
Evolutionary Conservation in Protein–Protein Interactions and Structures of the Elongator Sub-Complex ELP456 from Arabidopsis and Yeast

Abstract
1. Introduction
2. Results
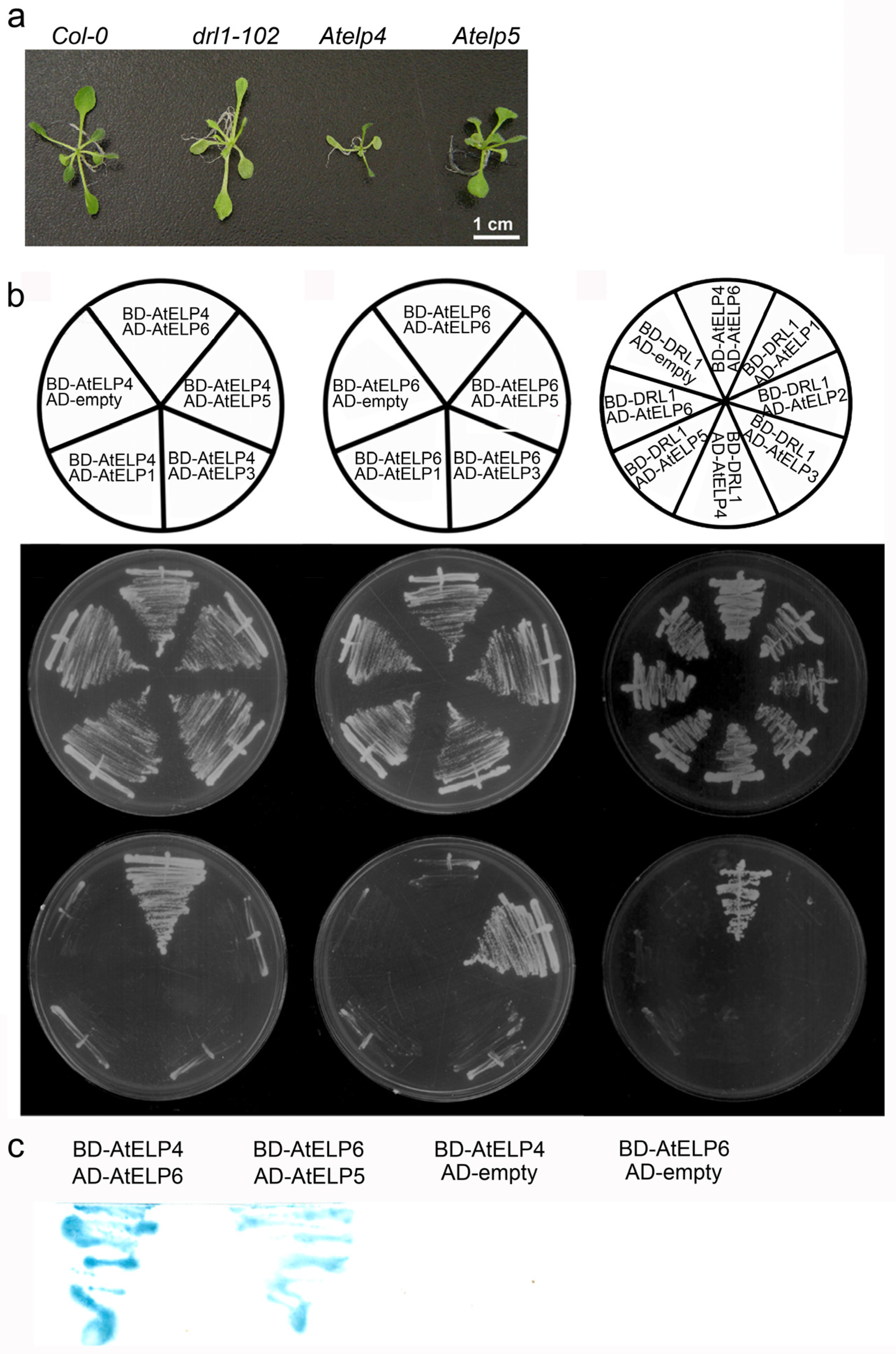
2.1. AtELP4 Interacts with AtELP6 but Not Directly with AtELP5
2.2. AtDRL1 Does Not Directly Interact with Each Subunit of the AtELP456 Sub-Complex
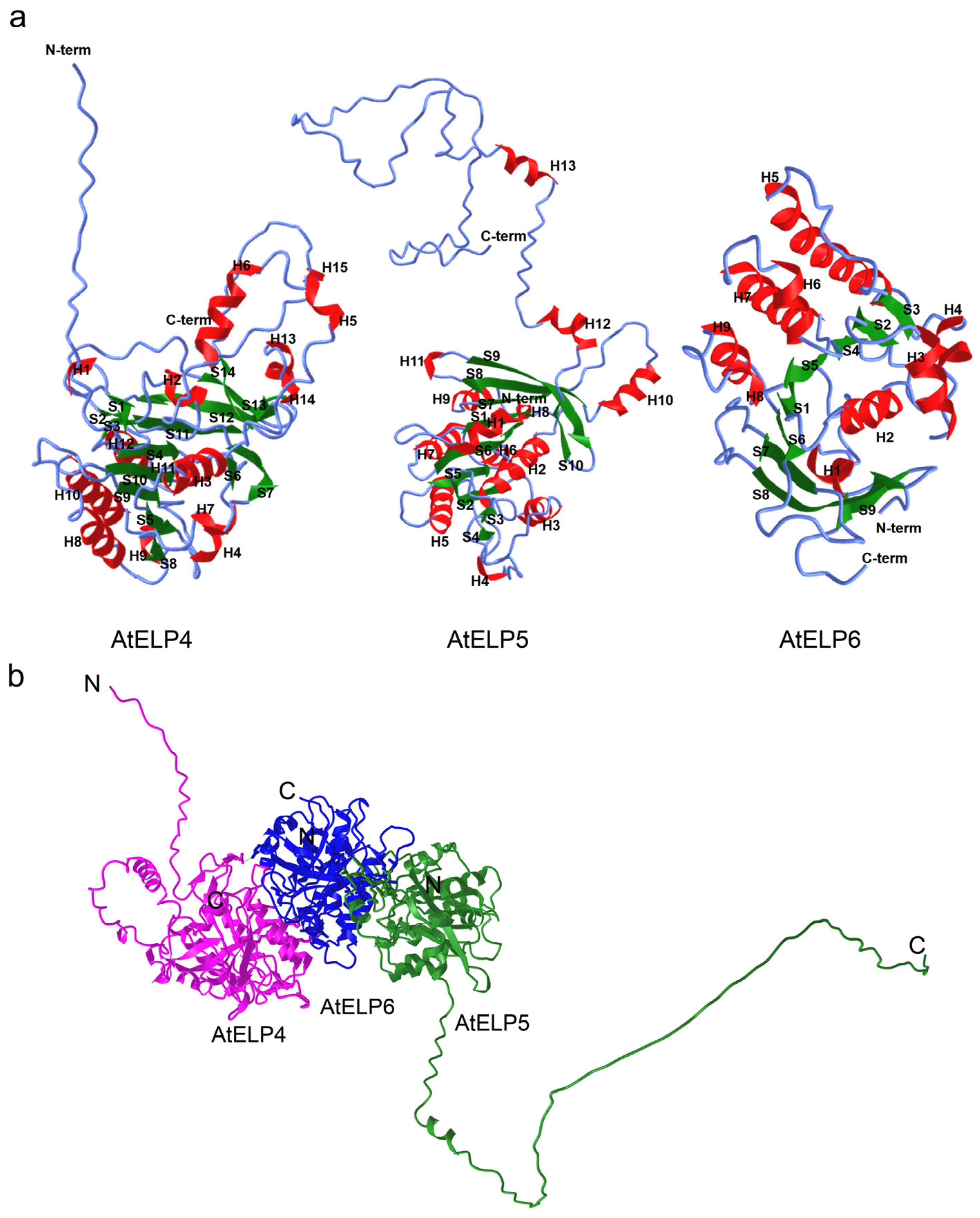
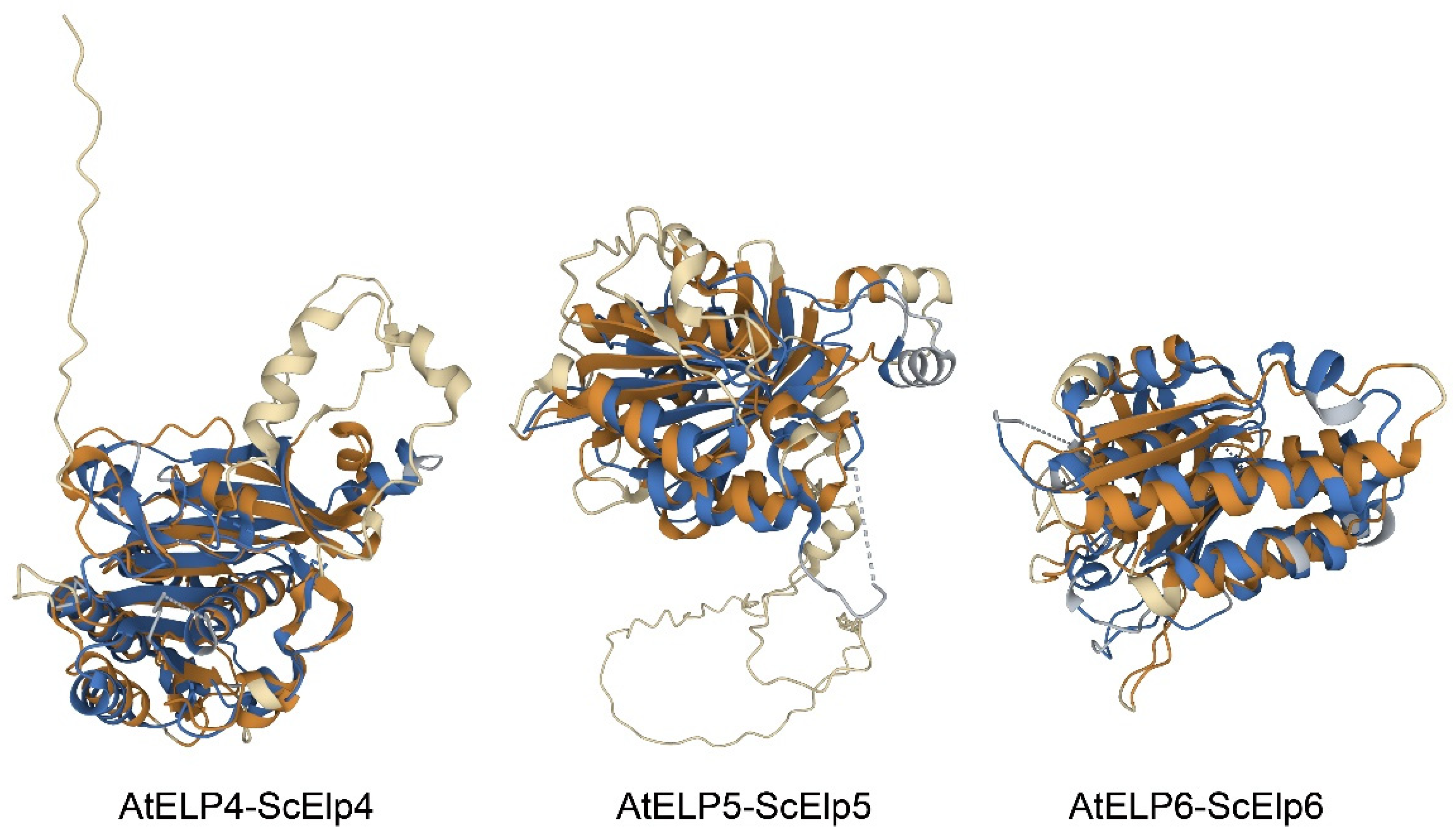
2.3. Structure Analysis of Arabidopsis ELP4, ELP5, and ELP6 Subunits


2.4. Interaction Prediction between AtELP4, AtELP5, and AtELP6 Proteins
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Yeast Two-Hybrid Assay and Yeast Growth Conditions
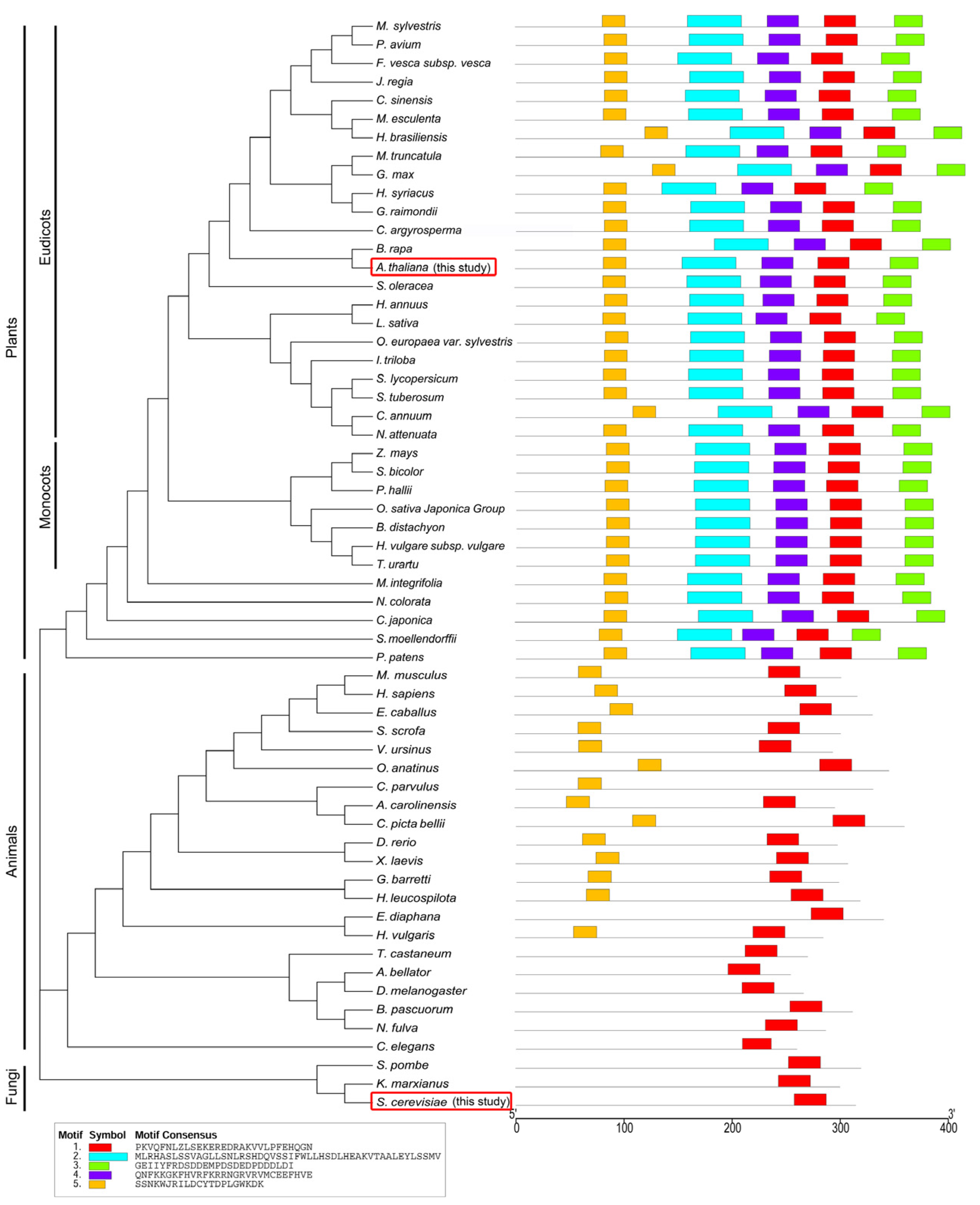
4.3. Phylogeny Relationship and Motif Analysis
4.4. Structure Prediction
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Otero, G.; Fellows, J.; Li, Y.; de Bizemont, T.; Dirac, A.M.G.; Gustafsson, C.M.; Erdjument-Bromage, H.; Tempst, P.; Svejstrup, J.Q. Elongator, a Multisubunit Component of a Novel RNA Polymerase II Holoenzyme for Transcriptional Elongation. Mol. Cell 1999, 3, 109–118. [Google Scholar] [CrossRef]
- Dauden, M.I.; Jaciuk, M.; Weis, F.; Lin, T.-Y.; Kleindienst, C.; Abbassi, N.E.H.; Khatter, H.; Krutyhołowa, R.; Breunig, K.D.; Kosinski, J.; et al. Molecular Basis of tRNA Recognition by the Elongator Complex. Sci. Adv. 2019, 5, eaaw2326. [Google Scholar] [CrossRef] [PubMed]
- Glatt, S.; Zabel, R.; Kolaj-Robin, O.; Onuma, O.F.; Baudin, F.; Graziadei, A.; Taverniti, V.; Lin, T.-Y.; Baymann, F.; Séraphin, B.; et al. Structural Basis for tRNA Modification by Elp3 from Dehalococcoides mccartyi. Nat. Struct. Mol. Biol. 2016, 23, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-Y.; Abbassi, N.E.H.; Zakrzewski, K.; Chramiec-Głąbik, A.; Jemioła-Rzemińska, M.; Różycki, J.; Glatt, S. The Elongator Subunit Elp3 is a non-Canonical tRNA Acetyltransferase. Nat. Commun. 2019, 10, 625. [Google Scholar] [CrossRef] [PubMed]
- Wittschieben, B.Ø.; Otero, G.; de Bizemont, T.; Fellows, J.; Erdjument-Bromage, H.; Ohba, R.; Li, Y.; Allis, C.D.; Tempst, P.; Svejstrup, J.Q. A Novel Histone Acetyltransferase Is an Integral Subunit of Elongating RNA Polymerase II Holoenzyme. Mol. Cell 1999, 4, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Fazly, A.M.; Zhou, H.; Huang, S.; Zhang, Z.; Stillman, B. The Elongator Complex Interacts with PCNA and Modulates Transcriptional Silencing and Sensitivity to DNA Damage Agents. PLOS Genet. 2009, 5, e1000684. [Google Scholar] [CrossRef]
- Rahl, P.B.; Chen, C.Z.; Collins, R.N. Elp1p, the Yeast Homolog of the FD Disease Syndrome Protein, Negatively Regulates Exocytosis Independently of Transcriptional Elongation. Mol. Cell 2005, 17, 841–853. [Google Scholar] [CrossRef]
- Woloszynska, M.; Le Gall, S.; Van Lijsebettens, M. Plant Elongator-Mediated Transcriptional Control in a Chromatin and Epigenetic Context. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2016, 1859, 1025–1033. [Google Scholar] [CrossRef]
- Grossenbacher, A.M.; Stadelmann, B.; Heyer, W.D.; Thuriaux, P.; Kohli, J.; Smith, C.; Agris, P.F.; Kuo, K.C.; Gehrke, C. Antisuppressor Mutations and Sulfur-Carrying Nucleosides in transfer RNAs of Schizosaccharomyces pombe. J. Biol. Chem. 1986, 261, 16351–16355. [Google Scholar] [CrossRef]
- Heyer, W.D.; Thuriaux, P.; Kohli, J.; Ebert, P.; Kersten, H.; Gehrke, C.; Kuo, K.C.; Agris, P.F. An Antisuppressor Mutation of Schizosaccharomyces pombe Affects the Post-Transcriptional Modification of the “wobble” Base in the Anticodon of tRNAs. J. Biol. Chem. 1984, 259, 2856–2862. [Google Scholar] [CrossRef]
- Schaffrath, R.; Leidel, S.A. Wobble Uridine Modifications—A Reason to Live, a Reason to Die?! RNA Biol. 2017, 14, 1209–1222. [Google Scholar] [CrossRef]
- Karlsborn, T.; Tükenmez, H.; Mahmud, A.K.M.F.; Xu, F.; Xu, H.; Byström, A.S. Elongator, a conserved complex required for wobble uridine modifications in Eukaryotes. RNA Biol. 2014, 11, 1519–1528. [Google Scholar] [CrossRef]
- Bednářová, A.; Hanna, M.; Durham, I.; VanCleave, T.; England, A.; Chaudhuri, A.; Krishnan, N. Lost in Translation: Defects in Transfer RNA Modifications and Neurological Disorders. Front. Mol. Neurosci. 2017, 10, 135. [Google Scholar] [CrossRef] [PubMed]
- Gaik, M.; Kojic, M.; Stegeman, M.R.; Öncü-Öner, T.; Kościelniak, A.; Jones, A.; Mohamed, A.; Chau, P.Y.S.; Sharmin, S.; Chramiec-Głąbik, A.; et al. Functional Divergence of the two Elongator Subcomplexes during Neurodevelopment. EMBO Mol. Med. 2022, 14, e15608. [Google Scholar] [CrossRef]
- Kojic, M.; Gawda, T.; Gaik, M.; Begg, A.; Salerno-Kochan, A.; Kurniawan, N.D.; Jones, A.; Drożdżyk, K.; Kościelniak, A.; Chramiec-Głąbik, A.; et al. Elp2 Mutations Perturb the Epitranscriptome and Lead to a Complex Neurodevelopmental Phenotype. Nat. Commun. 2021, 12, 2678. [Google Scholar] [CrossRef]
- Pereira, M.; Francisco, S.; Varanda, A.S.; Santos, M.; Santos, M.A.S.; Soares, A.R. Impact of tRNA Modifications and tRNA-Modifying Enzymes on Proteostasis and Human Disease. Int. J. Mol. Sci. 2018, 19, 3738. [Google Scholar] [CrossRef] [PubMed]
- Kojic, M.; Wainwright, B. The Many Faces of Elongator in Neurodevelopment and Disease. Front. Mol. Neurosci. 2016, 9, 115. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.J.; Xu, F.; Byström, A.S. Elongator—A tRNA modifying complex that promotes efficient translational decoding. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2018, 1861, 401–408. [Google Scholar] [CrossRef]
- Krogan, N.J.; Greenblatt, J.F. Characterization of a Six-Subunit Holo-Elongator Complex Required for the Regulated Expression of a Group of Genes in Saccharomyces cerevisiae. Mol. Cell. Biol. 2001, 21, 8203–8212. [Google Scholar] [CrossRef]
- Krutyhołowa, R.; Hammermeister, A.; Zabel, R.; Abdel-Fattah, W.; Reinhardt-Tews, A.; Helm, M.; Stark, M.J.R.; Breunig, K.D.; Schaffrath, R.; Glatt, S. Kti12, a PSTK-like tRNA dependent ATPase essential for tRNA modification by Elongator. Nucleic Acids Res. 2019, 47, 4814–4830. [Google Scholar] [CrossRef]
- Li, Y.; Takagi, Y.; Jiang, Y.; Tokunaga, M.; Erdjument-Bromage, H.; Tempst, P.; Kornberg, R.D. A Multiprotein Complex That Interacts with RNA Polymerase II Elongator*. J. Biol. Chem. 2001, 276, 29628–29631. [Google Scholar] [CrossRef]
- Winkler, G.S.; Petrakis, T.G.; Ethelberg, S.; Tokunaga, M.; Erdjument-Bromage, H.; Tempst, P.; Svejstrup, J.Q. RNA Polymerase II Elongator Holoenzyme Is Composed of Two Discrete Subcomplexes. J. Biol. Chem. 2001, 276, 32743–32749. [Google Scholar] [CrossRef]
- Frohloff, F.; Jablonowski, D.; Fichtner, L.; Schaffrath, R. Subunit Communications Crucial for the Functional Integrity of the Yeast RNA Polymerase II Elongator (γ-Toxin Target (TOT)) Complex. J. Biol. Chem. 2003, 278, 956–961. [Google Scholar] [CrossRef]
- Glatt, S.; Létoquart, J.; Faux, C.; Taylor, N.M.I.; Séraphin, B.; Müller, C.W. The Elongator subcomplex Elp456 is a hexameric RecA-like ATPase. Nat. Struct. Mol. Biol. 2012, 19, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Zhao, W.; Diao, W.; Xie, X.; Wang, Z.; Zhang, J.; Shen, Y.; Long, J. Crystal Structure of Elongator Subcomplex Elp4–6*. J. Biol. Chem. 2012, 287, 21501–21508. [Google Scholar] [CrossRef] [PubMed]
- Dauden, M.I.; Kosinski, J.; Kolaj-Robin, O.; Desfosses, A.; Ori, A.; Faux, C.; Hoffmann, N.A.; Onuma, O.F.; Breunig, K.D.; Beck, M.; et al. Architecture of the yeast Elongator complex. EMBO Rep. 2017, 18, 264–279. [Google Scholar] [CrossRef]
- Setiaputra, D.T.; Cheng, D.T.; Lu, S.; Hansen, J.M.; Dalwadi, U.; Lam, C.H.; To, J.L.; Dong, M.-Q.; Yip, C.K. Molecular architecture of the yeast Elongator complex reveals an unexpected asymmetric subunit arrangement. EMBO Rep. 2017, 18, 280–291. [Google Scholar] [CrossRef]
- Nelissen, H.; De Groeve, S.; Fleury, D.; Neyt, P.; Bruno, L.; Bitonti, M.B.; Vandenbussche, F.; Van Der Straeten, D.; Yamaguchi, T.; Tsukaya, H.; et al. Plant Elongator regulates auxin-related genes during RNA polymerase II transcription elongation. Proc. Natl. Acad. Sci. USA 2010, 107, 1678–1683. [Google Scholar] [CrossRef]
- Zhou, X.; Hua, D.; Chen, Z.; Zhou, Z.; Gong, Z. Elongator mediates ABA responses, oxidative stress resistance and anthocyanin biosynthesis in Arabidopsis. Plant J. 2009, 60, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Mou, Z. Elongator and its epigenetic role in plant development and responses to abiotic and biotic stresses. Front. Plant Sci. 2015, 6, 296. [Google Scholar] [CrossRef]
- Jun, S.E.; Cho, K.-H.; Manzoor, M.A.; Hwang, T.Y.; Kim, Y.S.; Schaffrath, R.; Kim, G.-T. AtELP4 a subunit of the Elongator complex in Arabidopsis, mediates cell proliferation and dorsoventral polarity during leaf morphogenesis. Front. Plant Sci. 2022, 13, 1033358. [Google Scholar] [CrossRef]
- Nakai, Y.; Horiguchi, G.; Iwabuchi, K.; Harada, A.; Nakai, M.; Hara-Nishimura, I.; Yano, T. tRNA Wobble Modification Affects Leaf Cell Development in Arabidopsis thaliana. Plant Cell Physiol. 2019, 60, 2026–2039. [Google Scholar] [CrossRef]
- Cho, H.; Cho, H.S.; Nam, H.; Jo, H.; Yoon, J.; Park, C.; Dang, T.V.T.; Kim, E.; Jeong, J.; Park, S.; et al. Translational control of phloem development by RNA G-quadruplex–JULGI determines plant sink strength. Nat. Plants 2018, 4, 376–390. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.E.; Cho, K.-H.; Hwang, J.-Y.; Abdel-Fattah, W.; Hammermeister, A.; Schaffrath, R.; Bowman, J.L.; Kim, G.-T. Comparative Analysis of the Conserved Functions of Arabidopsis DRL1 and Yeast KTI12. Mol. Cells 2015, 38, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Mehlgarten, C.; Prochaska, H.; Hammermeister, A.; Abdel-Fattah, W.; Wagner, M.; Krutyhołowa, R.; Jun, S.E.; Kim, G.-T.; Glatt, S.; Breunig, K.D.; et al. Use of a Yeast tRNase Killer Toxin to Diagnose Kti12 Motifs Required for tRNA Modification by Elongator. Toxins 2017, 9, 272. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.-H.; Choi, H.; Seki, M.; Jun, S.E.; Yi, Y.B.; Shinozaki, K.; Tsukaya, H.; Kim, G.-T. DRL1 regulates adaxial leaf patterning and shoot apical meristem activity in Arabidopsis. J. Plant Biol. 2007, 50, 467–474. [Google Scholar] [CrossRef]
- Petrakis, T.G.; Søgaard, T.M.M.; Erdjument-Bromage, H.; Tempst, P.; Svejstrup, J.Q. Physical and Functional Interaction between Elongator and the Chromatin-associated Kti12 Protein *. J. Biol. Chem. 2005, 280, 19454–19460. [Google Scholar] [CrossRef] [PubMed]
- RCSB Protein Data Bank. 4A8J: Crystal Structure of the Elongator Subcomplex Elp456. Available online: https://www.rcsb.org/structure/4a8j (accessed on 18 October 2022).
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X Version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.; O’Neill, M.; Pritzel, A.; Antropova, N.; Senior, A.; Green, T.; Žídek, A.; Bates, R.; Blackwell, S.; Yim, J.; et al. Protein Complex Prediction with AlphaFold-Multimer. bioRxiv 2022. [Google Scholar] [CrossRef]
- Gouet, P.; Robert, X.; Courcelle, E. ESPript/ENDscript: Extracting and rendering sequence and 3D information from atomic structures of proteins. Nucleic Acids Res. 2003, 31, 3320–3323. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Y. How significant is a protein structure similarity with TM-score = 0.5? Bioinformatics 2010, 26, 889–895. [Google Scholar] [CrossRef]
- Hubbard, R.E.; Kamran Haider, M. Hydrogen Bonds in Proteins: Role and Strength. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2010; ISBN 978-0-470-01590-2. [Google Scholar]
- Mehlgarten, C.; Jablonowski, D.; Wrackmeyer, U.; Tschitschmann, S.; Sondermann, D.; Jäger, G.; Gong, Z.; Byström, A.S.; Schaffrath, R.; Breunig, K.D. Elongator function in tRNA wobble uridine modification is conserved between yeast and plants. Mol. Microbiol. 2010, 76, 1082–1094. [Google Scholar] [CrossRef] [PubMed]
- Versées, W.; De Groeve, S.; Van Lijsebettens, M. Elongator, a conserved multitasking complex? Mol. Microbiol. 2010, 76, 1065–1069. [Google Scholar] [CrossRef] [PubMed]
- RCSB Protein Data Bank. 4EJS: Structure of Yeast Elongator Subcomplex Elp456. Available online: https://www.rcsb.org/structure/4EJS (accessed on 18 October 2022).
- Esberg, A.; Huang, B.; Johansson, M.J.O.; Byström, A.S. Elevated Levels of Two tRNA Species Bypass the Requirement for Elongator Complex in Transcription and Exocytosis. Mol. Cell 2006, 24, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Jarosz, M.; Van Lijsebettens, M.; Woloszynska, M. Plant Elongator—Protein Complex of Diverse Activities Regulates Growth, Development, and Immune Responses. Int. J. Mol. Sci. 2020, 21, 6912. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Youkharibache, P.; Zhang, D.; Lanczycki, C.J.; Geer, R.C.; Madej, T.; Phan, L.; Ward, M.; Lu, S.; Marchler, G.H.; et al. iCn3D, a web-based 3D viewer for sharing 1D/2D/3D representations of biomolecular structures. Bioinformatics 2020, 36, 131–135. [Google Scholar] [CrossRef]
- Sehnal, D.; Bittrich, S.; Deshpande, M.; Svobodová, R.; Berka, K.; Bazgier, V.; Velankar, S.; Burley, S.K.; Koča, J.; Rose, A.S. Mol* Viewer: Modern web app for 3D visualization and analysis of large biomolecular structures. Nucleic Acids Res. 2021, 49, W431–W437. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast | Arabidopsis | AGI | Homology (%) | Function |
|---|---|---|---|---|
| ELP1/ELO2/TOT1/KTI7/IKI3 | AtELP1 | AT5G13680 | 27 | Apoenzyme |
| ELP2/TOT2/KTI3 | AtELP2 | AT1G49540 | 29 | Apoenzyme |
| ELP3/ELO3/TOT3/KTI8 | AtELP3 | AT5G50320 | 63 | Coenzyme; histone acetyl transferase |
| ELP4/ELO1/TOT7/KTI9 | AtELP4 | AT3G11220 | 20 | Apoenzyme |
| TLP5/TOT5/IKI1 | AtELP5 | AT2G18410 | 12 | Apoenzyme |
| ELP6/TOT6/KEI4 | AtELP6 | AT4G10090 | 16 | Apoenzyme |
| ELO4/KTI12/TOT4 | DRL1 | AT1G13870 | 28 |
| Protein | Reference (Arabidopsis) | Target (Yeast) | RMSD | TM-Score | Sequence Identity | Equivalent Residues | Reference Coverage | Target Coverage |
|---|---|---|---|---|---|---|---|---|
| ELP4 | AF-Q9C778 | 4A8J_A | 3.08 | 0.65 | 20% | 261 | 74% | 94% |
| ELP5 | AF-F4IQJ2 | 4A8J_B | 3.05 | 0.43 | 11% | 187 | 50% | 85% |
| ELP6 | AF-Q8L9Y2 | 4A8J_C | 3.05 | 0.74 | 14% | 230 | 88% | 87% |
| In the Interface between AtELP4–AtELP6 | In the Interface between ScElp4–ScElp6 | In the Interface between AtELP5–AtELP6 | In the Interface between ScElp5–ScElp6 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AtELP4 | AtELP6 | Distance (nm) | ScElp4 | ScElp6 | AtELP5 | AtELP6 | Distance (nm) | ScElp5 | ScElp6 | |
| 1 | ASN(N)32:ND2 | GLU(E)250:OE1 | 0.37 | GLU(E)303:OE1 | SER(S)159:HG | TRP(W)100:O | ARG(R)119:NH2 | 0.32 | TYR(Y)36:OH | PRO(P)244:N |
| 2 | GLN(Q)180:OE1 | LYS(K)100:NZ | 0.36 | LYS(K)320:NZ | ASP(D)111:OD1 | TRP(W)100:N | TYR(Y)164:OH | 0.32 | SER(S)59:OG | ARG(R)176:NH2 |
| 3 | SER(S)231:OG | LEU(L)97:O | 0.26 | LYS(K)320:NZ | ASP(D)111:OD2 | TYR(Y)94:OH | HIS(H)166:ND1 | 0.31 | ASP(D)74:OD1 | ARG(R)176:NH1 |
| 4 | ARG(R)256:NH1 | GLU(E)147:OE2 | 0.33 | ASN(N)346:OD1 | ASN(N)186:ND2 | TYR(Y)94:OH | SER(S)170:OG | 0.36 | ASP(D)74:OD2 | ARG(R)176:NE |
| 5 | ARG(R)256:NH2 | GLU(E)147:OE2 | 0.20 | ASN(N)346:OD1 | GLN(Q)150:NH2 | ASP(D)231:OD1 | SER(S)237:OG | 0.27 | ASP(D)74:OD2 | ARG(R)176:NH1 |
| 6 | HIS(H)259:ND1 | HIS(H)181:ND1 | 0.36 | ASN(N)346:OD1 | GLN(Q)150:O | ASP(D)231:OD2 | SER(S)237:OG | 0.37 | TYR(Y)111:OH | ASN(N)167:ND2 |
| 7 | HIS(H)259:NE2 | ASP(D)183:OD | 0.26 | ASN(N)346:ND2 | GLN(Q)150:NE2 | PHE(F)232:N | ASN(N)239:O | 0.34 | TYR(Y)111:OH | SER(S)210:OG |
| 8 | ASN(N)346:ND2 | GLN(Q)150:OE1 | GLN(Q)230:O | LEU(L)241:N | 0.26 | LYS(K)140:O | ASN(N)206:ND2 | |||
| 9 | GLN(Q)230:N | LEU(L)241:O | 0.28 | LYS(K)140:N | ASN(N)206:ND2 | |||||
| 10 | VAL(V)228:O | ASN(N)243:N | 0.30 | LYS(K)140:N | ASN(N)206:O | |||||
| 11 | HIS(H)229:NE2 | TYR(Y)257:OH | 0.29 | LYS(K)140:NZ | GLN(Q)205:OE1 | |||||
| 12 | ARG(R)195:NH1 | GLU(E)258:OE1 | ||||||||
| 13 | ARG(R)195:NH2 | GLU(E)258:OE2 | ||||||||
| 14 | ASN(N)198:OD1 | ARG(R)240:NH1 | ||||||||
| 15 | ASN(N)198:ND2 | ARG(R)240:NH1 | ||||||||
| 16 | ASN(N)199:OD1 | ARG(R)240:NH2 | ||||||||
| 17 | ASN(N)199:OD1 | ARG(R)240:NH1 | ||||||||
| In the Interface between AtELP4–AtELP5 | In the Interface between ScElp4–ScElp5 | ||||
|---|---|---|---|---|---|
| AtELP4 | AtELP5 | Distance(nm) | ScElp4 | ScElp5 | |
| 1 | PRO(P)211:O | PRO(P)163:N | 0.38 | GLU(E)117:N | SER(S)212:O |
| 2 | PRO(P)211:O | SER(S)166:OG | 0.38 | THR(T)116:O | GLY(G)213:O |
| 3 | SER(S)247:O | ASN(N)196:ND2 | 0.30 | THR(T)116:O | GLY(G)213:N |
| 4 | HIS(H)68:NE2 | GLY(G)246:O | 0.29 | THR(T)116:N | ARG(R)214:N |
| 5 | TYR(Y)284:O | ARG(R)247:NH1 | 0.36 | THR(T)337:OG1 | ASN(N)153:O |
| 6 | ILE(I)270:O | VAL(V)248:N | 0.31 | SER(S)304:OG | ASN(N)153:ND2 |
| 7 | ASP(D)272:N | VAL(V)248:O | 0.26 | SER(S)304:N | ASN(N)153:ND2 |
| 8 | LYS(K)268:NZ | SER(S)279:O | 0.33 | PHE(F)302:O | ASN(N)153:ND2 |
| Name | Sequence | Tm Value (°C) | Purpose |
|---|---|---|---|
| ELP1-forward | CACCATGAAAAGAGATGAAGATTTGAC | 51.3 | cloning |
| ELP1-reverse | TCATGGGCTTATGAAGACCT | 53.4 | cloning |
| ELP2-forward | CACCATGTCAGAAAACACAAAAGTCGA | 53.1 | cloning |
| ELP2-reverse | TCAAAACTTGAAGTTAAAAACTCTC | 52.4 | cloning |
| ELP3-forward | CACCATGGCGACGGCGGTAG | 54.0 | cloning |
| ELP3-reverse | TCAAAGAAGATGCTTCACCA | 51.3 | cloning |
| ELP4-forward | CACCATGGCTGCACCAAACGTTC | 54.9 | cloning |
| ELP4-reverse | TCAAAAATCTAGTGCTCCGG | 53.4 | cloning |
| ELP5-forward | CACCATGGCGGAATCGATTTTCAG | 53.4 | cloning |
| ELP5-reverse | TTAAATGTCCAAATCATCATCAGGA | 54.0 | cloning |
| ELP6-forward | CACCATGGATCGTTCTTTGAATCTC | 51.9 | cloning |
| ELP6-reverse | TCAGCTTCTGCAACCAGGAT | 55.4 | cloning |
| DRL1-forward | CACCATGGCGCTAGTTGTGATTTG | 53.4 | cloning |
| DRL1-reverse | TCAAGCGTTATTACCTCCAAAC | 54.4 | cloning |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jun, S.E.; Cho, K.-H.; Schaffrath, R.; Kim, G.-T. Evolutionary Conservation in Protein–Protein Interactions and Structures of the Elongator Sub-Complex ELP456 from Arabidopsis and Yeast. Int. J. Mol. Sci. 2024, 25, 4370. https://doi.org/10.3390/ijms25084370
Jun SE, Cho K-H, Schaffrath R, Kim G-T. Evolutionary Conservation in Protein–Protein Interactions and Structures of the Elongator Sub-Complex ELP456 from Arabidopsis and Yeast. International Journal of Molecular Sciences. 2024; 25(8):4370. https://doi.org/10.3390/ijms25084370
Chicago/Turabian StyleJun, Sang Eun, Kiu-Hyung Cho, Raffael Schaffrath, and Gyung-Tae Kim. 2024. "Evolutionary Conservation in Protein–Protein Interactions and Structures of the Elongator Sub-Complex ELP456 from Arabidopsis and Yeast" International Journal of Molecular Sciences 25, no. 8: 4370. https://doi.org/10.3390/ijms25084370
APA StyleJun, S. E., Cho, K.-H., Schaffrath, R., & Kim, G.-T. (2024). Evolutionary Conservation in Protein–Protein Interactions and Structures of the Elongator Sub-Complex ELP456 from Arabidopsis and Yeast. International Journal of Molecular Sciences, 25(8), 4370. https://doi.org/10.3390/ijms25084370