
The Inhibition of TREK-1 K+ Channels via Multiple Compounds Contained in the Six Kamikihito Components, Potentially Stimulating Oxytocin Neuron Pathways

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
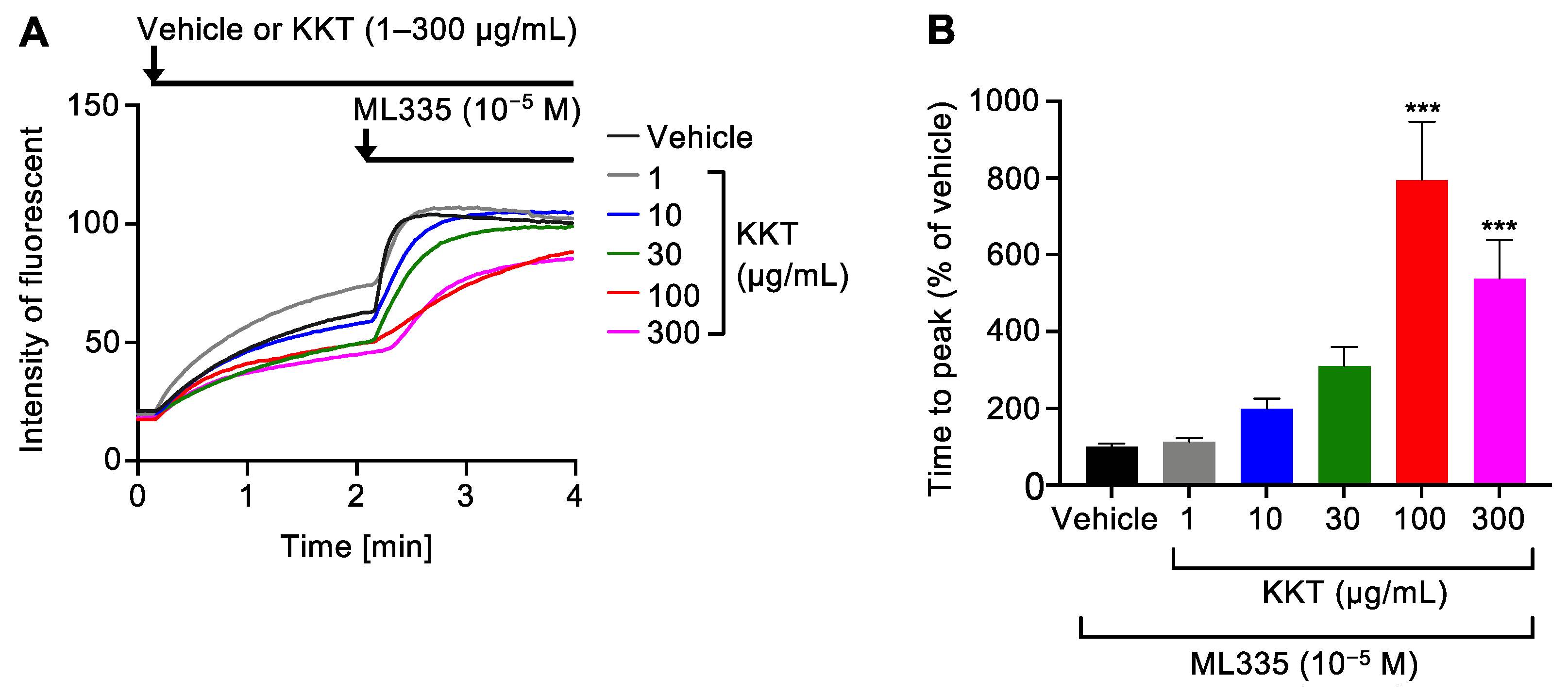
2.1. KKT Inhibited TREK-1 Activity Induced via a TREK-1 Agonist, ML335, in a Dose-Dependent Manner
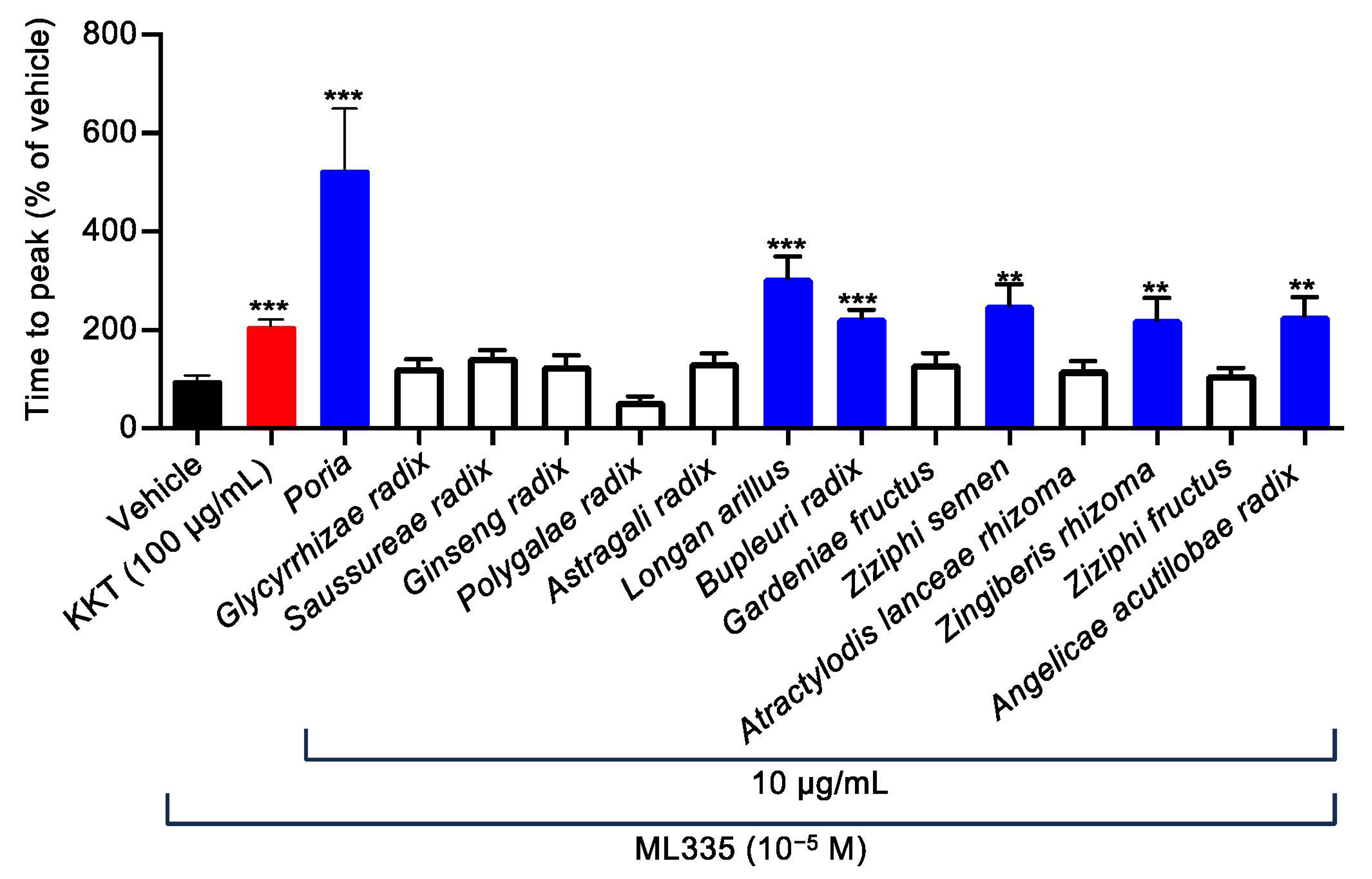
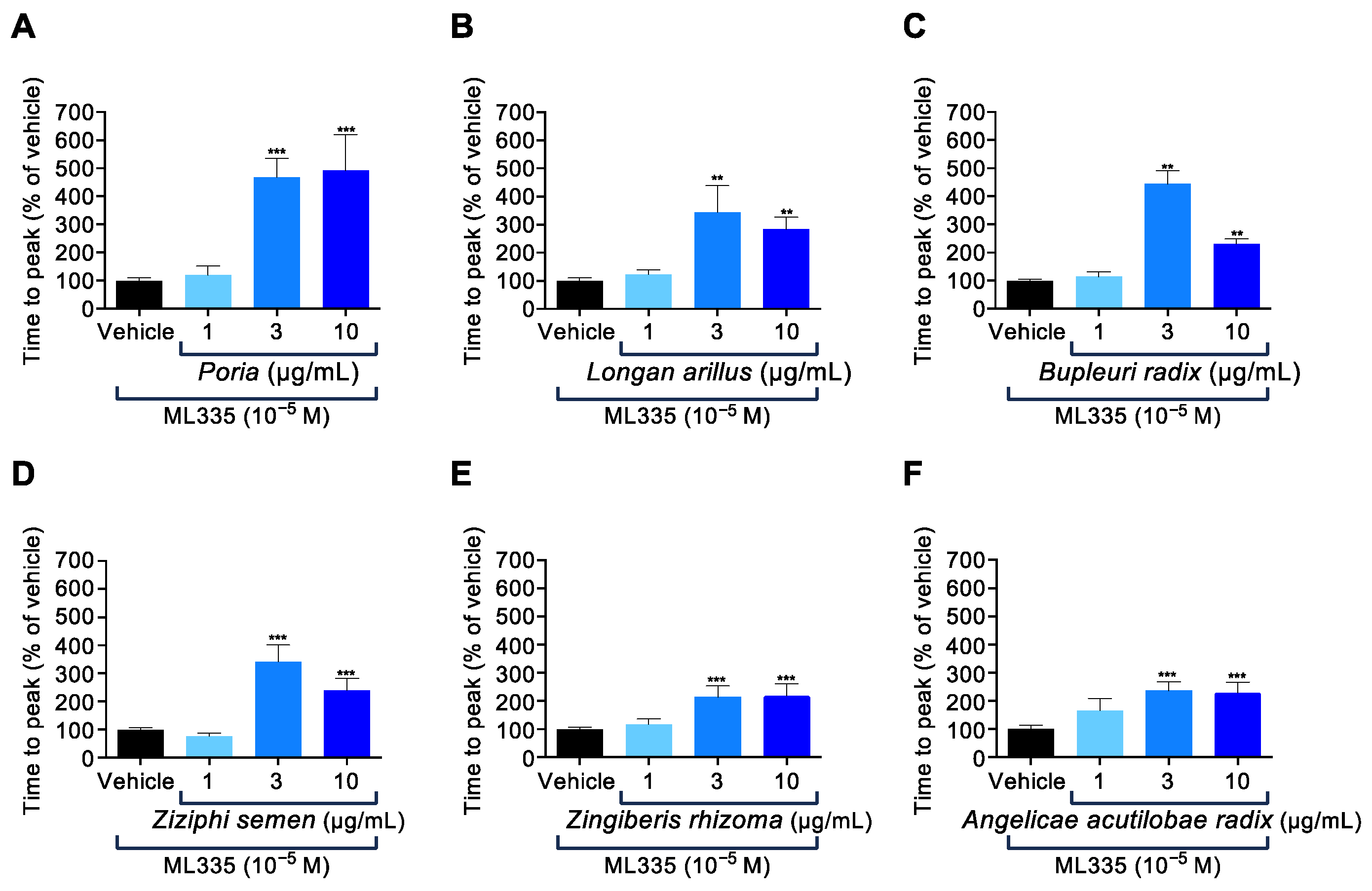
2.2. Six of the Fourteen Herbal Components Contained in KKT Were Involved in TREK-1 Inhibition
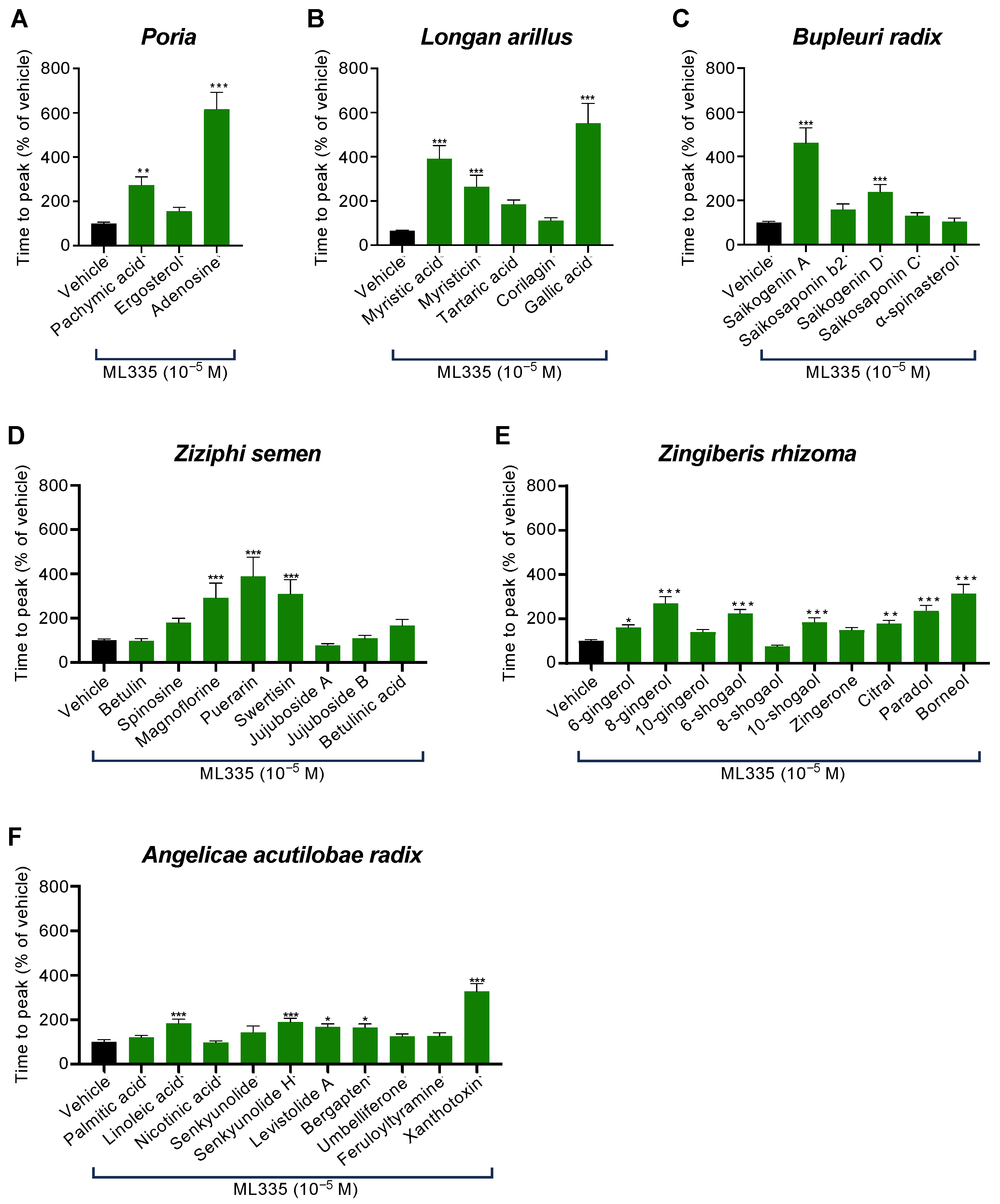
2.3. Specific Compounds Contained in Six of the Fourteen Herbal Components of KKT Were Involved in TREK-1 Inhibition
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plasmid Constructs and Transfection
4.3. Cell Culture
4.4. Measurement of TREK-1 Activity Using FluxORTM Potassium Ion Channel Assay
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dale, H.H. On some physiological actions of ergot. J. Physiol. 1906, 34, 163–206. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M.; Apps, D.; Menzies, J.; Patel, J.C.; Rice, M.E. Dendritic release of neurotransmitters. Compr. Physiol. 2016, 7, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S.; Kenkel, W.M.; MacLean, E.L.; Wilson, S.R.; Perkeybile, A.M.; Yee, J.R.; Ferris, C.F.; Nazarloo, H.P.; Porges, S.W.; Davis, J.M.; et al. Is oxytocin “Nature’s medicine”? Pharmacol. Rev. 2020, 72, 829–861. [Google Scholar] [CrossRef] [PubMed]
- Maejima, Y.; Horita, S.; Yokota, S.; Ono, T.; Proks, P.; Yoshida-Komiya, H.; Ueta, Y.; Nishimori, K.; Misaka, S.; Shimomura, K. Identification of oxytocin receptor activating chemical components from traditional Japanese medicines. J. Food Drug Anal. 2021, 29, 653–675. [Google Scholar] [CrossRef] [PubMed]
- Tsukada, M.; Ikemoto, H.; Lee, X.-P.; Takaki, T.; Tsuchiya, N.; Mizuno, K.; Inoue, T.; Tsunokawa, Y.; Okumo, T.; Matsuyama, T.; et al. Kamikihito, a traditional Japanese Kampo medicine, increases the secretion of oxytocin in rats with acute stress. J. Ethnopharmacol. 2021, 276, 114218. [Google Scholar] [CrossRef]
- Leckman, J.F.; Goodman, W.K.; North, W.G.; Chappell, P.B.; Price, L.H.; Pauls, D.L.; Anderson, G.M.; Riddle, M.A.; McDougle, C.J.; Barr, L.C.; et al. The role of central oxytocin in obsessive compulsive disorder and related normal behavior. Psychoneuroendocrinology 1994, 19, 723–749. [Google Scholar] [CrossRef] [PubMed]
- Talley, E.M.; Solorzano, G.; Lei, Q.; Kim, D.; Bayliss, D.A. Cns distribution of members of the two-pore-domain (KCNK) potassium channel family. J. Neurosci. 2001, 21, 7491–7505. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, P.; Singh, S.; Ravichandiran, V. Two-pore domain potassium channel in neurological disorders. J. Membr. Biol. 2021, 254, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Chemin, J.; Patel, A.J.; Duprat, F.; Sachs, F.; Lazdunski, M.; Honore, E. Up- and down-regulation of the mechano-gated K(2P) channel TREK-1 by PIP (2) and other membrane phospholipids. Pflugers Arch. 2007, 455, 97–103. [Google Scholar] [CrossRef]
- Chemin, J.; Patel, A.J.; Delmas, P.; Sachs, F.; Lazdunski, M.; Honore, E. Regulation of the mechano-gated K2P channel TREK-1 by membrane phospholipids. Curr. Top. Membr. 2007, 59, 155–170. [Google Scholar] [CrossRef]
- Maingret, F.; Patel, A.J.; Lesage, F.; Lazdunski, M.; Honoré, E. Mechano- or acid stimulation, two interactive modes of activation of the TREK-1 potassium channel. J. Biol. Chem. 1999, 274, 26691–26696. [Google Scholar] [CrossRef] [PubMed]
- Enyeart, J.A.; Danthi, S.J.; Enyeart, J.J. TREK-1 K+ channels couple angiotensin II receptors to membrane depolarization and aldosterone secretion in bovine adrenal glomerulosa cells. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E1154–E1165. [Google Scholar] [CrossRef] [PubMed]
- Enyeart, J.A.; Liu, H.; Enyeart, J.J. Curcumin inhibits bTREK-1 K+ channels and stimulates cortisol secretion from adrenocortical cells. Biochem. Biophys. Res. Commun. 2008, 370, 623–628. [Google Scholar] [CrossRef]
- Schroeter, C.B.; Nelke, C.; Schewe, M.; Spohler, L.; Herrmann, A.M.; Müntefering, T.; Huntemann, N.; Kuzikov, M.; Gribbon, P.; Albrecht, S.; et al. Validation of TREK1 ion channel activators as an immunomodulatory and neuroprotective strategy in neuroinflammation. Biol. Chem. 2023, 404, 355–375. [Google Scholar] [CrossRef] [PubMed]
- Lolicato, M.; Arrigoni, C.; Mori, T.; Sekioka, Y.; Bryant, C.; Clark, K.A.; Minor Jr, D.L. K2P2.1 (TREK-1)–activator complexes reveal a cryptic selectivity filter binding site. Nature 2017, 547, 364–368. [Google Scholar] [CrossRef]
- Mathie, A.; Veale, E.L.; Cunningham, K.P.; Holden, R.G.; Wright, P.D. Two-pore domain potassium channels as drug targets: Anesthesia and beyond. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 401–420. [Google Scholar] [CrossRef]
- Djillani, A.; Mazella, J.; Heurteaux, C.; Borsotto, M. Role of TREK-1 in health and disease, focus on the central nervous system. Front. Pharmacol. 2019, 10, 379. [Google Scholar] [CrossRef]
- Han, J.; Gnatenco, C.; Sladek, C.D.; Kim, D. Background and tandem-pore potassium channels in magnocellular neurosecretory cells of the rat supraoptic nucleus. J. Physiol. 2003, 546, 625–639. [Google Scholar] [CrossRef] [PubMed]
- Herbrechter, R.; Beltrán, L.R.; Ziemba, P.M.; Titt, S.; Lashuk, K.; Gottemeyer, A.; Levermann, J.; Hoffmann, K.M.; Beltrán, M.; Hatt, H.; et al. Effect of 158 herbal remedies on human TRPV1 and the two-pore domain potassium channels KCNK2, 3 and 9. J. Tradit. Complement. Med. 2020, 10, 446–453. [Google Scholar] [CrossRef]
- Wu, F.; Sun, H.; Gong, W.; Li, X.; Pan, Z.; Shan, H.; Zhang, Z. Genetic and pharmacological inhibition of two-pore domain potassium channel TREK-1 alters depression-related behaviors and neuronal plasticity in the hippocampus in mice. CNS Neurosci. Ther. 2021, 27, 220–232. [Google Scholar] [CrossRef]
- Djillani, A.; Pietri, M.; Mazella, J.; Heurteaux, C.; Borsotto, M. Fighting against depression with TREK-1 blockers: Past and future. A focus on spadin. Pharmacol. Ther. 2019, 194, 185–198. [Google Scholar] [CrossRef]
- Singh, S. From exotic spice to modern drug? Cell 2007, 130, 765–768. [Google Scholar] [CrossRef] [PubMed]
- Adachi, N.; Sakhri, F.Z.; Ikemoto, H.; Ohashi, Y.; Kato, M.; Inoue, T.; Hisamitsu, T.; Sunagawa, M. Kamikihito rescued depressive-like behaviors and hippocampus neurogenesis in chronic restraint stress rats. J. Tradit. Complement. Med. 2022, 12, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Uezono, Y.; Miyano, K.; Sudo, Y.; Suzuki, M.; Shiraishi, S.; Terawaki, K. A review of traditional Japanese medicines and their potential mechanism of action. Curr. Pharm. Des. 2012, 18, 4839–4853. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M. KAMPOmics: A framework for multidisciplinary and comprehensive research on Japanese traditional medicine. Gene 2022, 831, 146555. [Google Scholar] [CrossRef]
- Weaver, C.D.; Harden, D.; Dworetzky, S.I.; Robertson, B.; Knox, R.J. A thallium-sensitive, fluorescence-based assay for detecting and characterizing potassium channel modulators in mammalian cells. J. Biomol. Screen. 2004, 9, 671–677. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crude Drugs | Raw Materials |
|---|---|
| Astragali radix | Root of Astragalus membranaceus Bunge or Astragalus mongholicus Bunge |
| Bupleuri radix | Root of Bupleurum falcatum Linné |
| Ziziphi semen | Seed of Ziziphus jujuba Miller var. spinosa Hu ex H. F. Chou |
| Atractylodis lanceae rhizoma | Rhizome of Atractylodes lancea De Candolle or Atractylodes chinensis Koidzumi |
| Ginseng radix | Root of Panax ginseng C. A. Meyer |
| Poria | Sclerotium of Wolfiporia cocos Ryvarden et Gilbertson (Poria cocos Wolf) |
| Longan arillus | Arillus of Euphoria longana Lamarck |
| Polygalae radix | Root of Polygala tenuifolia Willdenow |
| Gardeniae fructus | Fruit of Gardenia jasminoides Ellis |
| Ziziphi fructus | Fruit of Ziziphus jujuba Miller var. inermis Rehder |
| Angelicae acutilobae radix | Root of Angelica acutiloba Kitagawa or Angelica acutiloba Kitagawa var. sugiyamae Hikino |
| Glycyrrhizae radix | Root of Glycyrrhiza uralensis Fischer or Glycyrrhiza glabra Linné |
| Zingiberis rhizoma | Rhizome of Zingiber officinale Roscoe |
| Saussureae radix | Root of Saussurea lappa Clarke |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miyano, K.; Nonaka, M.; Sakamoto, M.; Murofushi, M.; Yoshida, Y.; Komura, K.; Ohbuchi, K.; Higami, Y.; Fujii, H.; Uezono, Y. The Inhibition of TREK-1 K+ Channels via Multiple Compounds Contained in the Six Kamikihito Components, Potentially Stimulating Oxytocin Neuron Pathways. Int. J. Mol. Sci. 2024, 25, 4907. https://doi.org/10.3390/ijms25094907
Miyano K, Nonaka M, Sakamoto M, Murofushi M, Yoshida Y, Komura K, Ohbuchi K, Higami Y, Fujii H, Uezono Y. The Inhibition of TREK-1 K+ Channels via Multiple Compounds Contained in the Six Kamikihito Components, Potentially Stimulating Oxytocin Neuron Pathways. International Journal of Molecular Sciences. 2024; 25(9):4907. https://doi.org/10.3390/ijms25094907
Chicago/Turabian StyleMiyano, Kanako, Miki Nonaka, Masahiro Sakamoto, Mika Murofushi, Yuki Yoshida, Kyoko Komura, Katsuya Ohbuchi, Yoshikazu Higami, Hideaki Fujii, and Yasuhito Uezono. 2024. "The Inhibition of TREK-1 K+ Channels via Multiple Compounds Contained in the Six Kamikihito Components, Potentially Stimulating Oxytocin Neuron Pathways" International Journal of Molecular Sciences 25, no. 9: 4907. https://doi.org/10.3390/ijms25094907
APA StyleMiyano, K., Nonaka, M., Sakamoto, M., Murofushi, M., Yoshida, Y., Komura, K., Ohbuchi, K., Higami, Y., Fujii, H., & Uezono, Y. (2024). The Inhibition of TREK-1 K+ Channels via Multiple Compounds Contained in the Six Kamikihito Components, Potentially Stimulating Oxytocin Neuron Pathways. International Journal of Molecular Sciences, 25(9), 4907. https://doi.org/10.3390/ijms25094907