
Plasma Metabolite Profiling in the Search for Early-Stage Biomarkers for Lung Cancer: Some Important Breakthroughs

 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Evolution in NMR Metabolomics Research—Part I: Feasibility of Lung Cancer Detection via Blood Plasma Using NMR Spectroscopy
2.1. Feasibility: Lung Cancer Detection
2.1.1. Study Set-Up
2.1.2. Results
2.1.3. Supporting Evidence
2.2. Feasibility: Differentiation between Cancer Types: Lung Cancer versus Breast Cancer
2.2.1. Study Set-Up
2.2.2. Results
3. Evolution in NMR Metabolomics Research—Part II: Developments in Preanalytical Sample Preparation and NMR Measurement Procedure
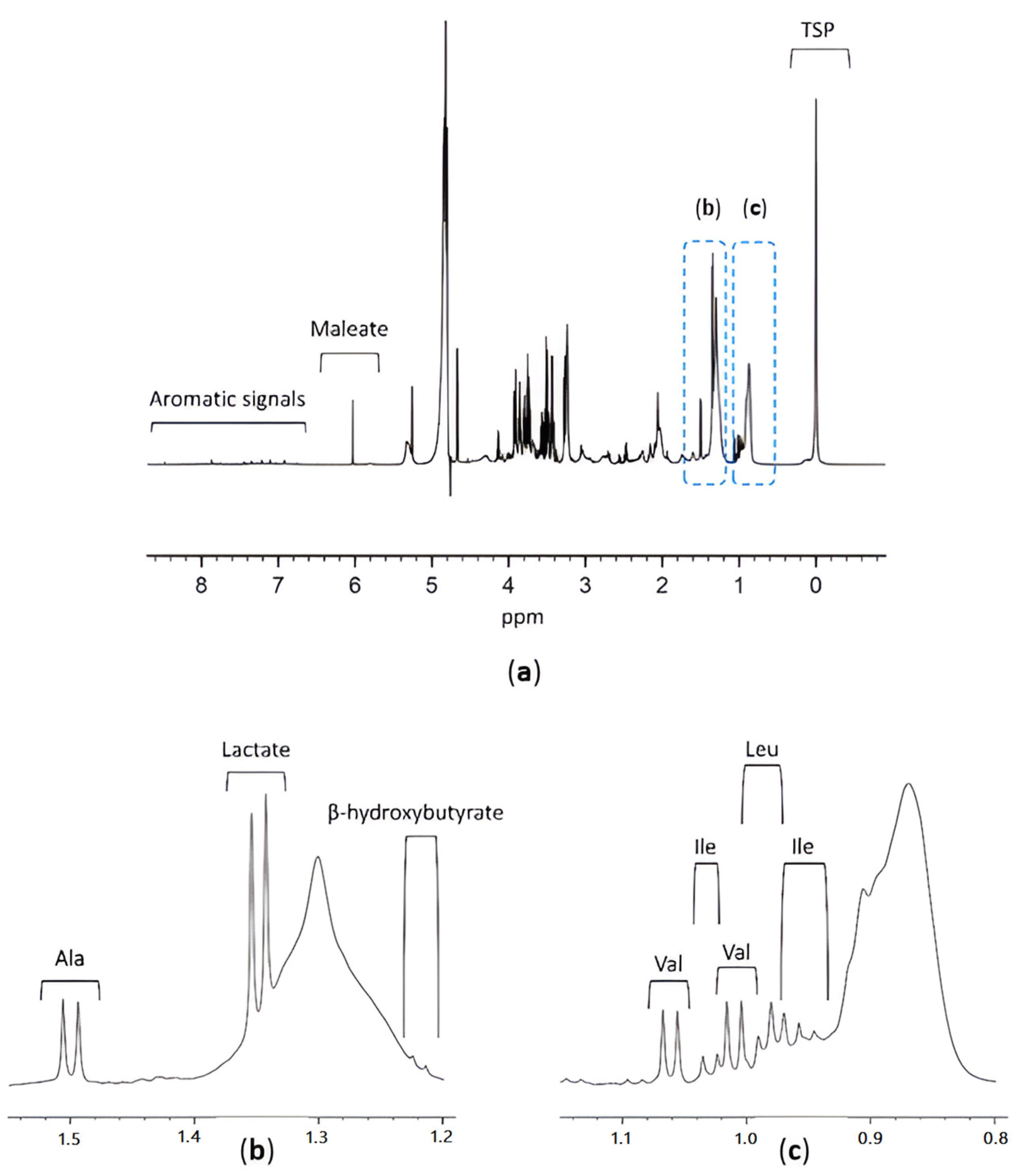
3.1. TSP as an HSA Binding Competitor and MA as an Internal Standard
3.2. Study Design
3.3. Results
4. Evolution in NMR Metabolomics Research—Part III: Plasma Biomarkers for Early-Stage NSCLC and Their Potential for Detection and Monitoring of NSCLC Recurrence
4.1. Study Design
4.2. Results
5. Discussion of Breakthroughs in NMR Metabolomics Research
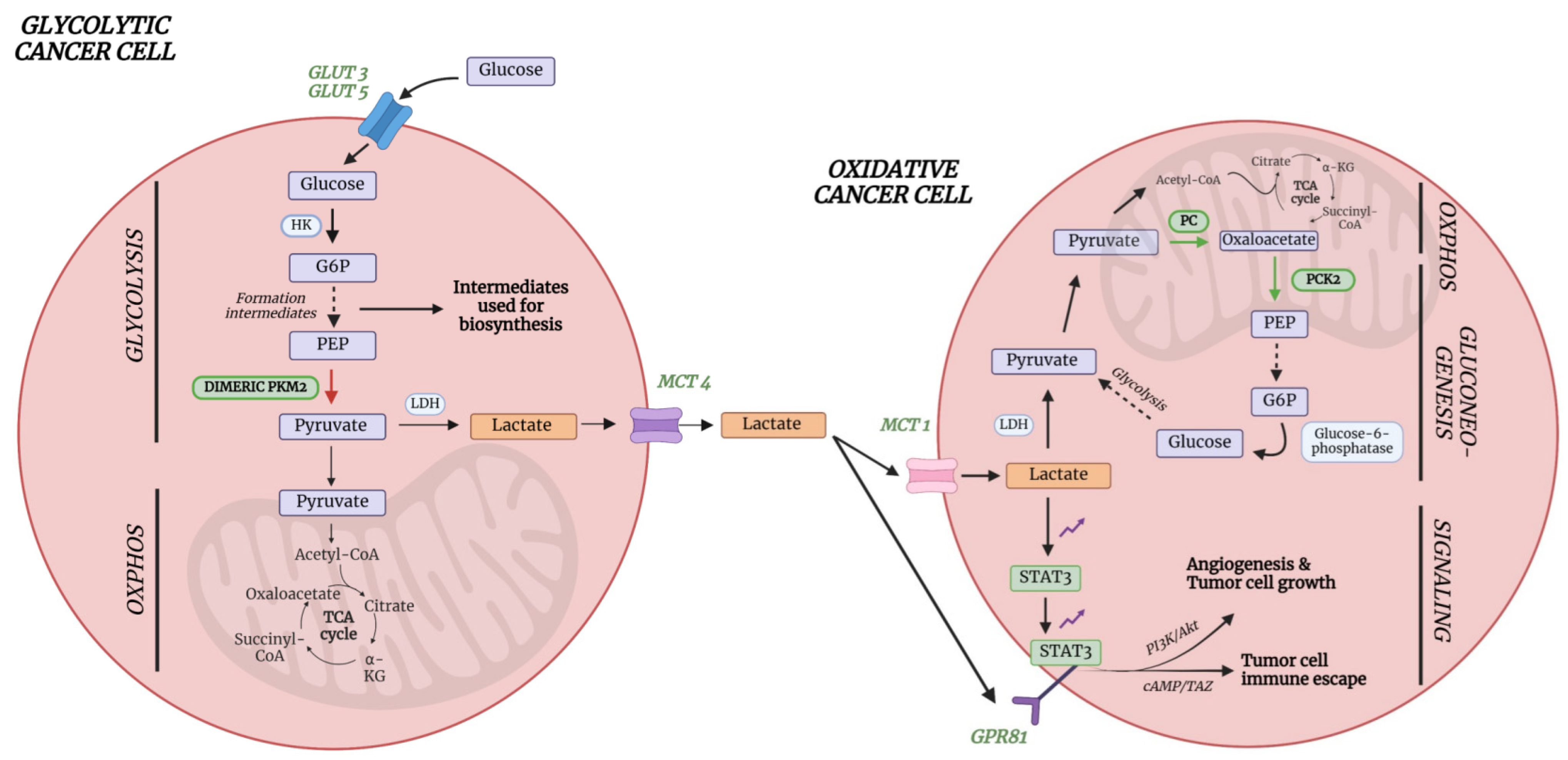
6. Metabolic Pathways Involved in (Lung) Cancer
6.1. Lactate
6.2. Acetate and Amino Acids Cysteine and Asparagine
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B
Appendix B.1. Sample Collection
Appendix B.2. Sample Preparation
Appendix B.3. Metabolite Spiking
Appendix B.4. NMR Data Acquisition
Appendix B.5. NMR Spectra Processing

References
- Hirsch, F.R.; Scagliotti, G.V.; Mulshine, J.L.; Kwon, R.; Curran, W.J.; Wu, Y.-L.; Paz-Ares, L. Lung cancer: Current therapies and new targeted treatments. Lancet 2017, 389, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.E.; Kazerooni, E.A.; Baum, S.L.; Eapen, G.A.; Ettinger, D.S.; Hou, L.; Jackman, D.M.; Klippenstein, D.; Kumar, R.; Lackner, R.P.; et al. Lung Cancer Screening, Version 3.2018, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2018, 16, 412–441. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, H.; Heuvelmans, M.A.; Oudkerk, M. Low-dose computed tomography lung cancer screening: Clinical evidence and implementation research. J. Intern. Med. 2022, 292, 68–80. [Google Scholar] [CrossRef] [PubMed]
- The National Lung Screening Trial Research Team. Reduced lung-cancer mortality with low-dose computed tomographic screening. N. Engl. J. Med. 2011, 365, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S.W.; Field, J. Mortality Reduction with Low-Dose CT Screening for Lung Cancer. N. Engl. J. Med. 2020, 382, 572–573. [Google Scholar] [CrossRef] [PubMed]
- Oudkerk, M.; Liu, S.; Heuvelmans, M.A.; Walter, J.E.; Field, J.K. Lung cancer LDCT screening and mortality reduction—Evidence, pitfalls and future perspectives. Nat. Rev. Clin. Oncol. 2021, 18, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Berg, C.D.; Aberle, D.R.; Wood, D.E. Lung Cancer Screening: Promise and Pitfalls. Am. Soc. Clin. Oncol. Educ. Book 2012, 32, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Farwell, M.D.; Pryma Da Fau-Mankoff, D.A.; Mankoff, D.A. PET/CT imaging in cancer: Current applications and future directions. Cancer 2014, 120, 3433–3445. [Google Scholar] [CrossRef] [PubMed]
- Deppen, S.A.; Blume, J.D.; Kensinger, C.D.; Morgan, A.M.; Aldrich, M.C.; Massion, P.P.; Walker, R.C.; McPheeters, M.L.; Putnam, J.B., Jr.; Grogan, E.L. Accuracy of FDG-PET to diagnose lung cancer in areas with infectious lung disease: A meta-analysis. JAMA 2014, 312, 1227–1236. [Google Scholar] [CrossRef]
- Rosenbaum, S.J.; Lind, T.; Antoch, G.; Bockisch, A. False-Positive FDG PET Uptake−the Role of PET/CT. Eur. Radiol. 2006, 16, 1054–1065. [Google Scholar] [CrossRef]
- Steven, B.Z.; Elizabeth, T.L.; Christopher, G.S.; David, H.A.; Paul, L.H.; Gregory, J.K.; Larry, G.K.; Leah, M.B. Preoperative PET and the Reduction of Unnecessary Surgery Among Newly Diagnosed Lung Cancer Patients in a Community Setting. J. Nucl. Med. 2014, 55, 379. [Google Scholar] [CrossRef]
- Vanhove, K.; Giesen, P.; Owokotomo, O.E.; Mesotten, L.; Louis, E.; Shkedy, Z.; Thomeer, M.; Adriaensens, P. The plasma glutamate concentration as a complementary tool to differentiate benign PET-positive lung lesions from lung cancer. BMC Cancer 2018, 18, 868. [Google Scholar] [CrossRef] [PubMed]
- Seijo, L.M.; Peled, N.; Ajona, D.; Boeri, M.; Field, J.K.; Sozzi, G.; Pio, R.; Zulueta, J.J.; Spira, A.; Massion, P.P.; et al. Biomarkers in Lung Cancer Screening: Achievements, Promises, and Challenges. J. Thorac. Oncol. 2019, 14, 343–357. [Google Scholar] [CrossRef] [PubMed]
- Clish, C.B. Metabolomics: An emerging but powerful tool for precision medicine. Cold Spring Harb. Mol. Case Stud. 2015, 1, a000588. [Google Scholar] [CrossRef]
- Vanhove, K.; Derveaux, E.; Mesotten, L.; Thomeer, M.; Criel, M.; Mariën, H.; Adriaensens, P. Unraveling the Rewired Metabolism in Lung Cancer Using Quantitative NMR Metabolomics. Int. J. Mol. Sci. 2022, 23, 5602. [Google Scholar] [CrossRef]
- Noreldeen, H.A.A.; Liu, X.; Xu, G. Metabolomics of lung cancer: Analytical platforms and their applications. J. Sep. Sci. 2020, 43, 120–133. [Google Scholar] [CrossRef]
- Pascale, R.; Calvisi, D.F.; Simile, M.M.; Feo, C.; Feo, F. The Warburg Effect 97 Years after Its Discovery. Cancers 2020, 12, 2819. [Google Scholar] [CrossRef] [PubMed]
- Louis, E.; Adriaensens, P.; Guedens, W.; Bigirumurame, T.; Baeten, K.; Vanhove, K.; Vandeurzen, K.; Darquennes, K.; Vansteenkiste, J.; Dooms, C.; et al. Detection of Lung Cancer through Metabolic Changes Measured in Blood Plasma. J. Thorac. Oncol. 2016, 11, 516–523. [Google Scholar] [CrossRef]
- Louis, E.; Adriaensens, P.; Guedens, W.; Vanhove, K.; Vandeurzen, K.; Darquennes, K.; Vansteenkiste, J.; Dooms, C.; de Jonge, E.; Thomeer, M.; et al. Metabolic phenotyping of human blood plasma: A powerful tool to discriminate between cancer types? Ann. Oncol. 2016, 27, 178–184. [Google Scholar] [CrossRef]
- Derveaux, E.; Thomeer, M.; Mesotten, L.; Reekmans, G.; Adriaensens, P. Detection of Lung Cancer via Blood Plasma and (1)H-NMR Metabolomics: Validation by a Semi-Targeted and Quantitative Approach Using a Protein-Binding Competitor. Metabolites 2021, 11, 537. [Google Scholar] [CrossRef]
- Derveaux, E.; Geubbelmans, M.; Criel, M.; Demedts, I.; Himpe, U.; Tournoy, K.; Vercauter, P.; Johansson, E.; Valkenborg, D.; Vanhove, K.; et al. NMR-Metabolomics Reveals a Metabolic Shift after Surgical Resection of Non-Small Cell Lung Cancer. Cancers 2023, 15, 2127. [Google Scholar] [CrossRef] [PubMed]
- Worley, B.; Powers, R. Multivariate Analysis in Metabolomics. Curr. Metabolomics 2013, 1, 92–107. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, L.; Byrne, T.; Johansson, E.; Trygg, J.; Vikström, C. Multi- and Megavariate Data Analysis Basic Principles and Applications; Umetrics Academy: Umeå, Sweden, 2013. [Google Scholar]
- Puchades-Carrasco, L.; Jantus-Lewintre, E.; Pérez-Rambla, C.; García-García, F.; Lucas, R.; Calabuig, S.; Blasco, A.; Dopazo, J.; Camps, C.; Pineda-Lucena, A. Serum metabolomic profiling facilitates the non-invasive identification of metabolic biomarkers associated with the onset and progression of non-small cell lung cancer. Oncotarget 2016, 7, 12904–12916. [Google Scholar] [CrossRef] [PubMed]
- Rocha, C.M.; Carrola, J.; Barros, A.S.; Gil, A.M.; Goodfellow, B.J.; Carreira, I.M.; Bernardo, J.; Gomes, A.; Sousa, V.; Carvalho, L.; et al. Metabolic signatures of lung cancer in biofluids: NMR-based metabonomics of blood plasma. J. Proteome Res. 2011, 10, 4314–4324. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Goodacre, R. On Splitting Training and Validation Set: A Comparative Study of Cross-Validation, Bootstrap and Systematic Sampling for Estimating the Generalization Performance of Supervised Learning. J. Anal. Test. 2018, 2, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Liu, J.; Lu, R. Subsampling bias and the best-discrepancy systematic cross validation. Sci. China Math. 2021, 64, 197–210. [Google Scholar] [CrossRef]
- Carrola, J.; Rocha, C.M.; Barros, A.S.; Gil, A.M.; Goodfellow, B.J.; Carreira, I.M.; Bernardo, J.; Gomes, A.; Sousa, V.; Carvalho, L.; et al. Metabolic signatures of lung cancer in biofluids: NMR-based metabonomics of urine. J. Proteome Res. 2011, 10, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Louis, R.; Louis, E.; Stinkens, K.; Mesotten, L.; de Jonge, E.; Thomeer, M.; Caenepeel, P.; Adriaensens, P. Metabolic Phenotyping of Blood Plasma by Proton Nuclear Magnetic Resonance to Discriminate between Colorectal Cancer, Breast Cancer and Lung cancer. Metabolomics 2016, 6, 187. [Google Scholar] [CrossRef]
- Pearce, J.T.M.; Athersuch, T.J.; Ebbels, T.M.D.; Lindon, J.C.; Nicholson, J.K.; Keun, H.C. Robust algorithms for automated chemical shift calibration of 1D 1H NMR spectra of blood serum. Anal. Chem. 2008, 80, 7158–7162. [Google Scholar] [CrossRef]
- Jonsson, P.; Wuolikainen, A.; Thysell, E.; Chorell, E.; Stattin, P.; Wikström, P.; Antti, H. Constrained randomization and multivariate effect projections improve information extraction and biomarker pattern discovery in metabolomics studies involving dependent samples. Metabolomics 2015, 11, 1667–1678. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, X.; Wang, C.; Zhang, H.; Cai, Z. Non-targeted and targeted metabolomics approaches to diagnosing lung cancer and predicting patient prognosis. Oncotarget 2016, 7, 63437–63448. [Google Scholar] [CrossRef] [PubMed]
- Larkin, J.; Anthony, S.; Johanssen, V.; Yeo, T.; Sealey, M.; Yates, A.G.; Smith, C.; Claridge, T.; Nicholson, B.D.; Moreland, J.; et al. Metabolomic Biomarkers in Blood Samples Identify Cancers in a Mixed Population of Patients with Nonspecific Symptoms. Clin. Cancer Res. 2022, 28, 1651–1661. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Lis, C.G. Pretreatment serum albumin as a predictor of cancer survival: A systematic review of the epidemiological literature. Nutr. J. 2010, 9, 69. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, T.; Kisluk, J.; Pietrowska, K.; Godzien, J.; Kozlowski, M.; Reszeć, J.; Sierko, E.; Naumnik, W.; Mróz, R.; Moniuszko, M.; et al. The Ability of Metabolomics to Discriminate Non-Small-Cell Lung Cancer Subtypes Depends on the Stage of the Disease and the Type of Material Studied. Cancers 2021, 13, 3314. [Google Scholar] [CrossRef] [PubMed]
- Ramteke, P.; Deb, A.; Shepal, V.; Bhat, M.K. Hyperglycemia Associated Metabolic and Molecular Alterations in Cancer Risk, Progression, Treatment, and Mortality. Cancers 2019, 11, 1402. [Google Scholar] [CrossRef] [PubMed]
- Kannampuzha, S.; Mukherjee, A.; Wanjari, U.; Gopalakrishnan, A.; Murali, R.; Namachivayam, A.; Renu, K.; Dey, A.; Vellingiri, B.; Madhyastha, H.; et al. A Systematic Role of Metabolomics, Metabolic Pathways, and Chemical Metabolism in Lung Cancer. Vaccines 2023, 11, 381. [Google Scholar] [CrossRef] [PubMed]
- Vanhove, K.; Graulus, G.J.; Mesotten, L.; Thomeer, M.; Derveaux, E.; Noben, J.P.; Guedens, W.; Adriaensens, P. The Metabolic Landscape of Lung Cancer: New Insights in a Disturbed Glucose Metabolism. Front. Oncol. 2019, 9, 1215. [Google Scholar] [CrossRef] [PubMed]
- Leithner, K.; Hrzenjak, A.; Trötzmüller, M.; Moustafa, T.; Köfeler, H.C.; Wohlkoenig, C.; Stacher, E.; Lindenmann, J.; Harris, A.L.; Olschewski, A.; et al. PCK2 activation mediates an adaptive response to glucose depletion in lung cancer. Oncogene 2015, 34, 1044–1050. [Google Scholar] [CrossRef] [PubMed]
- Mendes, C.; Serpa, J. Revisiting lactate dynamics in cancer—A metabolic expertise or an alternative attempt to survive? J. Mol. Med. 2020, 98, 1397–1414. [Google Scholar] [CrossRef]
- Wang, X.; Liu, H.; Ni, Y.; Shen, P.; Han, X. Lactate shuttle: From substance exchange to regulatory mechanism. Human Cell 2022, 35, 1–14. [Google Scholar] [CrossRef]
- Smolle, E.; Leko, P.; Stacher-Priehse, E.; Brcic, L.; El-Heliebi, A.; Hofmann, L.; Quehenberger, F.; Hrzenjak, A.; Popper, H.H.; Olschewski, H.; et al. Distribution and prognostic significance of gluconeogenesis and glycolysis in lung cancer. Cancers 2022, 14, 5344. [Google Scholar] [CrossRef] [PubMed]
- Kiesel, V.A.; Sheeley, M.P.; Coleman, M.F.; Cotul, E.K.; Donkin, S.S.; Hursting, S.D.; Wendt, M.K.; Teegarden, D. Pyruvate carboxylase and cancer progression. Cancer Metab. 2021, 9, 20. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Cui, J. Targeting the lactic acid metabolic pathway for antitumor therapy. Mol. Ther. Oncolytics 2023, 31, 100740. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Zhu, Z.; He, Y.; Zhang, Z.; Zhang, Y.; Wang, Y.; Luo, J.; Peng, T.; Cheng, F.; Gao, J.; et al. A lactate-induced Snail/STAT3 pathway drives GPR81 expression in lung cancer cells. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165576. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Yang, H.; Zhang, Y.; Wei, H.; Zhu, Z.; Zhu, B.; Yang, M.; Cao, W.; Wang, L.; Wu, Z. Tumor cell-derived lactate induces TAZ-dependent upregulation of PD-L1 through GPR81 in human lung cancer cells. Oncogene 2017, 36, 5829–5839. [Google Scholar] [CrossRef] [PubMed]
- Schug, Z.T.; Vande Voorde, J.; Gottlieb, E. The metabolic fate of acetate in cancer. Nat. Rev. Cancer 2016, 16, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Comerford, S.A.; Huang, Z.; Du, X.; Wang, Y.; Cai, L.; Witkiewicz, A.K.; Walters, H.; Tantawy, M.N.; Fu, A.; Manning, H.C.; et al. Acetate dependence of tumors. Cell 2014, 159, 1591–1602. [Google Scholar] [CrossRef] [PubMed]
- Migita, T.; Narita, T.; Nomura, K.; Miyagi, E.; Inazuka, F.; Matsuura, M.; Ushijima, M.; Mashima, T.; Seimiya, H.; Satoh, Y.; et al. ATP citrate lyase: Activation and therapeutic implications in non-small cell lung cancer. Cancer Res. 2008, 68, 8547–8554. [Google Scholar] [CrossRef]
- Bose, S.; Ramesh, V.; Locasale, J.W. Acetate Metabolism in Physiology, Cancer, and Beyond. Trends Cell Biol. 2019, 29, 695–703. [Google Scholar] [CrossRef]
- Horibata, Y.; Ando, H.; Itoh, M.; Sugimoto, H. Enzymatic and transcriptional regulation of the cytoplasmic acetyl-CoA hydrolase ACOT12. J. Lipid Res. 2013, 54, 2049–2059. [Google Scholar] [CrossRef]
- Yoon, S.; Combs, J.; Falzone, A.; Prieto-Farigua, N.; Caldwell, S.; Ackerman, H.; Flores, E.; DeNicola, G. Comprehensive Metabolic Tracing Reveals the Origin and Catabolism of Cysteine in Mammalian Tissues and Tumors. Cancer Res. 2023, 83, 1426–1442. [Google Scholar] [CrossRef] [PubMed]
- Watts, S.D.; Torres-Salazar, D.; Divito, C.B.; Amara, S.G. Cysteine transport through excitatory amino acid transporter 3 (EAAT3). PLoS ONE 2014, 9, e109245. [Google Scholar] [CrossRef] [PubMed]
- García-Gaytán, A.C.; Hernández-Abrego, A.; Díaz-Muñoz, M.; Méndez, I. Glutamatergic system components as potential biomarkers and therapeutic targets in cancer in non-neural organs. Front. Endocrinol. 2022, 13, 1029210. [Google Scholar] [CrossRef] [PubMed]
- Bonifácio, V.D.B.; Pereira, S.A.; Serpa, J.; Vicente, J.B. Cysteine metabolic circuitries: Druggable targets in cancer. Br. J. Cancer 2021, 124, 862–879. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fan, J.; Venneti, S.; Cross, J.R.; Takagi, T.; Bhinder, B.; Djaballah, H.; Kanai, M.; Cheng, E.H.; Judkins, A.R.; et al. Asparagine plays a critical role in regulating cellular adaptation to glutamine depletion. Mol. Cell 2014, 56, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, N.N.; Hui, S.; Ghergurovich, J.M.; Fan, J.; Intlekofer, A.M.; White, R.M.; Rabinowitz, J.D.; Thompson, C.B.; Zhang, J. As Extracellular Glutamine Levels Decline, Asparagine Becomes an Essential Amino Acid. Cell Metab. 2018, 27, 428.e5–438.e5. [Google Scholar] [CrossRef]
- Jiang, J.; Pavlova, N.N.; Zhang, J. Asparagine, a critical limiting metabolite during glutamine starvation. Mol. Cell Oncol. 2018, 5, e1441633. [Google Scholar] [CrossRef]
- Gadian, D.G. NMR and Its Applications to Living Systems; Oxford University Press: New York, NY, USA, 1996. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Louis et al., 2016 [18] | ||||
|---|---|---|---|---|
| Training Cohort | Validation Cohort | |||
| LC | C | LC | C | |
| Number of subjects, n | 233 | 226 | 98 | 89 |
| Sensitivity (%) | 78 | 71 | ||
| Specificity (%) | 92 | 81 | ||
| R2X (Cum) | 0.864 | - | ||
| R2Y (Cum) | 0.477 | - | ||
| Q2 (Cum) | 0.391 | - | ||
| AUC | 0.88 | 0.84 | ||
| Louis et al., 2016 [19] | ||||
|---|---|---|---|---|
| Training Cohort | Validation Cohort | |||
| LC | BC | LC | BC | |
| Number of subjects, n | 54 | 80 | 81 | 60 |
| Sensitivity (%) | 93 | 89 | ||
| Specificity (%) | 99 | 82 | ||
| R2X (Cum) | 0.82 | - | ||
| R2Y (Cum) | 0.73 | - | ||
| Q2 (Cum) | 0.63 | - | ||
| AUC | 0.96 | 0.94 | ||
| Derveaux et al., 2021 [20] | ||||
|---|---|---|---|---|
| Training Cohort | Validation Cohort | |||
| LC | C | LC | C | |
| Number of subjects, n | 80 | 80 | 34 | 38 |
| Sensitivity (%) | 85 | 74 | ||
| Specificity (%) | 93 | 74 | ||
| R2X (Cum) | 0.861 | - | ||
| R2Y (Cum) | 0.581 | - | ||
| Q2 (Cum) | 0.364 | - | ||
| AUC | 0.95 | - | ||
| Derveaux et al., 2023 [21] | |||||
|---|---|---|---|---|---|
| B/E | C/E | B/E | |||
| OPLS-DA | OPLS-EP | OPLS-DA | OPLS-EP | OPLS-DA | |
| TRAINING COHORT | |||||
| Number of subjects, n | 50 | 50 | 50 | 50 | 50 |
| Sensitivity (%) | 92 | - | 88 | - | 74 |
| Specificity (%) | 96 | - | 90 | - | 62 |
| R2X (Cum) | 0.55 | 0.59 | 0.53 | 0.57 | 0.31 |
| R2Y (Cum) | 0.67 | 0.89 | 0.61 | 0.83 | 0.15 |
| Q2 (Cum) | 0.42 | 0.76 | 0.36 | 0.60 | 0.08 |
| AUC | 0.99 | - | 0.97 | - | 0.72 |
| VALIDATION COHORT | |||||
| Number of subjects, n | 24 | 24 | 23 | 23 | 23 |
| Sensitivity (%) | 88 | - | 96 | - | 74 |
| Specificity (%) | 92 | - | 91 | - | 43 |
| AUC | 0.97 | - | 0.97 | - | 0.69 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meynen, J.; Adriaensens, P.; Criel, M.; Louis, E.; Vanhove, K.; Thomeer, M.; Mesotten, L.; Derveaux, E. Plasma Metabolite Profiling in the Search for Early-Stage Biomarkers for Lung Cancer: Some Important Breakthroughs. Int. J. Mol. Sci. 2024, 25, 4690. https://doi.org/10.3390/ijms25094690
Meynen J, Adriaensens P, Criel M, Louis E, Vanhove K, Thomeer M, Mesotten L, Derveaux E. Plasma Metabolite Profiling in the Search for Early-Stage Biomarkers for Lung Cancer: Some Important Breakthroughs. International Journal of Molecular Sciences. 2024; 25(9):4690. https://doi.org/10.3390/ijms25094690
Chicago/Turabian StyleMeynen, Jill, Peter Adriaensens, Maarten Criel, Evelyne Louis, Karolien Vanhove, Michiel Thomeer, Liesbet Mesotten, and Elien Derveaux. 2024. "Plasma Metabolite Profiling in the Search for Early-Stage Biomarkers for Lung Cancer: Some Important Breakthroughs" International Journal of Molecular Sciences 25, no. 9: 4690. https://doi.org/10.3390/ijms25094690
APA StyleMeynen, J., Adriaensens, P., Criel, M., Louis, E., Vanhove, K., Thomeer, M., Mesotten, L., & Derveaux, E. (2024). Plasma Metabolite Profiling in the Search for Early-Stage Biomarkers for Lung Cancer: Some Important Breakthroughs. International Journal of Molecular Sciences, 25(9), 4690. https://doi.org/10.3390/ijms25094690