
The Catalysis Mechanism of E. coli Nitroreductase A, a Candidate for Gene-Directed Prodrug Therapy: Potentiometric and Substrate Specificity Studies


 , , and
, , and
Abstract
:1. Introduction
2. Results
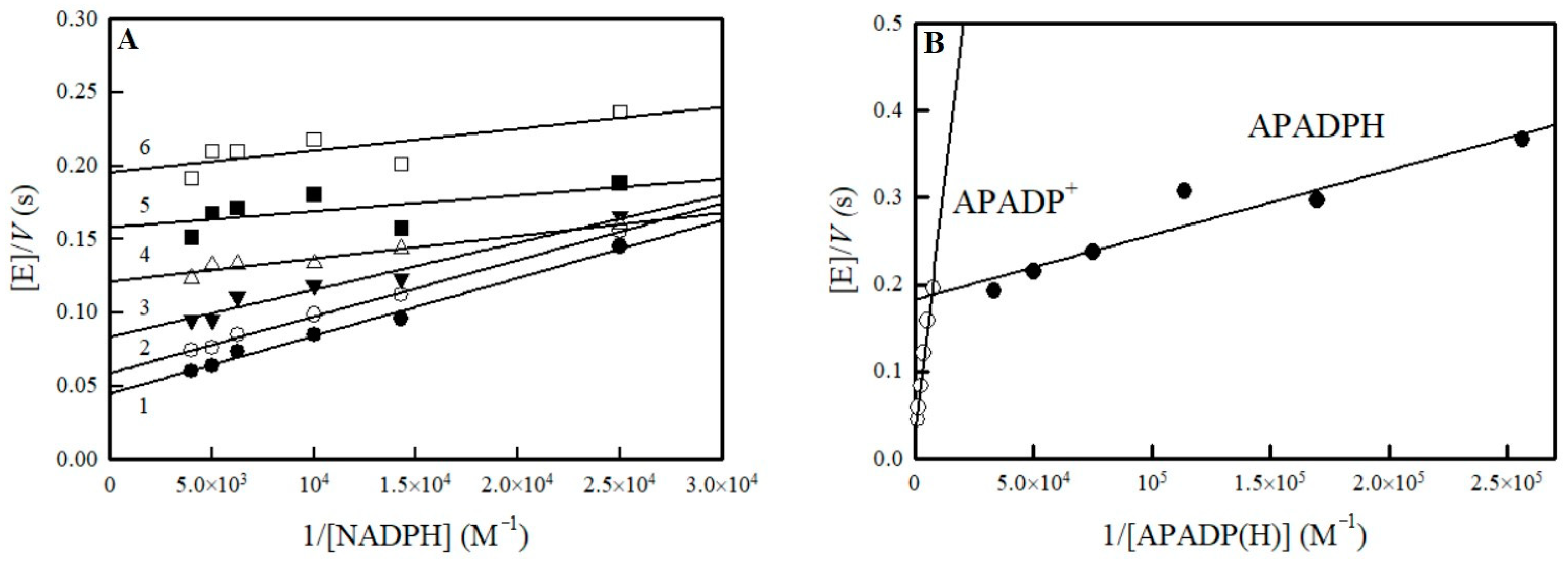
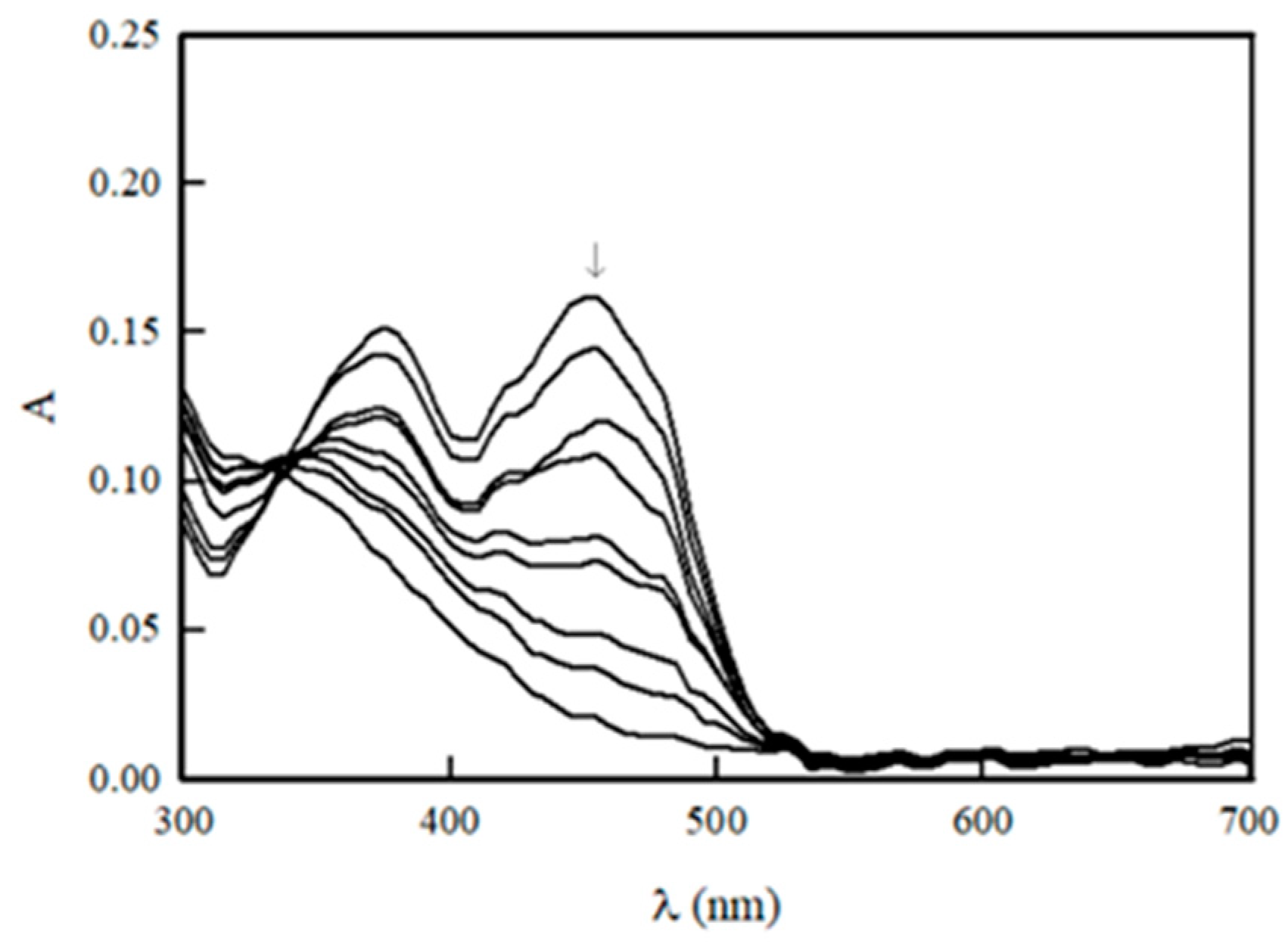
2.1. Potentiometric Studies of NfsA
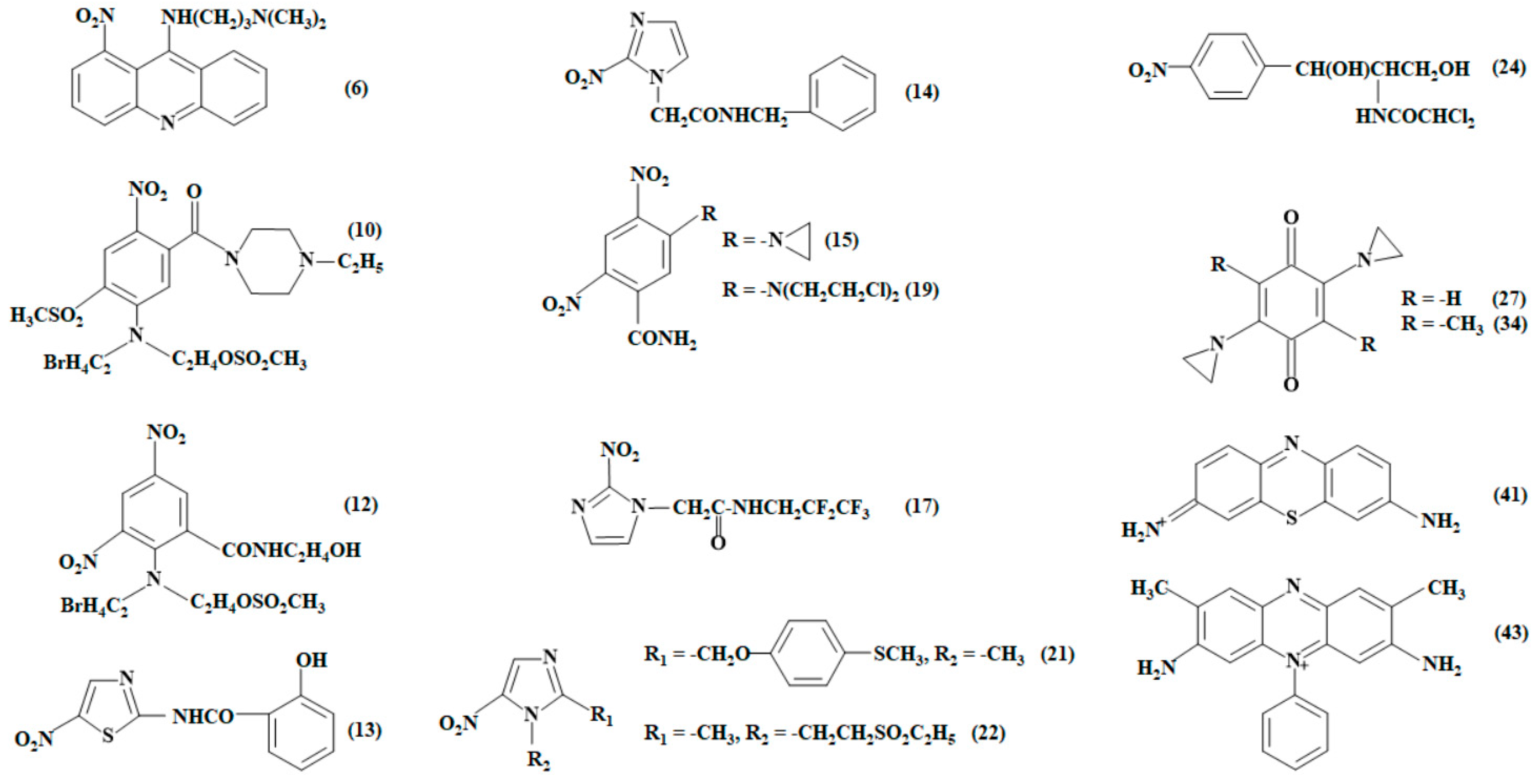
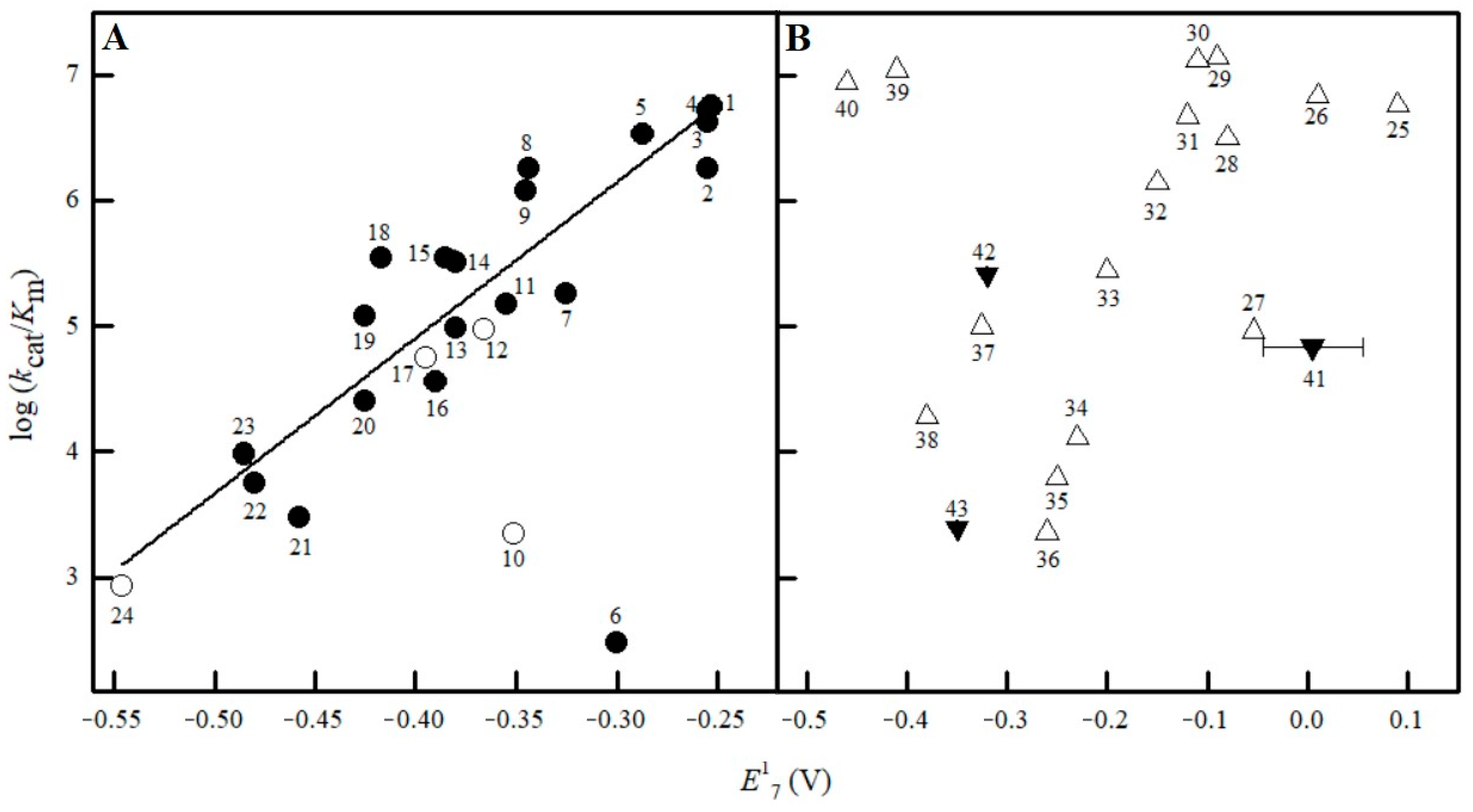
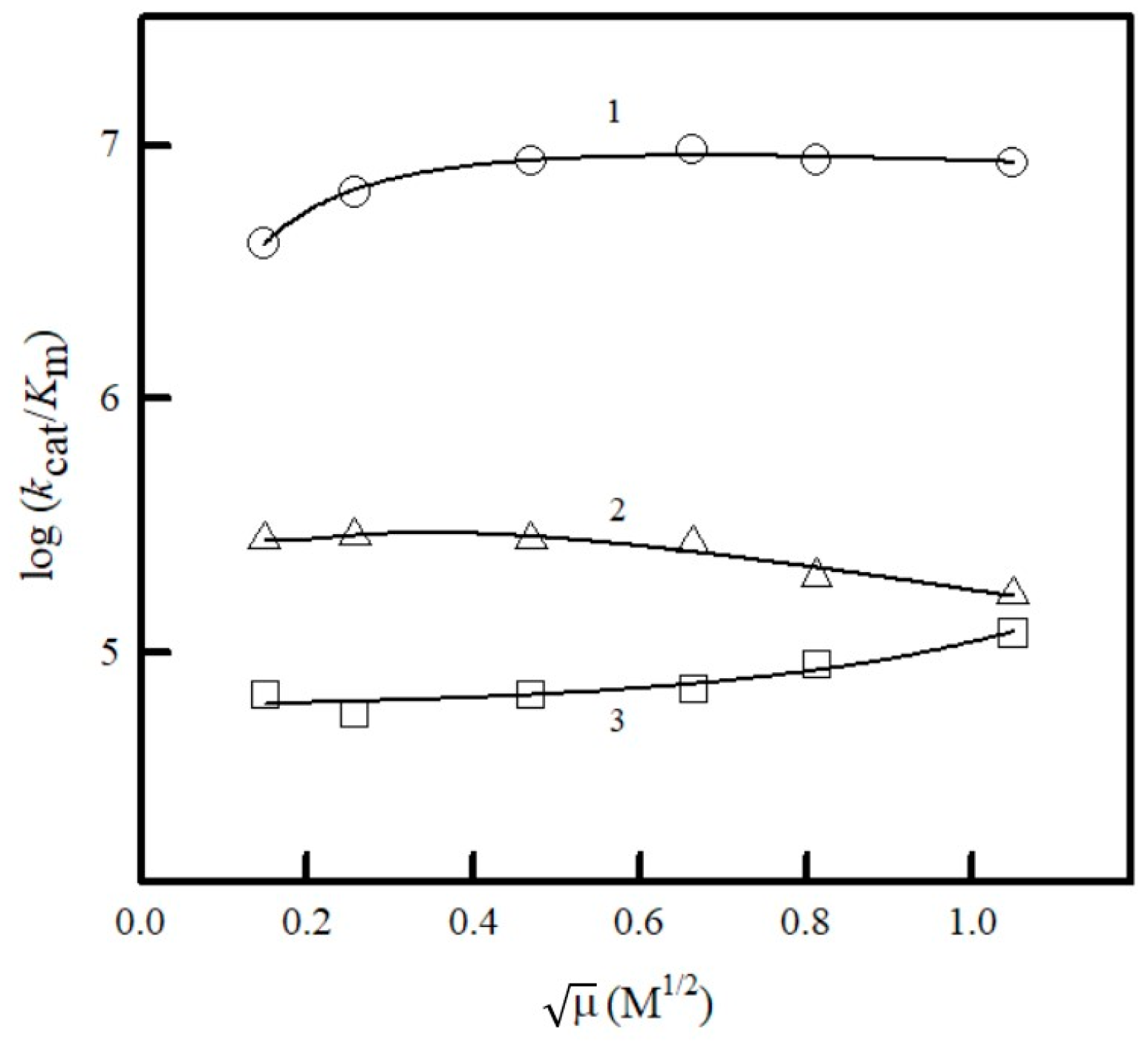
2.2. Studies of NfsA Oxidant Specificity

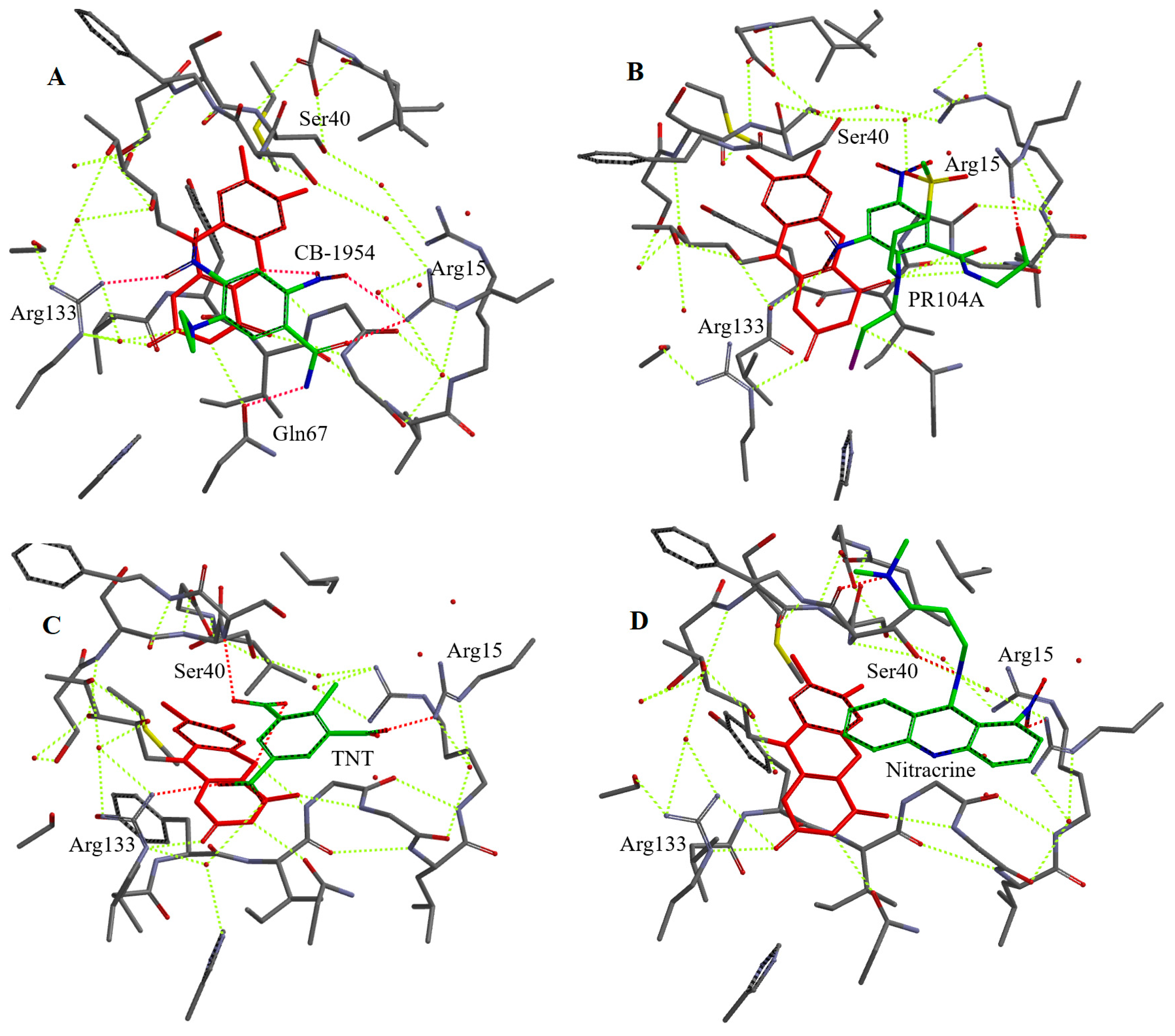
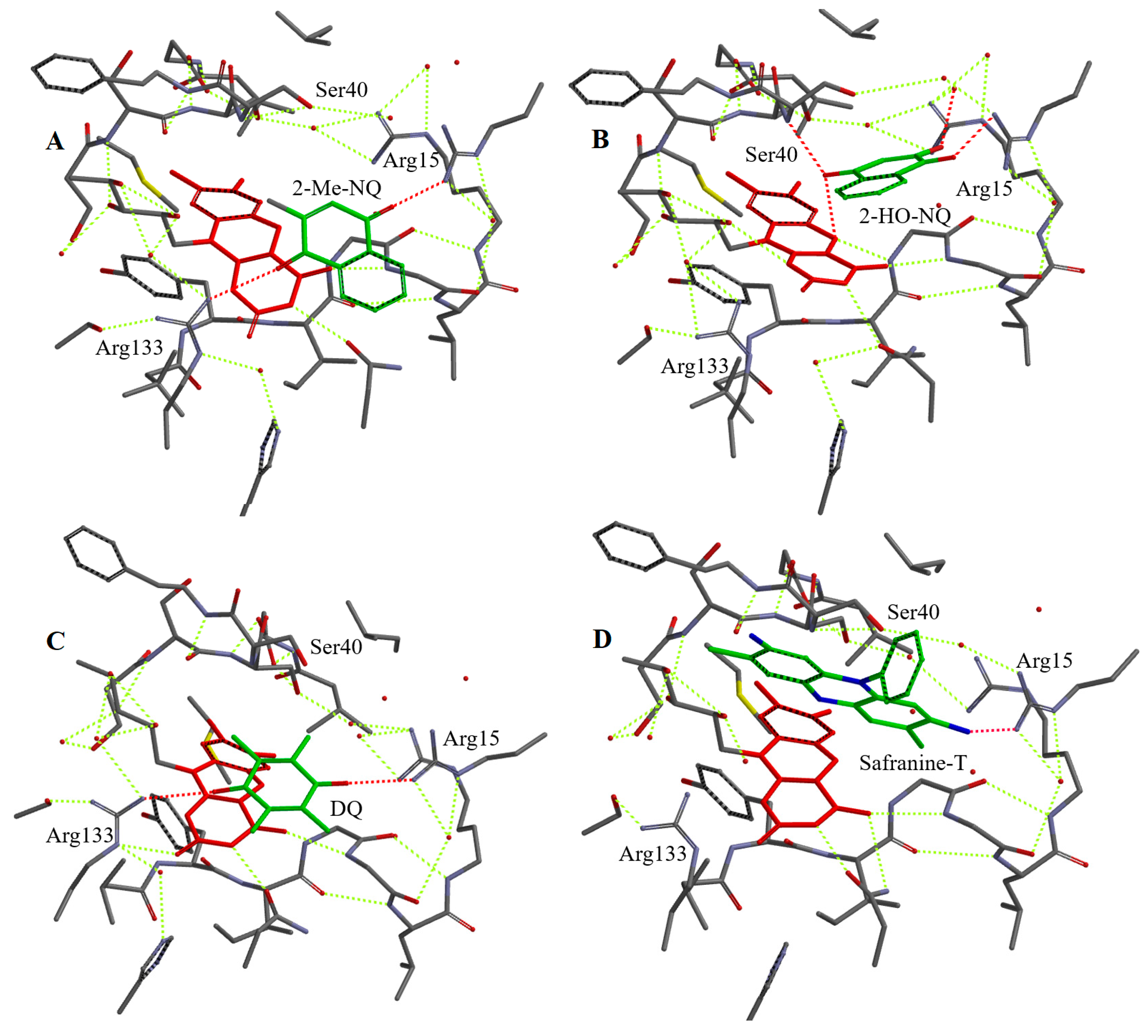
2.3. Molecular Modelling of Binding of Oxidants to Reduced NfsA
3. Discussion
4. Materials and Methods
4.1. Enzymes and Reagents
4.2. Steady-State Kinetic Studies of NfsA
4.3. Photoreduction of NfsA
4.4. Computer Modelling and Calculations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APADP(H) | 3-acetylpyridine adenine dinucleotide phosphate |
| ArNHOH | Hydroxylamine |
| ArNO | Nitroso compound |
| ArNO2 | Nitroaromatic compound |
| E07 | Midpoint potential of oxidized/reduced flavin redox couple |
| E17 | Single-electron reduction potential |
| E7(H−) | Hydride-transfer potential |
| FMN | Flavin mononucleotide |
| GDEPT | Gene-directed prodrug therapy |
| kcat | Catalytic constant |
| kcat/Km | Bimolecular rate constant |
| Ki | Inhibition constant |
| log D | Octanol/water distribution coefficient at pH 7.0 |
| MMFF | Molecular mechanics force field |
| NfsA | Escherichia coli nitroreductase A |
| NfsB | Escherichia coli nitroreductase B |
| NR | Nitroreductase |
| Q | Quinone |
| VdWvol | Van der Waals volume |
| ΔH≠ | Activation enthalpy |
| ΔS≠ | Activation entropy |
References
- Spain, J.C. Biodegradation of nitroaromatic compounds. Annu. Rev. Microbiol. 1995, 49, 523–555. [Google Scholar] [CrossRef] [PubMed]
- Purohit, V.; Basu, A.K. Mutagenicity of nitroaromatic compounds. Chem. Res. Toxicol. 2000, 13, 674–692. [Google Scholar] [CrossRef] [PubMed]
- Roldan, M.D.; Perez-Reinado, E.; Castillo, F.; Moreno-Vivian, E. Reduction of polynitroaromatic compounds: The bacterial nitroreductases. FEMS Microbiol. Rev. 2008, 32, 474–500. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Mishra, K.; Ramanathan, G. Bioremediation of nitroaromatic compounds. In Wastewater Treatment Engineering; Samer, M., Ed.; IntechOpen: London, UK, 2015; pp. 51–83. [Google Scholar]
- Nepali, K.; Lee, H.-J.; Liou, J.-P. Nitro-group-containing drugs. J. Med. Chem. 2019, 62, 2851–2893. [Google Scholar] [CrossRef] [PubMed]
- Čėnas, N.; Nemeikaitė-Čėnienė, A.; Kosychova, L. Single- and two-electron reduction of nitroaromatic compounds by flavoenzymes: Mechanisms and implications for cytotoxicity. Int. J. Mol. Sci. 2021, 22, 8534. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wu, K.; Zhang, D.; Sherman, M.; Knox, R.; Yang, C.S. Molecular characterization of binding of substrates and inhibitors to DT-diaphorase: Combined approach involving site-directed mutagenesis, inhibitor-binding analysis, and computer modeling. Mol. Pharmacol. 1999, 56, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Knox, R.J.; Chen, S. Quinone reductase-mediated nitro-reduction: Clinical applications. Methods Enzymol. 2004, 382, 194–220. [Google Scholar] [PubMed]
- Misevičienė, L.; Anusevičius, Ž.; Šarlauskas, J.; Čėnas, N. Reduction of nitroaromatic compounds by NAD(P)H:quinone oxidoreductase (NQO1): The role of electron-accepting potency and structural parameters in the substrate specificity. Acta Biochim. Pol. 2006, 53, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Denny, W.A. Nitroreductase-based GDEPT. Curr. Pharm. Des. 2002, 8, 1349–1361. [Google Scholar] [CrossRef]
- Williams, E.M.; Little, R.F.; Mowday, A.M.; Rich, M.H.; Chan-Hyams, J.V.; Copp, J.N.; Smaill, J.B.; Patterson, A.V.; Ackerley, D.F. Nitroreductase gene-directed enzyme prodrug therapy: Insights and advances towards clinical utility. Biochem. J. 2015, 471, 131–153. [Google Scholar] [CrossRef]
- Akiva, E.; Copp, J.N.; Tokuriki, N.; Babbitt, P.C. Evolutionary and molecular foundations of multiple contemporary functions of the nitroreductase superfamily. Proc. Natl. Acad. Sci. USA 2017, 114, E9549–E9558. [Google Scholar] [CrossRef] [PubMed]
- Zenno, S.; Koike, H.; Kumar, A.N.; Jayaraman, R.; Tanokura, M.; Saigo, K. Biochemical characterization of NfsA, the Escherichia coli major nitroreductase exhibiting a high amino acid sequence homology to Frp, a Vibrio harveyi flavin oxidoreductase. J. Bacteriol. 1996, 178, 4508–4514. [Google Scholar] [CrossRef] [PubMed]
- Zenno, S.; Koike, H.; Tanokura, M.; Saigo, K. Gene cloning, purification, and characterization of NfsB, a minor oxygen-insensitive nitroreductase from Escherichia coli, similar in biochemical properties to FRase I, the major flavin reductase in Vibrio fischeri. J. Biochem. 1996, 120, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Nivinskas, H.; Koder, R.L.; Anusevičius, Ž.; Šarlauskas, J.; Miller, A.F.; Čėnas, N. Quantitative structure-activity relationships in two-electron reduction of nitroaromatic compounds by Enterobacter cloacae NAD(P)H:nitroreductase. Arch. Biochem. Biophys. 2001, 385, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Nivinskas, H.; Staškevičienė, S.; Šarlauskas, J.; Koder, R.L.; Miller, A.F.; Čėnas, N. Two-electron reduction of quinones by Enterobacter cloacae NAD(P)H:nitroreductase: Quantitative structure-activity relationships. Arch. Biochem. Biophys. 2002, 403, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Race, P.R.; Lovering, A.L.; Green, R.M.; Ossor, A.; White, S.A.; Searle, P.F.; Wrighton, C.J.; Hyde, E.I. Structural and mechanistic studies of Escherichia coli nitroreductase with the antibiotic nitrofurazone. Reversed binding orientations in different redox states of the enzyme. J. Biol. Chem. 2005, 280, 13256–13264. [Google Scholar] [CrossRef] [PubMed]
- Anusevičius, Ž.; Misevičiene, L.; Šarlauskas, J.; Rouhier, N.; Jacquot, J.-P.; Čėnas, N. Quinone- and nitroreductase reactions of Thermotoga maritima peroxiredoxin-nitroreductase hybrid enzyme. Arch. Biochem. Biophys. 2012, 528, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Pitsawong, W.; Hoben, J.P.; Miller, A.-F. Understanding the broad substrate repertoire of nitroreductase based on its kinetic mechanism. J. Biol. Chem. 2014, 289, 15203–15214. [Google Scholar] [CrossRef] [PubMed]
- Valiauga, B.; Williams, E.M.; Ackerley, D.F.; Čėnas, N. Reduction of quinones and nitroaromatic compounds by Escherichia coli nitroreductase A (NfsA): Characterization of kinetics and substrate specificity. Arch. Biochem. Biophys. 2017, 624, 14–22. [Google Scholar] [CrossRef]
- Day, M.A.; Jarrom, D.; Christofferson, A.J.; Graziano, A.E.; Anderson, J.L.R.; Searle, P.F.; Hyde, E.I.; White, S.A. The structures of E. coli NfsA bound to the antibiotic nitrofurantoin, to 1,4-benzoquinone and to FMN. Biochem. J. 2021, 478, 2601–2617. [Google Scholar] [CrossRef]
- Day, M.A.; Christofferson, A.J.; Anderson, J.L.R.; Vass, S.O.; Evans, A.; Searle, P.F.; White, S.A.; Hyde, E.I. Structure and dynamics of three Escherichia coli NfsB nitro-reductase mutants selected for enhanced activity with the cancer prodrug CB1954. Int. J. Molec. Sci. 2023, 24, 5987. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, G.; Skelly, J.; Neidle, S. Crystal structure of FMN-dependent nitroreductase from Escherichia coli B: A prodrug-activating enzyme. J. Med. Chem. 2000, 43, 3624–3631. [Google Scholar] [CrossRef] [PubMed]
- Johansson, E.; Parkinson, G.N.; Denny, W.A.; Neidle, S. Studies on the nitroreductase prodrug-activating system. Crystal structures of complexes with the inhibitor dicoumarol and dinitrobenzamide prodrugs and of the enzyme active form. J. Med. Chem. 2003, 46, 4009–4020. [Google Scholar] [CrossRef]
- Kobori, T.; Sasaki, H.; Lee, W.C.; Zenno, S.; Saigo, K.; Murphy, M.E.; Tanokura, M. Structure and site-directed mutagenesis of a flavoprotein from Escherichia coli that reduces nitrocompounds: Alteration of pyridine dinucleotidenbinding by a single amino acid substitution. J. Biol. Chem. 2001, 276, 2816–2823. [Google Scholar] [CrossRef] [PubMed]
- Haynes, C.A.; Koder, R.L.; Miller, A.-F.; Rodgers, D.W. Structure of nitroreductase in three states: Effects of inhibitor binding and reduction. J. Biol. Chem. 2002, 277, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Koder, R.L.; Haynes, C.A.; Rodgers, M.E.; Rodgers, D.W.; Miller, A.-F. Flavin thermodynamics explain the thermodynamics of enteric nitroreductases. Biochemistry 2002, 48, 14197–14205. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, G.; Chen, S.; Massey, V. DT-diaphorase. Redox potential, steady-state, and rapid reaction studies. J. Biol. Chem. 1995, 270, 1198–1204. [Google Scholar] [CrossRef] [PubMed]
- Hay, M.P.; Anderson, R.F.; Ferry, D.M.; Wilson, W.R.; Denny, W.A. Synthesis and evaluation of nitroheterocyclic carbamate prodrugs for use with nitroreductase-mediated gene-directed enzyme prodrug therapy. J. Med. Chem. 2003, 46, 5533–5545. [Google Scholar] [CrossRef]
- Mowday, A.M.; Ashoorzadeh, A.; Williams, E.M.; Copp, J.N.; Abbatista, M.R.; Anderson, R.F.; Flanagan, J.U.; Guise, C.P.; Ackerley, D.F.; Smaill, J.B.; et al. Rational design of an ACR1C3-resistant analog of PR-104 for enzyme-prodrug therapy. Biochem. Pharmacol. 2016, 116, 176–187. [Google Scholar] [CrossRef]
- Copp, J.N.; Mowday, A.M.; Williams, E.M.; Guise, C.P.; Ashoorzadeh, A.; Sharrock, A.V.; Flanagan, J.U.; Smaill, J.B.; Patterson, A.V.; Ackerley, D.F. Engineering a multifunctional nitroreductase for improved activation of prodrugs and PET probes for cancer gene therapy. Cell Chem. Biol. 2017, 24, 391–403. [Google Scholar] [CrossRef]
- Mowday, A.M.; Copp, J.N.; Syddal, S.P.; Dubois, L.J.; Wang, J.; Lieuwes, N.G.; Biermans, R.; Ashoorzadeh, A.; Abbatista, M.R.; Williams, E.M.; et al. E. coli nitroreductase NfsA is a reporter gene for non-invasive PET imaging in cancer gene therapy applications. Theranostics 2020, 10, 10548–10562. [Google Scholar] [CrossRef] [PubMed]
- Vass, S.O.; Jarrom, D.; Wilson, W.R.; Hyde, E.I.; Searle, P.F. E. coli NfsA: An alternative nitroreductase for prodrug activation gene therapy in combination with CB1954. Br. J. Cancer 2009, 100, 1903–1911. [Google Scholar] [CrossRef] [PubMed]
- Wardman, P.; Dennis, M.F.; Everett, S.A.; Patel, K.B.; Stratford, M.R.; Tracy, M. Radicals from one-electron reduction of nitro compounds, aromatic N-oxides and quinones: The kinetic basis for hypoxia-selective, bioreductive drugs. Biochem. Soc. Symp. 1995, 61, 171–194. [Google Scholar]
- Cheng, Z.; Arscott, L.D.; Ballou, D.P.; Williams, C.H., Jr. The relationship of the redox potentials of thioredoxin and thioredoxin reductase from Drosophila melanogaster to the enzymatic mechanisms: Reduced thioredoxin is the reductant of glutathione in Drosophila. Biochemistry 2007, 46, 7875–7885. [Google Scholar] [CrossRef]
- Lesanavičius, M.; Seo, D.; Čėnas, N. Thioredoxin reductase-type ferredoxin:NADP+ oxidoreductase of Rhodopseudomonas palustris: Potentiometric characteristics and reactions with nonphysiological oxidants. Antioxidants 2022, 11, 1000. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, N.O.; Ciotti, M.M. Chemistry and properties of the 3-acetylpyridine analogue of diphosphopyridine dinucleotide. J. Biol. Chem. 1956, 221, 823–832. [Google Scholar] [CrossRef]
- Chan-Hyams, J.V.E. Characterisation and Optimisation of Nitroreductase-Prodrug Combinations for Bacterial-Directed Enzyme-Prodrug Therapy. Ph.D. Thesis, Victoria University of Wellington, Wellington, New Zealand, 2020. [Google Scholar]
- Hall, K.R.; Robins, K.J.; Williams, E.M.; Rich, M.H.; Calcott, M.J.; Copp, J.N.; Little, R.F.; Schwörer, R.; Evans, G.B.; Patrick, W.M.; et al. Intracellular complexities of acquiring a new enzymatic function revealed by a mass-randomisation of active site residues. eLife 2020, 9, e59081. [Google Scholar] [CrossRef]
- Čėnas, N.; Anusevičius, Ž.; Nivinskas, H.; Misevičienė, L.; Šarlauskas, J. Structure-activity relationships in two-electron reduction of quinones. Methods Enzymol. 2004, 382, 258–277. [Google Scholar]
- Wardman, P. Reduction potentials of one-electron couples involving free radicals in aqueous solutions. J. Phys. Chem. Ref. Data 1989, 18, 1637–1755. [Google Scholar] [CrossRef]
- Van der Wiel, A.M.A.; Jackson-Patel, V.; Niemans, R.; Yaromina, A.; Liu, E.; Marcus, D.; Mowday, A.M.; Lieuwes, N.G.; Biemans, R.; Lin, X.; et al. Selectively targeting tumor hypoxia with the hypoxia-activated prodrug CP-506. Mol. Cancer Ther. 2021, 20, 2372–2383. [Google Scholar] [CrossRef]
- Williams, E.M. Development of Bacterial Nitroreductase Enzymes for Noninvasive Imaging in Cancer Cell Therapy. Ph.D. Thesis, Victoria University of Wellington, Wellington, New Zealand, 2013. [Google Scholar]
- Guha, S.N.; Moorthy, P.N.; Kishore, K.; Naik, D.B.; Rao, K.N. One-electron reduction of thionine studied by pulse radiolysis. Proc. Indian Acad. Sci. 1987, 99, 261–271. [Google Scholar] [CrossRef]
- Kubota, L.T.; Gorton, L. Electrochemical study of flavins, phenazines, phenoxazines and phenothiazines immobilized on zirconium phosphate. Electroanalysis 1999, 11, 719–728. [Google Scholar] [CrossRef]
- Guha, S.N.; Mittal, J.P. Pulse radiolysis study of one-electron reduction of safranine T. J. Chem. Soc. Faraday Trans. 1997, 93, 3647–3652. [Google Scholar] [CrossRef]
- Sharrock, A.V.; Mumm, J.S.; Bagdžiūnas, G.; Čėnas, N.; Arcus, V.L.; Ackerley, D.F. The crystal structure of engineered nitroreductase NTR 2.0 and impact of F70A and F108Y substitutions on substrate specificity. Int. J. Molec. Sci. 2023, 24, 6633. [Google Scholar] [CrossRef]
- Lei, B.; Wang, H.; Yu, Y.; Tu, S.-C. Redox potential and equilibrium in the reductive half-reaction of Vibrio harveyi NADPH-FMN oxidoreductase. Biochemistry 2005, 44, 261–267. [Google Scholar] [CrossRef]
- Musila, J.M.; Rokita, S.E. Sequence conservation does not always signify a functional imperative as observed in the nitroreductase family. Biochemistry 2022, 61, 703–711. [Google Scholar] [CrossRef]
- Günger, T.; Önder, F.C.; Tokay, E.; Gülhan, Ű.G.; Hacioğlu, N.; Tok, T.T.; Ҫelik, A.; Köckar, F.; Ay, M. Prodrugs for nitroreductase based cancer therapy -2: Novel amide/Ntr combination targeting PC3 cancer cells. Eur. J. Med. Chem. 2019, 171, 383–400. [Google Scholar] [CrossRef]
- Hall, B.S.; Meredith, E.L.; Wilkinson, S.R. Targeting the substrate preference of a type I nitroreductase to develop antitrypanosomal quinone-based prodrugs. Antimicrob. Agents Chemother. 2012, 56, 5821–5830. [Google Scholar] [CrossRef]
- Leelakriangsak, M.; Huyen, N.T.T.; Töve, S.; Duy, N.V.; Becher, D.; Hecker, M.; Antelmann, H.; Zuber, P. Regulation of quinone detoxification by the thiol stress sensing DUF24/MarR-like repressor, YodB in Bacillus subtilis. Mol. Microbiol. 2008, 67, 1108–1124. [Google Scholar] [CrossRef]
- Rau, J.; Stolz, A. Oxygen insensitive nitroreductases NfsA and NfsB of Escherichia coli function under anaerobic conditions as lawsone-dependent azoreductases. Appl. Environ. Microbiol. 2003, 69, 3448–3455. [Google Scholar] [CrossRef]
- Chalansonnet, V.; Mercier, C.; Orenga, S.; Gilbert, C. Identification of Enterococcus faecalis enzymes with azoreductases and/or nitroreductase activity. BMC Microbiol. 2017, 17, 126. [Google Scholar] [CrossRef]
- Wang, Z.; Sapienza, P.J.; Abeysinghe, T.; Luzum, C.; Lee, A.L.; Finer-Moore, J.S.; Stroud, R.M.; Kohen, A. Mg2+ binds to the surface of thymidylate synthase and affects hydride transfer at the interior active site. J. Am. Chem. Soc. 2013, 135, 7583–7592. [Google Scholar] [CrossRef]
- Fan, Y.; Cembran, A.; Ma, S.; Gao, J. Connecting protein conformational dynamics with catalytic function as illustrated in dihydrofolate reductase. Biochemistry 2013, 52, 2036–2049. [Google Scholar] [CrossRef]
- Yang, J.; Zhan, J.; Bai, J.; Liu, P.; Xue, Y.; Yang, Q. Residue Phe42 is critical for the catalytic activity of Escherichia coli major nitroreductase NfsA. Biotechnol. Lett. 2013, 35, 1693–1700. [Google Scholar] [CrossRef]
- Radveikienė, I.; Palinauskas, D.; Ragauskaitė, E.; Bagdžiūnas, G. Self-asembled cyclohextrins-based nanostructures on indium-tin-oxide for a detection of catecholamine neurotransmitters. Appl. Surf. Sci. 2022, 600, 154170. [Google Scholar] [CrossRef]
- Anlezark, G.M.; Melton, R.G.; Sherwood, R.F.; Wilson, W.R.; Denny, W.A.; Palmer, B.D.; Knox, R.J.; Friedlos, F.; Williams, A. Bioactivation of dinitrobenzamide mustards by an E. coli B nitroreductase. Biochem. Pharmacol. 1995, 50, 609–618. [Google Scholar] [CrossRef]
- Chou, F.; Chan, A.H.; Driscoll, J.S. Potential central nervous system antitumour agents. Aziridinylbenzoquinones. J. Med. Chem. 1976, 28, 1302–1308. [Google Scholar] [CrossRef]
- Khan, A.H.; Ross, W.C.J. Tumor-growth inhibitory nitrophenylaziridines and related compounds: Structure-activity relationships. Chem.-Biol. Interact. 1969, 1, 27–47. [Google Scholar] [CrossRef]
- Palmer, B.D.; Wilson, W.R.; Atwell, G.J.; Schultz, D.; Xu, X.Z.; Denny, W.A. Hypoxia-selective antitumor agents. 9. Structure-activity relationships for hypoxia-selective cytotoxicity among analogues of 5-[N,N-bis(2-chloroethyl)amino]-2,4-dinitrobenzamide. J. Med. Chem. 1994, 37, 2175–2184. [Google Scholar] [CrossRef]
- Weber, M.; Kaplan, N. Flavoenzyme-catalyzed pyridine nucleotide transfer reactions. J. Biol. Chem. 1957, 225, 909–920. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | E17 (V) | E7(H−) (V) | kcat(app.) (s−1) | kcat/Km (M−1s−1) | VdWvol (Å3) | Log D |
|---|---|---|---|---|---|---|---|
| Nitroaromatic Compounds | |||||||
| 1 | 2,4,6-Trinitrotoluene | −0.253 a | − | 60 ± 5.0 | 5.6 ± 0.31 × 106 | 183 | 2.31 |
| 2 | p-Dinitrobenzene | −0.255 | − | 71 ± 5.0 | 1.8 ± 0.10 × 106 | 142 | 1.85 |
| 3 | Nifuroxime | −0.255 | − | 152 ± 8.0 180 ± 20.0 b | 4.2 ± 0.32 × 106 5.9 ± 0.60 × 106 b | 133 | −0.34 |
| 4 | Nitrofurantoin | −0.255 | − | 119 ± 7.0 136 ± 18.0 b | 5.2 ± 0.2 × 106 7.2 ± 1.0 × 106 b | 203 | −0.25 |
| 5 | o-Dinitrobenzene | −0.287 | − | 60 ± 5.5 | 3.4 ± 0.29 × 106 | 142 | 1.85 |
| 6 | Nitracrine | −0.303 | − | − | ≤3.0 × 102 | 336 | 0.00 |
| 7 | p-Nitrobenzaldehyde | −0.325 | − | 31 ± 3.0 | 1.8 ± 0.15 × 105 | 142 | 1.63 |
| 8 | 3,5-Dinitrobenzoic acid | −0.344 | − | 64 ± 5.0 | 1.8 ± 0.10 × 106 | 170 | −1.79 |
| 9 | m-Dinitrobenzene | −0.345 | − | 48 ± 5.0 55 ± 6.0 b | 1.0 ± 0.08 × 106 1.2 ± 0.10 × 106 b | 142 | 1.85 |
| 10 | SN-36506 c | −0.351 | − | 1.5 ± 0.2 | 2.2 ± 0.6 × 103 | 464 | −3.40 |
| 11 | p-Nitroacetophenone b | −0.355 | − | 59 ± 5.0 | 1.5 ± 0.20 × 105 | 161 | 1.47 |
| 12 | PR-104A d | −0.366 | − | 8.4 ± 0.6 | 9.3 × 104 | 386 | 0.95 |
| 13 | Tizoxanide | −0.380 a | − | 24 ± 1.8 | 9.6 ± 0.82 × 104 | 226 | 2.16 |
| 14 | Benznidazole | −0.380 a | − | 24 ± 2.1 | 3.2 ± 0.24 × 105 | 251 | 1.32 |
| 15 | CB-1954 | −0.385 | − | 25 ± 2.0 | 3.5 ± 0.20 × 105 | 215 | 0.64 |
| 16 | Nitrothiophene | −0.390 | − | 9.0 ± 0.8 | 3.6 ± 0.27 × 104 | 108 | 1.86 |
| 17 | EF-5 e | −0.395 | − | 8.9 ± 0.3 | 5.6 ± 0.80 × 104 | 197 | 1.25 |
| 18 | 2-Amino-4,6-dinitrotoluene | −0.417 a | − | 61 ± 4.8 | 3.5 ± 0.18 × 105 | 170 | 1.54 |
| 19 | SN-23862 | −0.425 | − | 18 ± 1.4 | 1.2 ± 0.10 × 105 | 305 | 2.14 |
| 20 | p-Nitrobenzoic acid b | −0.425 | − | 64 ± 7.0 | 2.5 ± 0.20 × 104 | 149 | −1.66 |
| 21 | Fexinidazole | −0.458 a | − | ≥0.6 | 3.0 ± 0.25 × 103 | 265 | 2.38 |
| 22 | Tinidazole | −0.480 | − | 2.0 ± 0.3 | 5.6 ± 0.36 × 103 | 223 | −0.58 |
| 23 | Nitrobenzene b | −0.485 | − | 14 ± 2.0 | 9.6 ± 0.80 × 103 | 122 | 1.91 |
| 24 | Chloramphenicol f | −0.546 a | − | 0.89 ± 0.03 | 8.6 ± 0.90 × 102 | 270 | 1.10 |
| Quinones | |||||||
| 25 | 1,4-Benzoquinone b | 0.090 | 0.195 | 62 ± 6.0 | 5.8 ± 0.60 × 106 | 109 | 1.02 |
| 26 | 2-Methyl-1,4-benzoquinone b | 0.010 | 0.120 | 54 ± 6.0 | 6.8 ± 0.70 × 106 | 128 | 1.42 |
| 27 | DZQ | −0.054 | 0.000 | 38 ± 3.0 | 9.2 ± 0.50 × 104 | 194 | 0.0 |
| 28 | 2,6-Dimethyl-1,4-benzoquinone b | −0.080 | 0.058 | 51 ± 6.0 | 3.2 ± 0.40 × 106 | 146 | 1.82 |
| 29 | 5-Hydroxy-1,4-naphthoquinone b | −0.090 | −0.060 | 22 ± 3.0 | 1.4 ± 0.20 × 107 | 166 | 1.38 |
| 30 | 5,8-Dihydroxy-1,4-naphthoquinone b | −0.110 | −0.084 | 12 ± 2.0 | 1.3 ± 0.10 × 107 | 171 | 2.19 |
| 31 | 9,10-Phenanthrene quinone b | −0.120 | −0.034 | 54 ± 6.0 | 4.7 ± 0.50 × 106 | 212 | 2.92 |
| 32 | 1,4-Naphthoquinone b | −0.150 | −0.029 | 46 ± 5.0 | 1.4 ± 0.20 × 106 | 161 | 1.49 |
| 33 | 2-Methyl-1,4-naphthoquinone b | −0.200 | −0.114 | 36 ± 4.0 | 2.8 ± 0.30 × 105 | 179 | 1.89 |
| 34 | MeDZQ | −0.230 | −0.128 | 2.9 ± 0.2 | 1.3 ± 0.10 × 104 | 229 | 0.79 |
| 35 | 9,10-Anthraquinone-2,6-disulfonate | −0.250 | −0.210 | ≥0.7 | 6.2 ± 0.35 × 103 | 287 | −3.47 |
| 36 | Tetramethyl-1,4-benzoquinone b | −0.260 | −0.086 | 2.5 ± 0.3 | 2.3 ± 0.20 × 103 | 181 | 2.61 |
| 37 | 1,8-Dihydroxy-9,10-anthraquinone b | −0.325 | − | 0.7 ± 0.1 | 1.0 ± 0.15 × 105 | 224 | 3.61 |
| 38 | 9,10-Anthraquinone-2-sulfonate b | −0.380 | −0.250 | 1.4 ± 0.1 | 1.9 ± 0.20 × 104 | 246 | −0.27 |
| 39 | 2-Hydroxy-1,4-naphthoquinone b | −0.410 | −0.200 | 72 ± 8.0 | 1.1 ± 0.10 × 107 | 166 | −0.99 |
| 40 | 2-Methyl-3-hydroxy-1,4-naphthoquinone b | −0.460 | −0.240 | 56 ± 5.0 | 8.8 ± 1.50 × 106 | 184 | −0.69 |
| Miscellaneous | |||||||
| 41 | Thionine | 0.055 ÷ −0.045 g | 0.064 h | 56 ± 5.0 | 6.9 ± 0.50 × 104 | 219 | 0.8 |
| 42 | Riboflavin b | −0.320 | −0.210 | 4.2 ± 0.6 | 2.6 ± 0.20 × 105 | 348 | −1.18 |
| 43 | Safranine-T | −0.350 i | −0.286 h | ≥0.3 | 2.5 ± 0.21 × 103 | 333 | n.d. |
| 44 | Ferricyanide j | 0.410 | 89 ± 6.0 | 8.6 ± 1.40 × 106 | n.d. | n.d. | |
| Oxidant | ΔH≠ (kJ mol−1) | ΔS≠ (J mol−1 K−1) | −TΔS≠ (kJ mol−1) |
|---|---|---|---|
| Nitroaromatic Compounds | |||
| 2,4,6-Trinitrotoluene | 24.2 ± 1.7 | −34.2 ± 5.7 | 10.2 ± 1.7 |
| p-Nitrobenzaldehyde | 27.2 ± 0.9 | −55.2 ± 3.1 | 16.4 ± 0.9 |
| o-Dinitrobenzene | 23.5 ± 1.4 | −33.6 ± 5.2 | 10.0 ± 1.5 |
| Quinones | |||
| 2-Methyl-3-Hydroxy-1,4-naphthoquinone | 38.7 ± 1.1 | 9.7 ± 3.7 | -2.9 ± 1.1 |
| 2-Hydroxy-1,4-naphthoquinone | 28.3 ± 2.1 | −16.0 ± 7.0 | 4.8 ± 2.1 |
| 2-Methyl-1,4-naphthoquinone | 23.4 ± 1.8 | −54.4 ± 6.0 | 16.2 ± 1.8 |
| Tetramethyl-1,4-benzoquinone | 16.4 ± 1.4 | −123.0 ± 4.5 | 36.7 ± 1.3 |
| Compound | H Bonds | Shortest Plane-to-Plane Distance to Isoalloxazine (Å) | Distance to N5 of Isoalloxazine (Å) |
|---|---|---|---|
| Nitroaromatic Compounds | |||
| CB-1954 | 2-NO2…Arg133, 4-NO2…Arg15, -CONH2…Arg133, Gln67 | 3.7 | 2-NO2: 2.9 (O), 3.9 (N) |
| PR-104A | -C2H4OH…Arg15, 6-NO2…H2O, -OSO2CH3…H2O | 3.8 | 4-NO2: 3.8 (O), 3.4 (N) |
| 2,4,6-Trinitrotoluene | 2-NO2…Arg15, 4-NO2…Arg133, 6-NO2…Ser40. | 4.0 | 2-NO2: 3.0 (O), 3.4 (N) |
| o-Dinitrobenzene | 1-NO2…Arg15, 2-NO2…H2O | 3.9 | 1-NO2: 3.0 (O), 3.9 (N) |
| p-Nitrobenzaldehyde | -NO2…Arg15, -CHO…Arg133 | 4.6 | 3.9 (O), 4.6 (N) |
| Nitracrine | -NO2…Arg15, -N(CH3)2…Ser40 | 7.2 | 8.1 (O), 9.0 (N) |
| Quinones and Heteroaromatics | |||
| 2-Hydroxy-1,4-naphthoquinone | 1-C=O…Arg15, 4-C=O…Ser40, 2-O− …H2O | 3.8 | 4-C=O: 3.0 (O), 3.5 (C) |
| 2-Methyl-3-hydroxy-1,4-naphthoquinone | 1-C=O…Ser40, 4-C=O…Arg15, 3-O−…H2O | 4.2 | 1-C=O: 3.1 (O), 3.6 (C) |
| 2-Methyl-1,4-naphthoquinone | 1-C=O…Arg133, 4-C=O…Arg15 | 3.3 | 1-C=O: 5.1 (O), 4.9 (C) |
| Tetramethyl-1,4-benzoquinone | 1-C=O…Arg133, 4-C=O…Arg15 | 3.3 | 1-C=O: 5.1 (O), 4.7 (C) |
| DZQ | 1-C=O…Arg133, 4-C=O…Arg15 | 3.3 | 1-C=O: 4.1 (O), 4.1 (C) |
| 9,10-Anthraquinone-2,6-disulfonate | 2-SO3−…Arg15, 6-SO3−…Arg133 | 4.5 | 10-C=O: 6.0 (O), 6.3 (C) |
| Thionine | 3(7)-NH2…Arg15 | 3.6 | 5.4 (S), 5.3 (N10) |
| Safranine-T | 3(7)-NH2…Arg15 | 4.5 | 5.3 (N5(10)) |
| Enzyme | Standard Redox Potential | Conditions |
|---|---|---|
| E. cloacae NR (B) [27] | −0.190 V | Equilibration with redox mediator, PIPES, KCl, pH 7.0 |
| Vibrio fischeri NAD(P)H: FMN oxidoreductase (B) [48] | −0.215 V | Equilibration with redox mediator, phosphate, pH 7.0 |
| E. coli NfsB [49] | −0.199 V | Equilibration with redox mediator, Tris, NaCl, pH 7.0 |
| E. coli NfsB [22] | −0.218 V | Direct electrochemical reduction, phosphate, KCl, 10% glycerol, pH 7.5 |
| Vibrio harveyi NADPH: FMN oxidoreductase (A) [48] | −0.255 V | Equilibration with redox mediator, phosphate, pH 7.0 |
| E. coli NfsA [21] | −0.264 V FMNox/sq: −0.268 V FMNsq/red: −0.272 V a | Direct electrochemical reduction, phosphate, KCl, 10% glycerol, pH 7.5 |
| E. coli NfsA, this work | −0.215 V | Reactions with APADP+/APADPH, phosphate, pH 7.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valiauga, B.; Bagdžiūnas, G.; Sharrock, A.V.; Ackerley, D.F.; Čėnas, N. The Catalysis Mechanism of E. coli Nitroreductase A, a Candidate for Gene-Directed Prodrug Therapy: Potentiometric and Substrate Specificity Studies. Int. J. Mol. Sci. 2024, 25, 4413. https://doi.org/10.3390/ijms25084413
Valiauga B, Bagdžiūnas G, Sharrock AV, Ackerley DF, Čėnas N. The Catalysis Mechanism of E. coli Nitroreductase A, a Candidate for Gene-Directed Prodrug Therapy: Potentiometric and Substrate Specificity Studies. International Journal of Molecular Sciences. 2024; 25(8):4413. https://doi.org/10.3390/ijms25084413
Chicago/Turabian StyleValiauga, Benjaminas, Gintautas Bagdžiūnas, Abigail V. Sharrock, David F. Ackerley, and Narimantas Čėnas. 2024. "The Catalysis Mechanism of E. coli Nitroreductase A, a Candidate for Gene-Directed Prodrug Therapy: Potentiometric and Substrate Specificity Studies" International Journal of Molecular Sciences 25, no. 8: 4413. https://doi.org/10.3390/ijms25084413