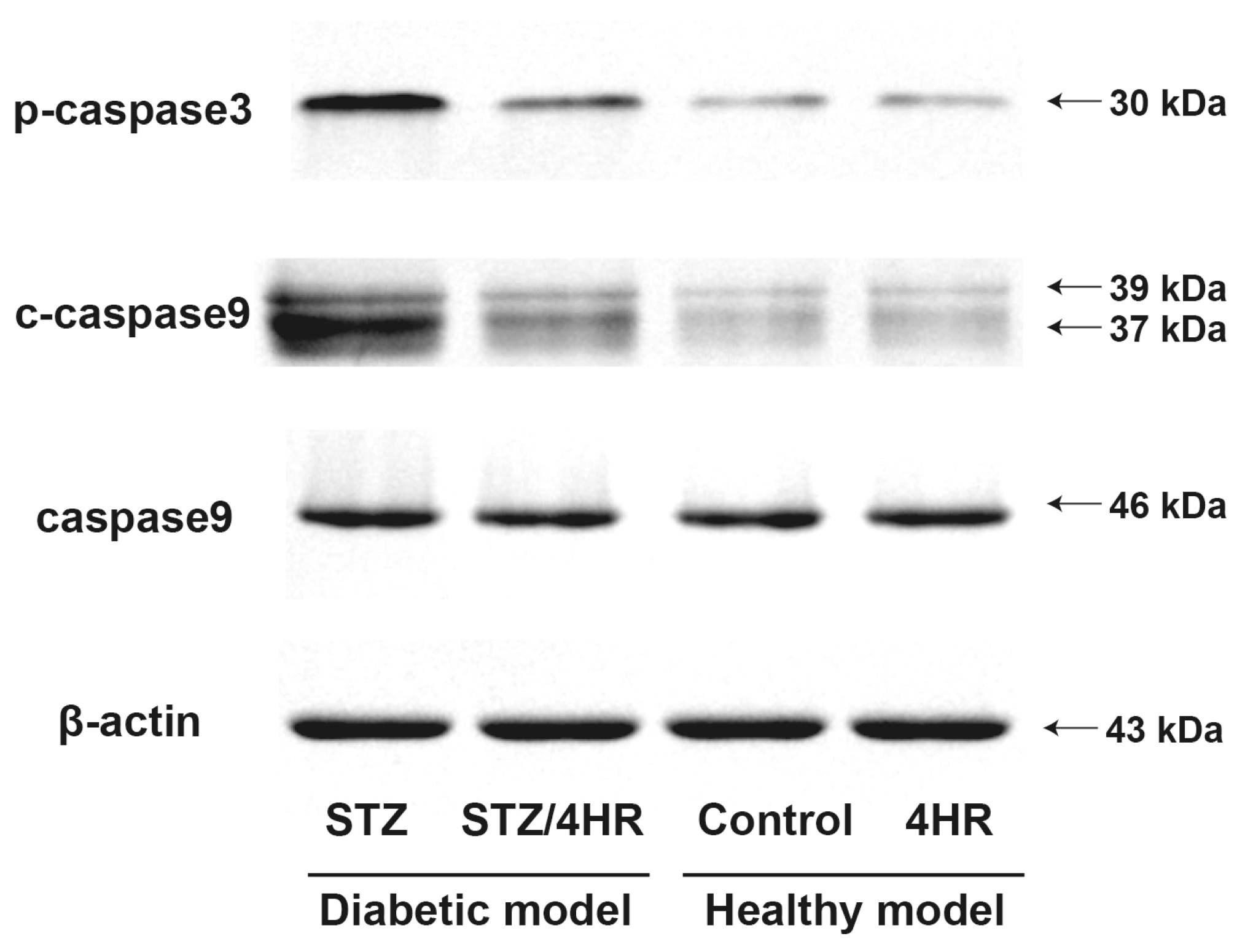
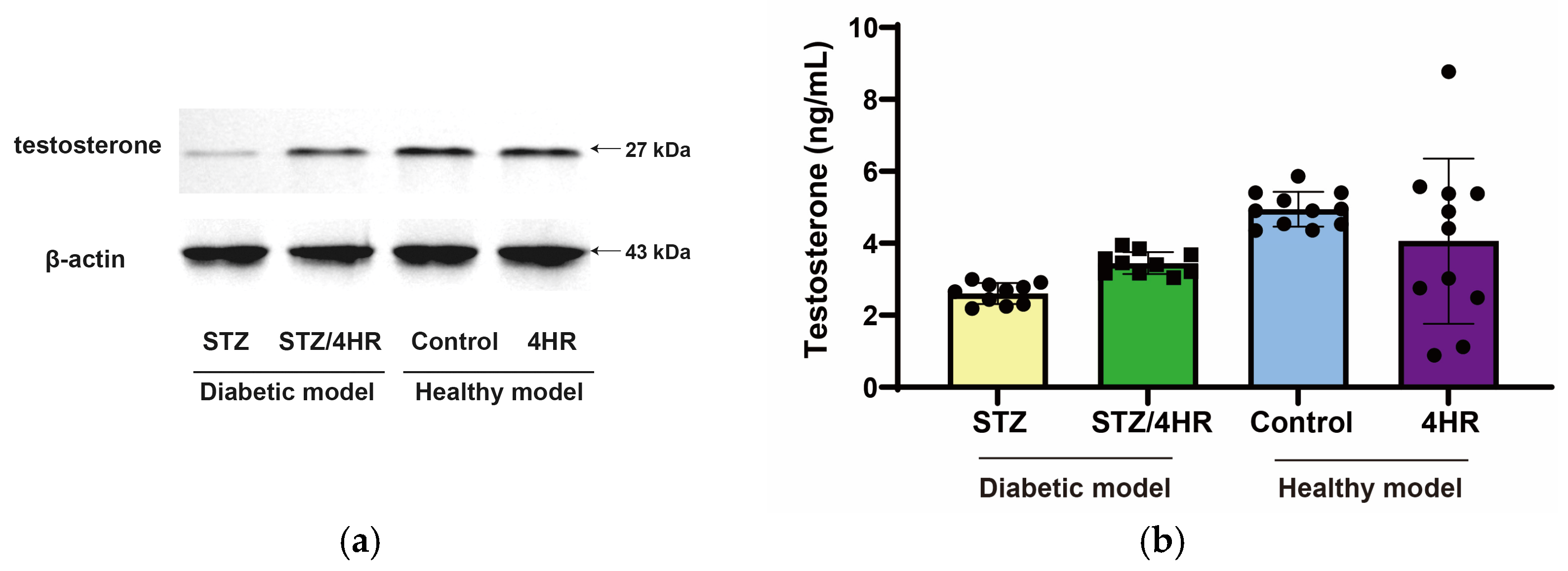
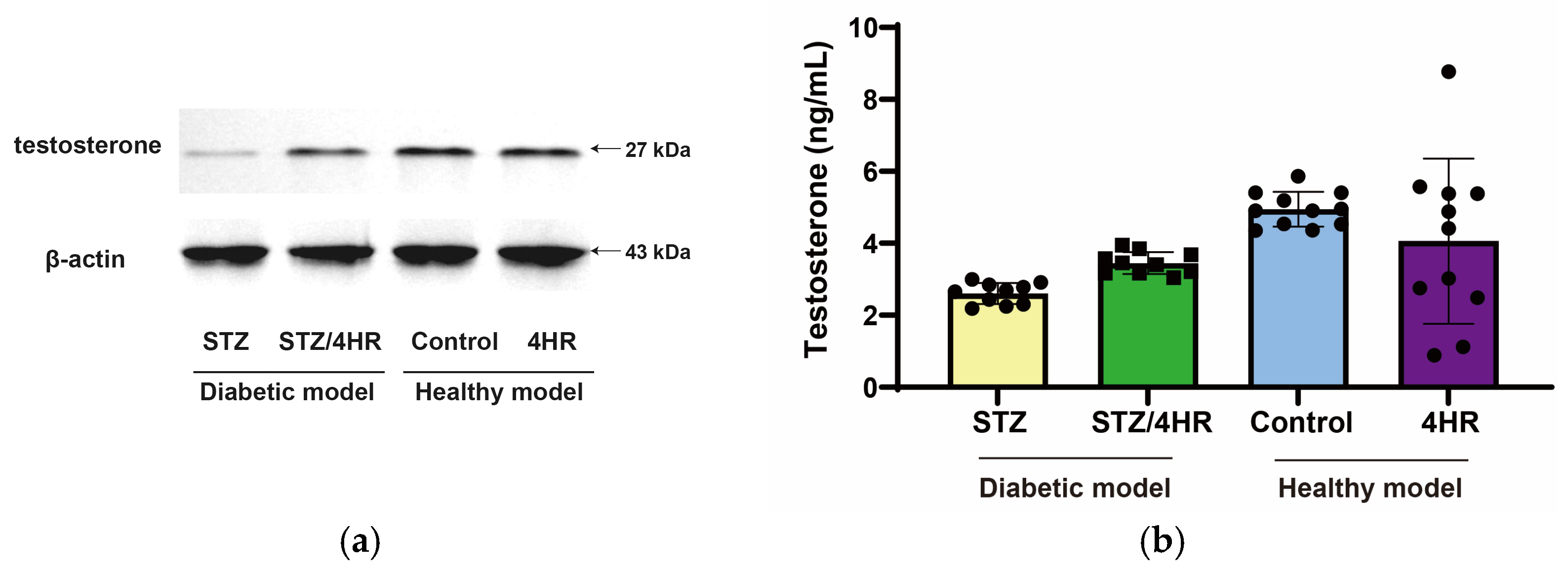
2.2. Effects of 4HR on Sperm Morphology, Apoptosis, and Serum Testosterone Levels in Diabetic Rats
3. Discussion
4. Materials and Methods
4.1. Animal Study
4.2. Sperm Motility Assay
4.3. Histological Staining and Immunohistochemistry
4.4. TUNEL Assay
4.5. Western Blot
4.6. Measuring Serum Testosterone Level
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barkabi-Zanjani, S.; Ghorbanzadeh, V.; Aslani, M.; Ghalibafsabbaghi, A.; Chodari, L. Diabetes mellitus and the impairment of male reproductive function: Possible signaling pathways. Diabetes Metab. Syndr. 2020, 14, 1307–1314. [Google Scholar] [CrossRef] [PubMed]
- Panghal, A.; Jena, G. Gut-gonad perturbations in type-1 diabetes mellitus: Role of dysbiosis, oxidative stress, inflammation and energy-dysbalance. Curr. Diabetes Rev. 2023; Epub ahead of print. [Google Scholar] [CrossRef]
- Desai, A.; Chen, R.; Cayetano, A.; Jayasena, C.N.; Minhas, S. Understanding and treating ejaculatory dysfunction in men with diabetes mellitus. Andrology 2023, 11, 379–398. [Google Scholar] [CrossRef] [PubMed]
- Malavige, L.S.; Levy, J.C. Erectile dysfunction in diabetes mellitus. J. Sex. Med. 2009, 6, 1232–1247. [Google Scholar] [CrossRef] [PubMed]
- Lotti, F.; Maggi, M. Effects of diabetes mellitus on sperm quality and fertility outcomes: Clinical evidence. Andrology 2023, 11, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Baccetti, B.; La Marca, A.; Piomboni, P.; Capitani, S.; Bruni, E.; Petraglia, F.; De Leo, V. Insulin-dependent diabetes in men is associated with hypothalamo-pituitary derangement and with impairment in semen quality. Hum. Reprod. 2002, 17, 2673–26777. [Google Scholar] [CrossRef] [PubMed]
- Adashi, E.Y.; Hsueh, A.J.; Yen, S.S. Insulin enhancement of luteinizing hormone and follicle-stimulating hormone release by cultured pituitary cells. Endocrinology 1981, 108, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Garris, D.R.; Williams, S.K.; Coleman, D.L.; Morgan, C.R. Glucose utilization by the mouse brain: Influence of age and diabetes. Brain Res. 1984, 317, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Seethalakshmi, L.; Menon, M.; Diamond, D. The effect of streptozotocin-induced diabetes on the neuroendocrine-male reproductive tract axis of the adult rat. J. Urol. 1987, 138, 190–194. [Google Scholar] [CrossRef]
- Ghosh, S.; Chowdhury, S.; Das, A.K.; Sil, P.C. Taurine ameliorates oxidative stress induced inflammation and ER stress mediated testicular damage in STZ-induced diabetic Wistar rats. Food Chem. Toxicol. 2019, 124, 64–80. [Google Scholar] [CrossRef]
- Heeba, G.H.; Hamza, A.A. Rosuvastatin ameliorates diabetes-induced reproductive damage via suppression of oxidative stress, inflammatory and apoptotic pathways in male rats. Life Sci. 2015, 141, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Tabak, O.; Gelisgen, R.; Erman, H.; Erdenen, F.; Muderrisoglu, C.; Aral, H.; Uzun, H. Oxidative lipid, protein, and DNA damage as oxidative stress markers in vascular complications of diabetes mellitus. Clin. Investig. Med. 2011, 34, E163–E171. [Google Scholar] [CrossRef]
- Palmeira, C.M.; Santos, D.L.; Seiça, R.; Moreno, A.J.; Santos, M.S. Enhanced mitochondrial testicular antioxidant capacity in Goto-Kakizaki diabetic rats: Role of coenzyme Q. Am. J. Physiol. Cell Physiol. 2001, 281, C1023–C1028. [Google Scholar] [CrossRef]
- Khosravi, Z.; Sedaghat, R.; Baluchnejadmojarad, T.; Roghani, M. Diosgenin ameliorates testicular damage in streptozotocin-diabetic rats through attenuation of apoptosis, oxidative stress, and inflammation. Int. Immunopharmacol. 2019, 70, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Tsounapi, P.; Honda, M.; Dimitriadis, F.; Kawamoto, B.; Hikita, K.; Muraoka, K.; Saito, M.; Sofikitis, N.; Takenaka, A. Impact of antioxidants on seminal vesicles function and fertilizing potential in diabetic rats. Asian J. Androl. 2017, 19, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Nna, V.U.; Bakar, A.B.A.; Ahmad, A.; Mohamed, M. Diabetes-induced testicular oxidative stress, inflammation, and caspase-dependent apoptosis: The protective role of metformin. Arch. Physiol. Biochem. 2020, 126, 377–388. [Google Scholar] [CrossRef]
- He, W.; Liu, H.; Hu, L.; Wang, Y.; Huang, L.; Liang, A.; Wang, X.; Zhang, Q.; Chen, Y.; Cao, Y.; et al. Icariin improves testicular dysfunction via enhancing proliferation and inhibiting mitochondria-dependent apoptosis pathway in high-fat diet and streptozotocin-induced diabetic rats. Reprod. Biol. Endocrinol. 2021, 19, 168. [Google Scholar] [CrossRef]
- Chen, Y.; Jiao, N.; Jiang, M.; Liu, L.; Zhu, Y.; Wu, H.; Chen, J.; Fu, Y.; Du, Q.; Xu, H.; et al. Loganin alleviates testicular damage and germ cell apoptosis induced by AGEs upon diabetes mellitus by suppressing the RAGE/p38MAPK/NF-κB pathway. J. Cell Mol. Med. 2020, 24, 6083–6095. [Google Scholar] [CrossRef]
- Aziz, N.M.; Kamel, M.Y.; Mohamed, M.S.; Ahmed, S.M. Antioxidant, anti-inflammatory, and anti-apoptotic effects of zinc supplementation in testes of rats with experimentally induced diabetes. Appl. Physiol. Nutr. Metab. 2018, 43, 1010–1018. [Google Scholar] [CrossRef]
- Alsieni, M.A.; El Rabey, H.A.; Al-Sieni, A.I.; Al-Seeni, M.N. Comparison between the antioxidant and antidiabetic activity of fenugreek and buckthorn in streptozotocin-induced diabetic male rats. Biomed. Res. Int. 2021, 2021, 7202447. [Google Scholar] [CrossRef]
- Sharifi, M.; Ghadamyari, M.; Sajedi, R.H.; Mahmoodi, N.O. Effects of 4-hexylresorcinol on the phenoloxidase from Hyphantria cunea (Lepidoptera: Arctiidae): In vivo and in vitro studies. Insect Sci. 2015, 22, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Jo, Y.Y.; Garagiola, U.; Choi, J.Y.; Kang, Y.J.; Oh, J.H.; Kim, S.G. Increased level of vascular endothelial growth factors by 4-hexylresorcinol is mediated by transforming growth factor-beta1 and accelerates capillary regeneration in the burns in diabetic animals. Int. J. Mol. Sci. 2020, 21, 3473. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Ma, P.; Torshizi, R. The impact of some natural phenolic compounds on α-glucosidase and sorbitol dehydrogenase enzymes, and anti-leukemia cancer potential, spin density distributions, and in silico studies. J. Oleo Sci. 2022, 71, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.G. 4-Hexylresorcinol: Pharmacologic chaperone and its application for wound healing. Maxillofac. Plast. Reconstr. Surg. 2022, 44, 5. [Google Scholar] [CrossRef]
- Yi, W.; Liu, T.; Gao, X.; Xie, Y.; Liu, M. 4-Hexylresorcinol inhibits osteoclastogenesis by suppressing the NF-κB signaling pathway and reverses bone loss in ovariectomized mice. Exp. Ther. Med. 2021, 21, 354. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Liu, Q.; Chai, W.M.; Xia, S.S.; Yu, Z.Y.; Wei, Q.M. Inhibitory potential of 4-hexylresorcinol against alpha-glucosidase and non-enzymatic glycation: Activity and mechanism. J. Biosci. Bioeng. 2021, 131, 241–249. [Google Scholar] [CrossRef]
- Kizilay, G.; Ersoy, O.; Cerkezkayabekir, A.; Topcu-Tarladacalisir, Y. Sitagliptin and fucoidan prevent apoptosis and reducing ER stress in diabetic rat testes. Andrologia 2021, 53, e13858. [Google Scholar] [CrossRef] [PubMed]
- Chilukoti, S.R.; Sahu, C.; Jena, G. Protective role of eugenol against diabetes-induced oxidative stress, DNA damage, and apoptosis in rat testes. J. Biochem. Mol. Toxicol. 2024, 38, e23593. [Google Scholar] [CrossRef] [PubMed]
- Koroglu Aydın, P.; Karabulut-Bulan, O.; Bugan, I.; Turkyilmaz, I.B.; Altun, S.; Yanardag, R. The protective effect of metformin against testicular damage in diabetes and prostate cancer model. Cell Biochem. Funct. 2022, 40, 60–70. [Google Scholar] [CrossRef]
- Shokri, A.; Pourheydar, B.; Hossein Farjah, G.; Krimipour, M.; Pourheydar, M. Effects of glibenclamide and troxerutin on the sperm parameters and histopathological changes of testis in streptozotocin-induced diabetic male rats: An experimental study. Int. J. Reprod. Biomed. 2023, 21, 123–138. [Google Scholar] [CrossRef]
- Kucuk Baloglu, F.; Guldag Tas, D.; Yilmaz, O.; Severcan, F. The recovery effect of vitamin C on structural alterations due to Streptozotocin-Induced diabetes in rat testicular tissues. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2023, 288, 122149. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Qiao, X.; Cheng, L.; Liu, M.; Deng, Y.; Zhuo, X. LGR4 silence aggravates ischemic injury by modulating mitochondrial function and oxidative stress via ERK signaling pathway in H9c2 cells. J. Mol. Histol. 2021, 52, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Gaida, D.; Park, Y.W.; Kim, S.G. 4-Hexylresorcinol and its effects on circumvallate papillae taste buds in diabetic and healthy rats: An initial investigation. Appl. Sci. 2023, 13, 11617. [Google Scholar] [CrossRef]
- Elshafey, M.; Erfan, O.S.; Risha, E.; Badawy, A.M.; Ebrahim, H.A.; El-Sherbiny, M.; El-Shenbaby, I.; Enan, E.T.; Almadani, M.E.; Eldesoqui, M. Protective effect of stevia on diabetic induced testicular damage: An immunohistochemical and ultrastructural study. Eur. Rev. Med. Pharmacol. Sci. 2023, 27, 11039–11056. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Hu, Y.; Shao, M.; Chen, S.; Qi, S. Chromium picolinate protects against testicular damage in STZ-induced diabetic rats via anti-inflammation, anti-oxidation, inhibiting apoptosis, and regulating the TGF-β1/Smad pathway. Molecules 2023, 28, 7669. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.S.; Wu, C.Y.; Wang, Y.H.; Hu, Q.Q.; Sun, R.Y.; Pan, M.J.; Lu, X.Y.; Zhu, T.; Luo, S.; Yang, H.J.; et al. The protective effects of icariin against testicular dysfunction in type 1 diabetic mice Via AMPK-mediated Nrf2 activation and NF-κB p65 inhibition. Phytomedicine 2023, 123, 155217. [Google Scholar] [CrossRef] [PubMed]
- Nna, V.U.; Abu Bakar, A.B.; Ahmad, A.; Eleazu, C.O.; Mohamed, M. Oxidative stress, NF-κB-mediated inflammation and apoptosis in the testes of streptozotocin-induced diabetic rats: Combined protective effects of Malaysian propolis and metformin. Antioxidants 2019, 8, 465. [Google Scholar] [CrossRef] [PubMed]
- Minas, A.; Talebi, H.; Taravat Ray, M.; Yari Eisalou, M.; Alves, M.G.; Razi, M. Insulin treatment to type 1 male diabetic rats protects fertility by avoiding testicular apoptosis and cell cycle arrest. Gene 2021, 799, 145847. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Bai, X.; Le, L.; Tian, D.; Gao, H.; Qi, L.X.; Hu, K.P. Eucommia ulmoides Oliv. leaf extract improves erectile dysfunction in streptozotocin-induced diabetic rats by protecting endothelial function and ameliorating hypothalamic-pituitary-gonadal axis function. Evid. Based Complement. Altern. Med. 2019, 2019, 1782953. [Google Scholar] [CrossRef]
- Dhindsa, S.; Prabhakar, S.; Sethi, M.; Bandyopadhyay, A.; Chaudhuri, A.; Dandona, P. Frequent occurrence of hypogonadotropic hypogonadism in type 2 diabetes. J. Clin. Endocrinol. Metab. 2004, 89, 5462–5468. [Google Scholar] [CrossRef]
- Aly, H.A.A. Mitochondria-Mediated Apoptosis Induced Testicular Dysfunction in Diabetic Rats: Ameliorative Effect of Resveratrol. Endocrinology 2021, 162, bqab018. [Google Scholar] [CrossRef] [PubMed]
- Tanizawa, H.; Terada, M.; Watanabe, Y. Experimental studies on anthelmintics (XXVI). Biochemical and pharmacological studies of 4-iodothymol on Ascaris lumbricoides suum. Nihon Yakurigaku Zasshi 1976, 72, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Amadasi, A.; Mozzarelli, A.; Meda, C.; Maggi, A.; Cozzini, P. Identification of xenoestrogens in food additives by an integrated in silico and in vitro approach. Chem. Res. Toxicol. 2009, 22, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Omolaoye, T.S.; Skosana, B.T.; du Plessis, S.S. Diabetes mellitus- induction: Effect of different streptozotocin doses on male reproductive parameters. Acta Histochem. 2018, 120, 103–109. [Google Scholar] [CrossRef]
- Koh, P.O. Streptozotocin-induced diabetes increases apoptosis through JNK phosphorylation and Bax activation in rat testes. J. Vet. Med. Sci. 2007, 69, 969–971. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.H.; Kim, D.W.; Lee, S.K.; Kim, S.G.; Kim, T.W. The Administration of 4-Hexylresorcinol Accelerates Orthodontic Tooth Movement and Increases the Expression Level of Bone Turnover Markers in Ovariectomized Rats. Int. J. Mol. Sci. 2020, 21, 1526. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.S.; Kim, D.W.; Oh, J.H.; Lee, S.K.; Choi, J.Y.; Kim, S.G.; Kim, T.W. Effects of 4-Hexylresorcinol on Craniofacial Growth in Rats. Int. J. Mol. Sci. 2021, 22, 8935. [Google Scholar] [CrossRef]
- Omolaoye, T.; Windvogel, S.; du Plessis, S. Testicular oxidative stress and apoptosis status in streptozotocin-induced diabetic rats after treatment with rooibos (Aspalathus linearis), honeybush (Cyclopia intermedia), and sutherlandia (Lessertia frutescens) infusions. Asian Pac. J. Reprod. 2021, 10, 11–20. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, J.-H.; Choi, J.-Y.; Kim, D.-W.; Kim, S.-G.; Garagiola, U. Therapeutic Potential of 4-Hexylresorcinol in Preserving Testicular Function in Streptozotocin-Induced Diabetic Rats. Int. J. Mol. Sci. 2024, 25, 4316. https://doi.org/10.3390/ijms25084316
Oh J-H, Choi J-Y, Kim D-W, Kim S-G, Garagiola U. Therapeutic Potential of 4-Hexylresorcinol in Preserving Testicular Function in Streptozotocin-Induced Diabetic Rats. International Journal of Molecular Sciences. 2024; 25(8):4316. https://doi.org/10.3390/ijms25084316
Chicago/Turabian StyleOh, Ji-Hyeon, Je-Yong Choi, Dae-Won Kim, Seong-Gon Kim, and Umberto Garagiola. 2024. "Therapeutic Potential of 4-Hexylresorcinol in Preserving Testicular Function in Streptozotocin-Induced Diabetic Rats" International Journal of Molecular Sciences 25, no. 8: 4316. https://doi.org/10.3390/ijms25084316
APA StyleOh, J.-H., Choi, J.-Y., Kim, D.-W., Kim, S.-G., & Garagiola, U. (2024). Therapeutic Potential of 4-Hexylresorcinol in Preserving Testicular Function in Streptozotocin-Induced Diabetic Rats. International Journal of Molecular Sciences, 25(8), 4316. https://doi.org/10.3390/ijms25084316