Abstract
Hidradenitis suppurativa (HS) is a chronic skin condition that primarily affects areas with dense hair follicles and apocrine sweat glands, such as the underarms, groin, buttocks, and lower breasts. Intense pain and discomfort in HS have been commonly noted, primarily due to the lesions’ effects on nearby tissues. Pain is a factor that can influence DNA methylation patterns, though its exact role in HS is not fully understood. We aim to identify molecular markers of chronic pain in HS patients. We performed DNA methylome of peripheral blood DNA derived from a group of 24 patients with HS and 24 healthy controls, using Illumina methylation array chips. We identified 253 significantly differentially methylated CpG sites across 253 distinct genes regulating pain sensitization in HS, including 224 hypomethylated and 29 hypermethylated sites. Several genes with pleiotropic roles include transporters (ABCC2, SLC39A8, SLC39A9), wound healing (MIR132, FGF2, PDGFC), ion channel regulators (CACNA1C, SCN1A), oxidative stress mediators (SCN8A, DRD2, DNMT1), cytochromes (CYP19A, CYP1A2), cytokines (TGFB1, IL4), telomere regulators (CSNK1D, SMAD3, MTA1), circadian rhythm (IL1R2, ABCG1, RORA), ultradian rhythms (PHACTR1, TSC2, ULK1), hormonal regulation (PPARA, NR3C1, ESR2), and the serotonin system (HTR1D, HTR1E, HTR3C, HTR4, TPH2). They also play roles in glucose metabolism (POMC, IRS1, GNAS) and obesity (DRD2, FAAH, MMP2). Gene ontology and pathway enrichment analysis identified 43 pathways, including calcium signaling, cocaine addiction, and nicotine addiction. This study identified multiple differentially methylated genes involved in chronic pain in HS, which may serve as biomarkers and therapeutic targets. Understanding their epigenetic regulation is crucial for personalized pain management and could enhance the identification of high-risk patients, leading to better preventative therapies and improved maternal and neonatal outcomes.
1. Introduction
Hidradenitis suppurativa (HS) is a chronic skin disease marked by recurrent painful nodules, abscesses, and sinus tracts in apocrine gland-rich areas, such as the armpits, groin, and buttocks, often accompanied by severe comorbidities [1]. HS is a multifactorial condition with an unclear exact cause, but it is widely accepted to result from a combination of genetic and epigenetic factors, environmental and lifestyle triggers, chronic inflammation, and immune system dysfunction, which collectively underlie its pathogenesis [2,3,4,5,6,7]. HS often goes undiagnosed [8], due to the absence of biological, pathological, or specific diagnostic tests [9]. Pain is a central aspect of living with HS, with intense discomfort commonly caused by the lesions’ effects on surrounding tissues [10]. Abscesses, inflammation, and scarring contribute to varying levels of pain, from mild to severe, depending on the disease’s stage and severity [11]. These symptoms profoundly impact the quality of life, leading to discomfort, social isolation, and stigmatization [12].
Pain management is crucial in HS, as both acute and chronic pain have a significant impact on the well-being of HS patients [13]. Psychological factors and lifestyle-related conditions, including stress, anxiety, depression, poor sleep, obesity, and diabetes, are common comorbidities of HS and significantly influence pain perception [14,15,16,17]. Acute pain occurs during flare-ups triggered by abscesses, inflammation, infection, or post-surgical trauma and is often accompanied by anxiety or emotional distress, which can usually be diagnosed and treated [18]. Chronic pain, which persists even between flare-ups, is often caused by scar tissue formation, nerve involvement, and the development of sinus tracts under the skin [13,19]. This persistent pain is exacerbated by factors like malodorous discharge and scarring and can lead to depression, anxiety, fatigue, irritability, muscle weakness, sleep disruptions, and social isolation [10,20]. Acute pain, however, can become chronic, and both acute and chronic pain significantly impact the quality of life for individuals with HS [13].
DNA methylation, a fundamental epigenetic mechanism, is pivotal in gene regulation and has been linked to various conditions that cause pain, including chronic pain, post-surgical complications, obesity, cancer, fibromyalgia, neuropathic pain disorders, arthritis, migraines, and endometriosis [21]. Although HS-related pain has been studied extensively, including in large population-based studies [13,22,23,24], the mechanisms of this condition are complex and multifactorial, involving interactions among genetic, environmental, and epigenetic factors [25]. Earlier studies have shown that DNA methylation in genes related to ion channels, transporters, cytochromes, cytokines, non-coding RNAs, circadian rhythms, telomeres, glucose metabolism, and neural pathways plays a significant role in pain perception across various diseases including HS [26,27,28,29,30,31,32,33,34]. Furthermore, several pain-associated genes, including OPRM1, ADRB2, CACNA2D3, GNA12, LPL, NAXD, and ASPHD1, exhibit methylation variations [35]. Despite these findings, the precise mechanism of pain perception remains unclear, with HS-related pain yet to be evaluated in the context of genome-wide DNA methylation changes.
This study examines the link between DNA methylation and chronic pain in HS patients to identify epigenetic biomarkers of pain severity. By analyzing differentially methylated CpG sites in pain-related genes, we reveal the pathways involved in pain sensitization, potentially guiding personalized treatments and new pain management strategies for HS.
2. Results
We identified 253 significantly differentially methylated genes in HS patients, which are implicated in pain mechanisms, by leveraging a pain-associated published database. These genes, linked to pain sensitization, were analyzed for their biological roles using public datasets and compared with genome-wide data (Supplementary Table S3). Table 1 highlights 224 hypomethylated and 29 hypermethylated CpG sites (Also provided Supplementary Table S1) tab, including genes involved in transport (ABCC2, SLC39A8, SLC39A9); apocrine gland genes (DOCK4, TUG1); dopamine-related genes (DDC, DRD2, and SLC6A3); serotonin-associated genes (HTR1D, HTR1E, HTR3C, HTR4, TPH2); endorphin-associated genes (BDNF MORC2 OPRM1 POMC); oxytosin-related (OXTR CD38); risk of suicide (DRD2, SLC6A3, BDNF, OPRM1); wound healing (MIR132, FGF2, PDGFC), autophagy genes (ZCCHC14, ULK1, MAP1LC3B, ESR2); ion channel regulators (CACNA1C, SCN1A); oxidative stress mediators (SCN8A, DNMT1, NR3C1, NOS3), cytochromes (CYP19A, CYP1A2), cytokines (TGFB1, IL4), telomere regulators (DNMT1, AHR), circadian rhythm (IL1R2, ABCG1, RORA GNA11), ultradian rhythms (PHACTR1, TSC2, ULK1, CLCN6); hypoxia related (MMP3, ACE, CX3CR1, NOTCH4); hormonal regulation (PPARA, NR3C1, ESR2), glucose metabolism (POMC, NR3C1, IRS1, GNAS), and obesity (DRD2, FAAH, MMP2, OPRM1).

Table 1.
Top 25 significantly differentially methylated CpG sites associated with pain in HS cases (based on FDR-adjusted p-values).
2.1. Heatmaps and Principal Component Analysis
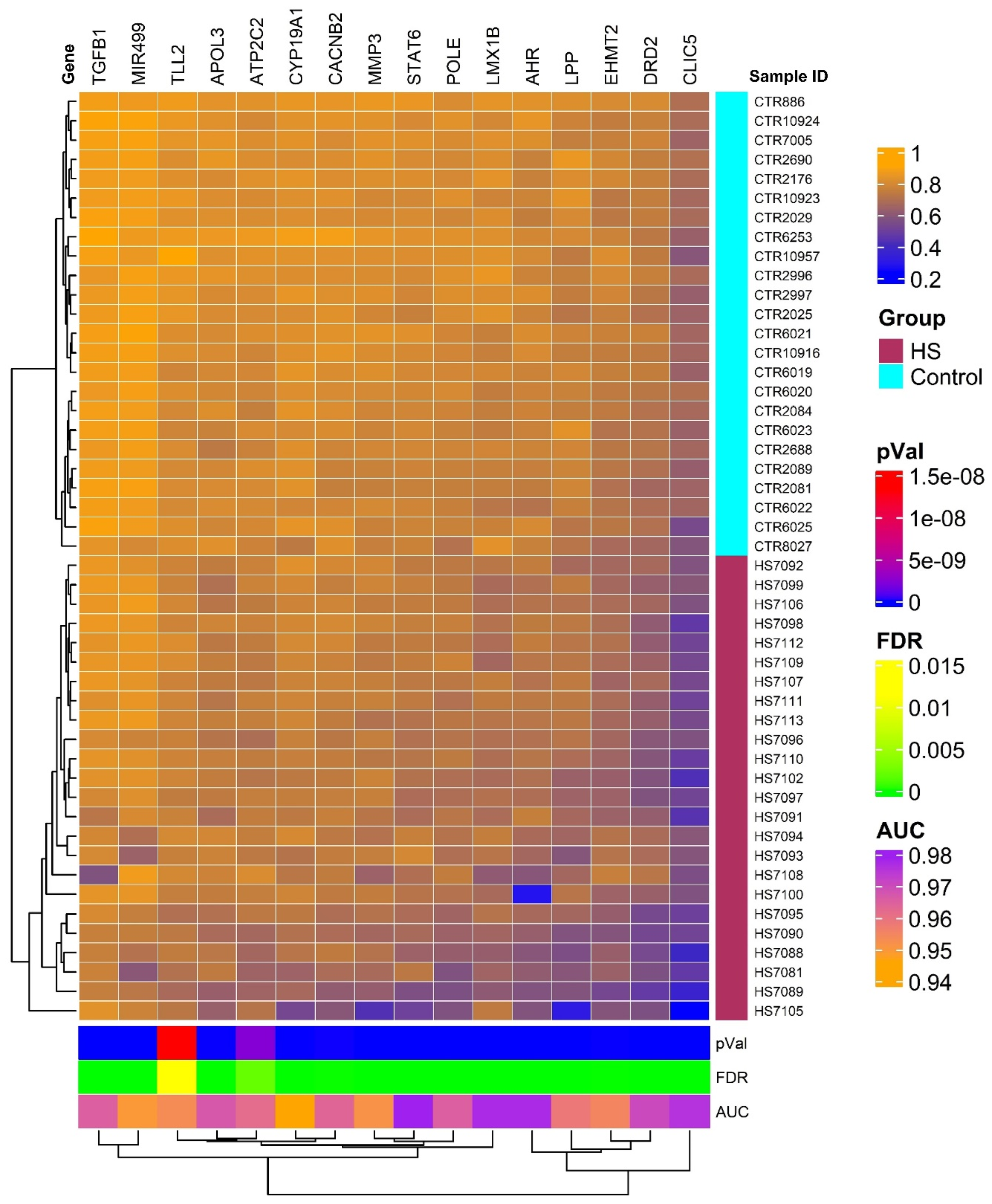
The heatmap generated from our analysis delineates two distinct clusters of genes with differential methylation profiles, one representing individuals with HS and another healthy controls. This visualization is depicted in Figure 1 and underscores the substantial methylation differences between the two groups, reflecting the pathophysiological alterations in HS patients compared to unaffected individuals. The genes selected for this heatmap all showed high discriminative power, with AUC values > 0.9 and FDR p-values ≤ 0.05, confirming their relevance in the methylation patterns associated with HS. The hierarchical clustering used in this analysis further supports the robustness of these methylation markers in distinguishing between affected and non-affected individuals.
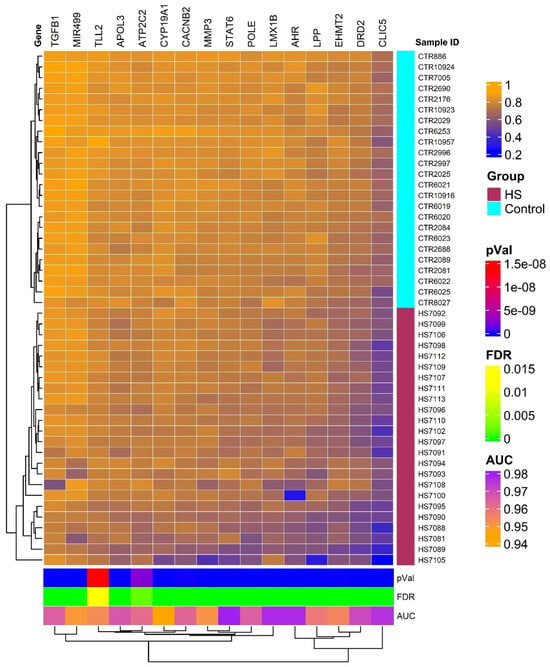
Figure 1.
Hierarchical clustering and heatmap of methylation data of 16 distinct genes with significant methylation differences between HS patients and controls based on AUC and FDR filters. The heatmap colors indicate levels of methylation as detailed in the color legend, with patients exhibiting the HS phenotype shown in maroon and normal controls in cyan. This clear segregation enhances our understanding of the potential biological mechanisms underlying HS-related pain and may facilitate the development of targeted therapies.
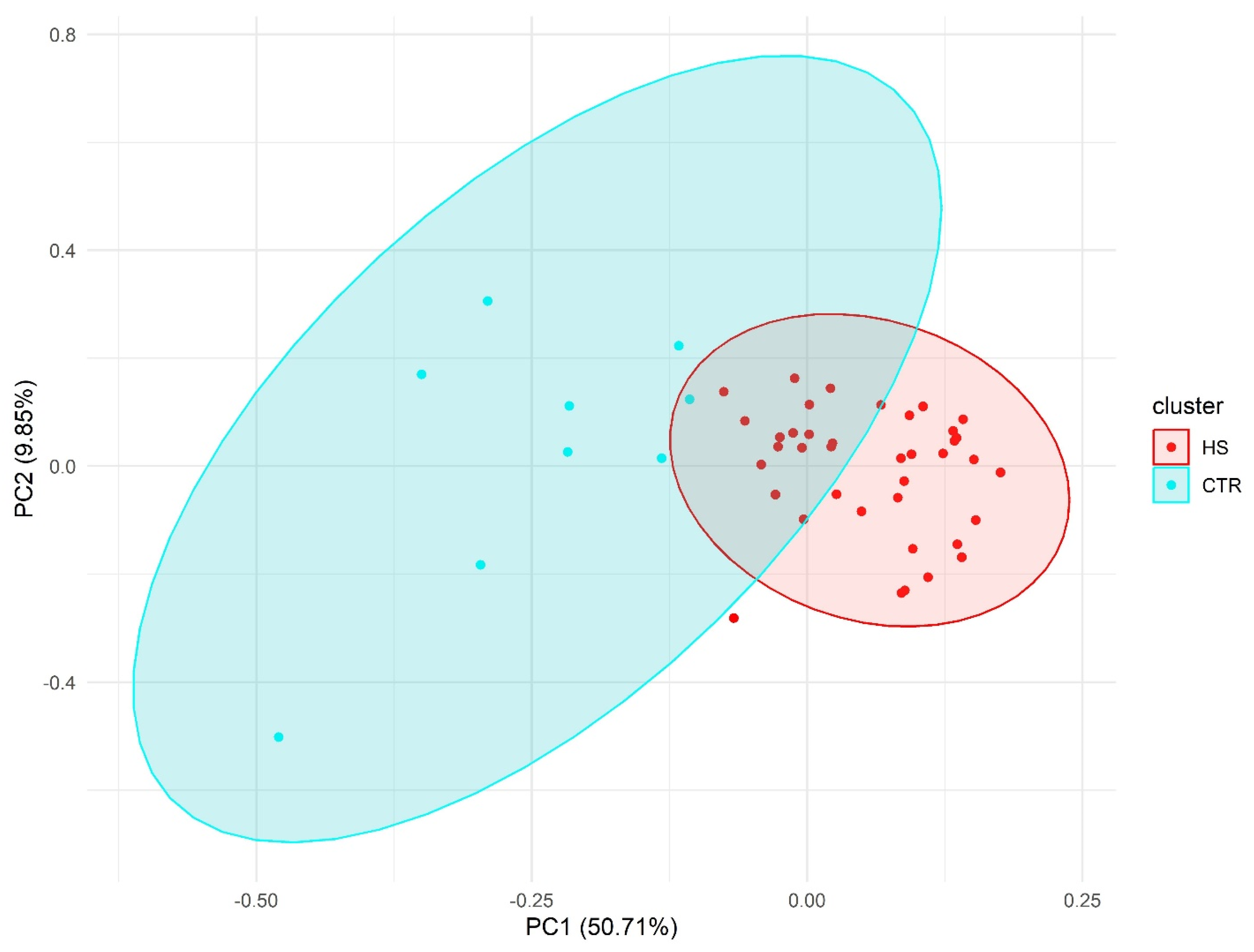
2.2. Principal Component Analysis
PCA results showed a clear separation between HS individuals and the control group (Figure 2).
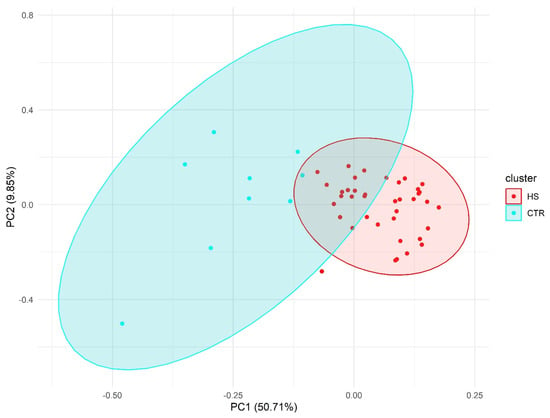
Figure 2.
Principal component analysis (PCA) using pain-associated genetic markers.
2.3. Network Analysis
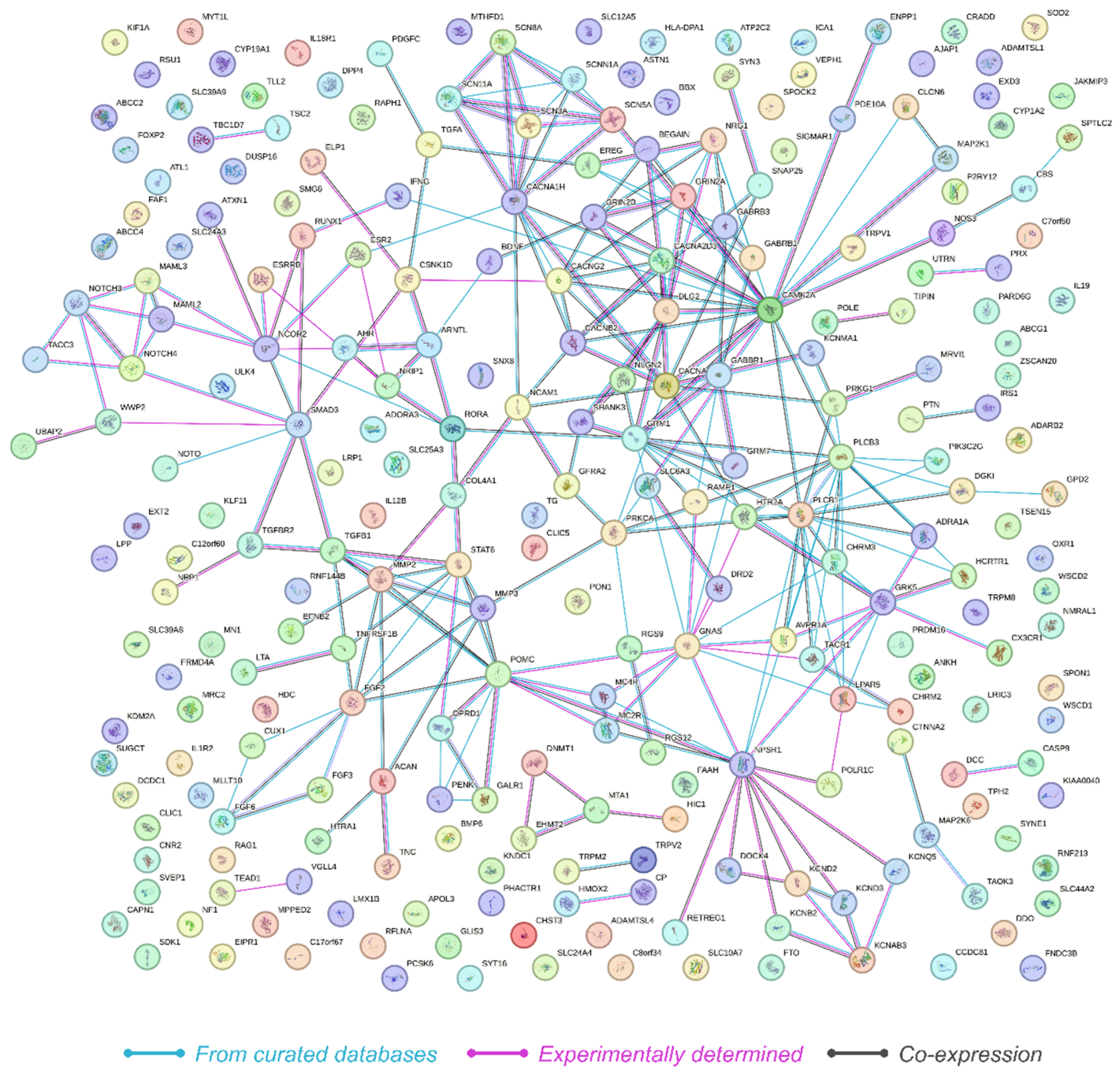
The protein–protein interaction (PPI) network constructed from the 249 genes revealed significant interaction dynamics as visualized in the PPI network (Figure 3). The network consisted of 249 nodes and 268 edges, with an average node degree of 2.15, indicating that on average, each protein interacts with approximately two other proteins. The average local clustering coefficient was calculated to be 0.321, suggesting a moderate level of clustering within the network. Importantly, the analysis revealed that the expected number of edges in a random network of similar size would be 110. The observed network had 268 edges, with a PPI enrichment p-value below 1.0 × 10−16. This highly significant enrichment indicates that the network has substantially more interactions than expected by chance, suggesting that the proteins are more interconnected and potentially involved in shared biological processes related to HS.
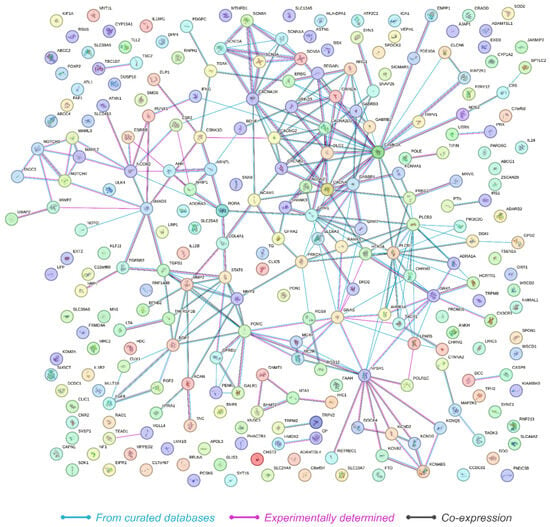
Figure 3.
Illustration of the protein–protein interaction network derived from STRING analysis, showing 249 interconnected genes with significant biological coherence, as indicated by a PPI enrichment p-value of <1.0 × 10−16. Nodes represent genes, which are color-coded to denote the source of interaction data, with lines reflecting connectivity.
2.4. Gene Ontology (GO) and KEGG Pathway Analysis
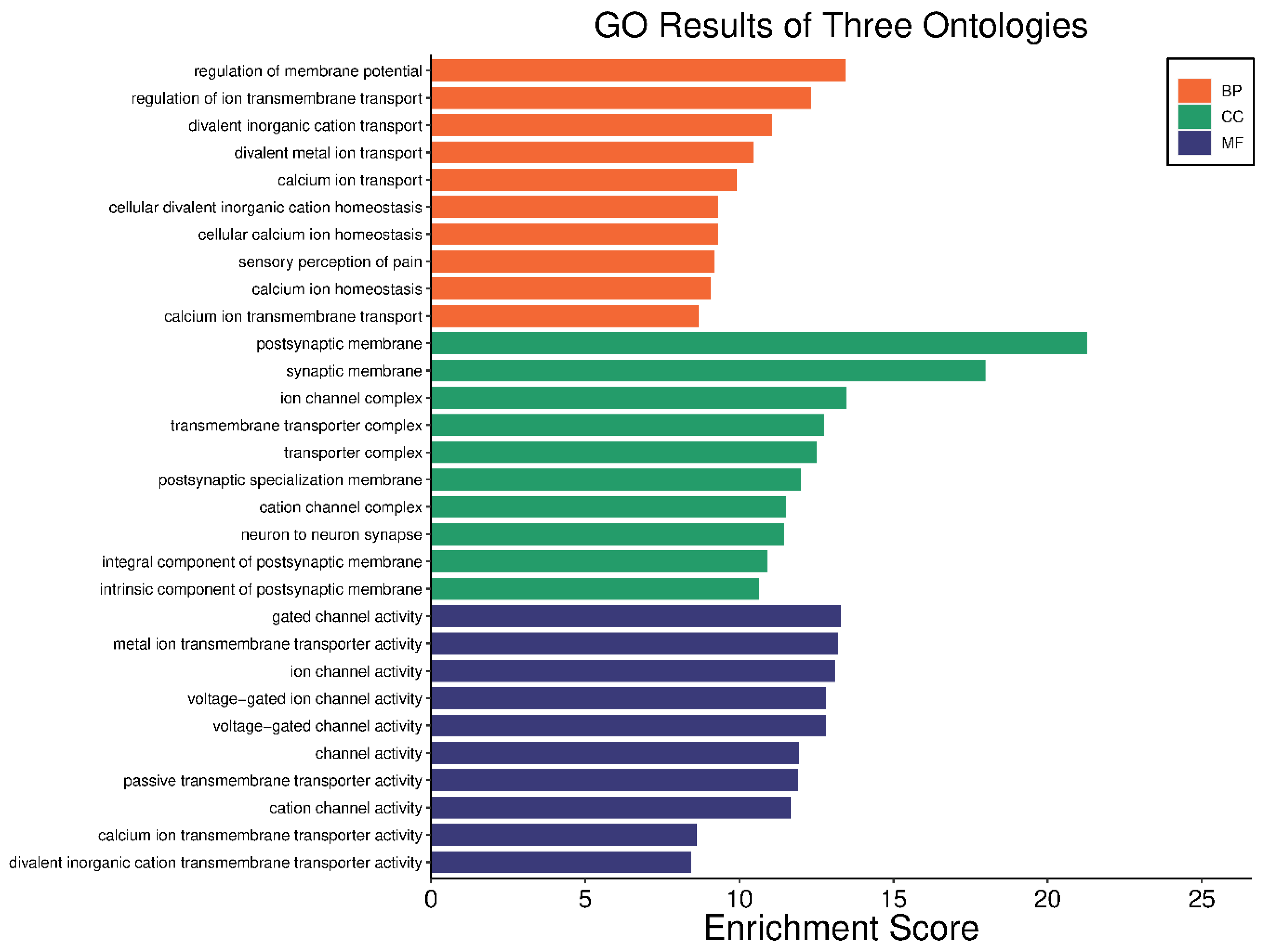
In our comprehensive gene ontology (GO) analysis, we identified significant enrichments across 688 biological processes (BP) terms, 64 cellular components (CC) terms, and 70 molecular functions (MF) terms, all with adjusted p-values < 0.05. For clarity and focus, the top ten teams from each category are visually represented in Figure 4. In the BP category, terms such as “regulation of membrane potential” and “regulation of ion transmembrane transport” were prominent, highlighting the importance of ion transport and membrane potential regulation in the pathophysiology of HS. Similarly, CC terms like “postsynaptic membrane” and “synaptic membrane” suggest alterations in synaptic structures that could play a critical role in the disease. In the MF category, terms such as “ion channel activity” and “voltage-gated ion channel activity” point to disruptions in ion channel functions, which are crucial for proper cell signaling and function. The complete list of enriched GO terms is available in Supplementary Tables S4–S6, providing a detailed breakdown of these categories.
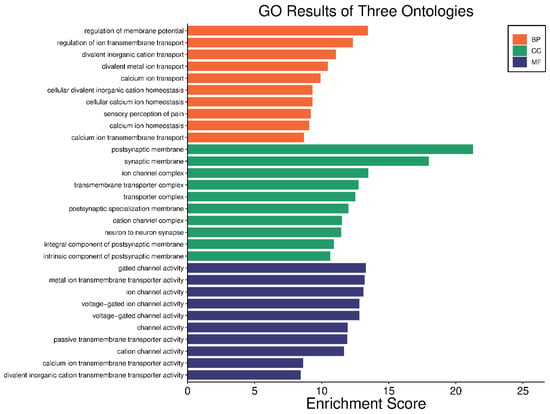
Figure 4.
Visual representation of the top ten GO terms; biological processes, cellular components, and molecular functions. Each bar represents the enrichment score of the terms, illustrating the relative significance and impact of each term.
2.5. KEGG Pathway Analysis
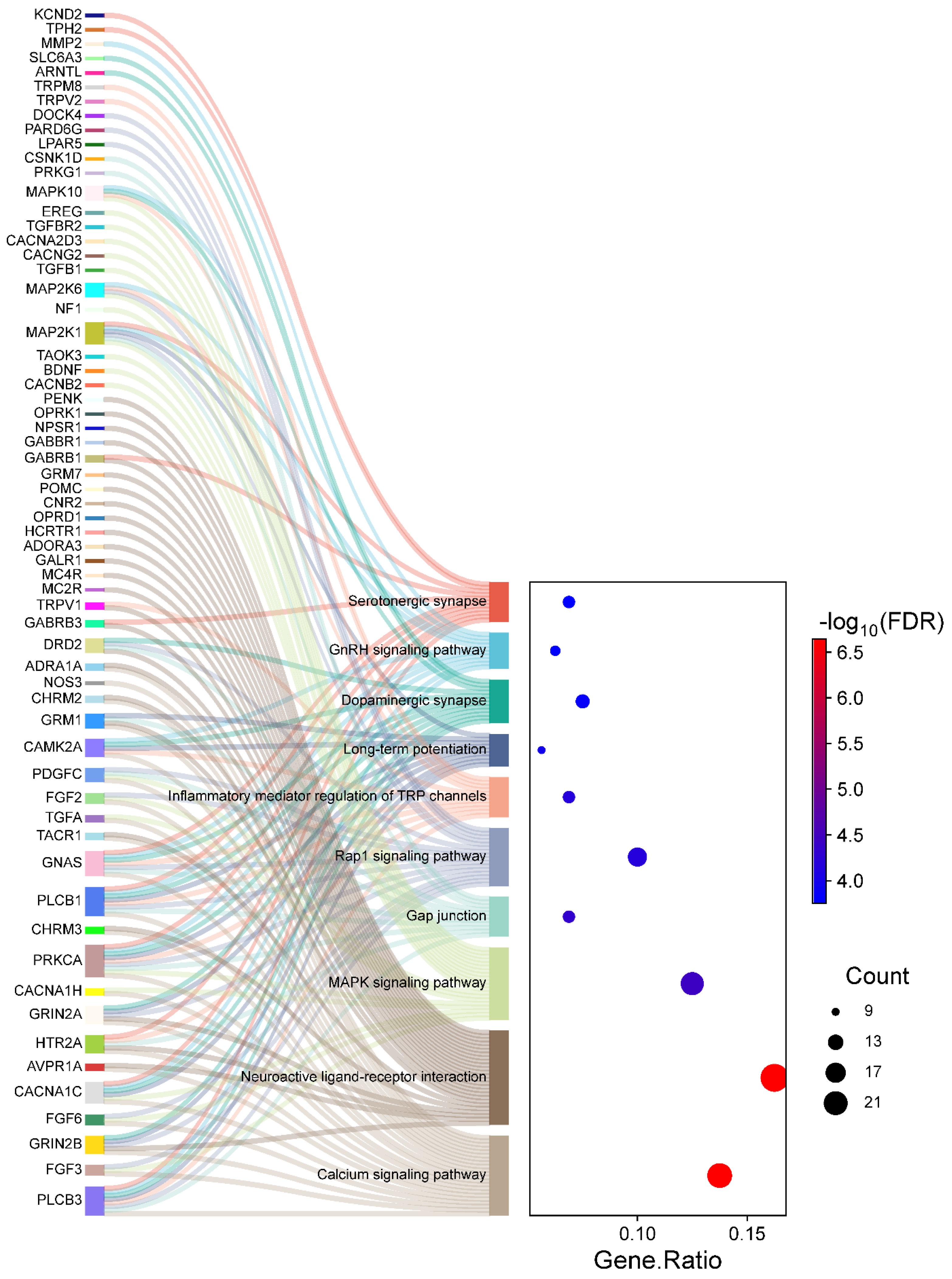
The KEGG pathway analysis identified 76 significantly enriched pathways, again with adjusted p-values < 0.05. For clarity and focus, the top ten pathways are visually represented in Figure 5. The top significantly enriched pathways identified in the KEGG analysis include the Calcium signaling pathway, Glutamatergic synapse, Long-term potentiation, and Circadian entrainment, all of which are integral to neuronal communication and inflammatory responses in HS. Notably, pathways related to neurotransmitter signaling and ion channel regulation, such as Calcium signaling and Glutamatergic synapse, underscore the role of disrupted neuronal processes in chronic pain mechanisms. As depicted in Figure 5, the left section (Sankey plot) visualizes the genes distributed across these enriched pathways, while the right section (dot plot) quantifies gene involvement. Specifically, the gene ratio measures the proportion of genes associated with each pathway relative to the total input genes, allowing for direct comparison of pathway involvement. Larger dots indicate higher gene counts within a pathway, and dot colors represent the statistical significance of the enrichment (FDR). Complete details of all enriched pathways are provided in Supplementary Table S7.
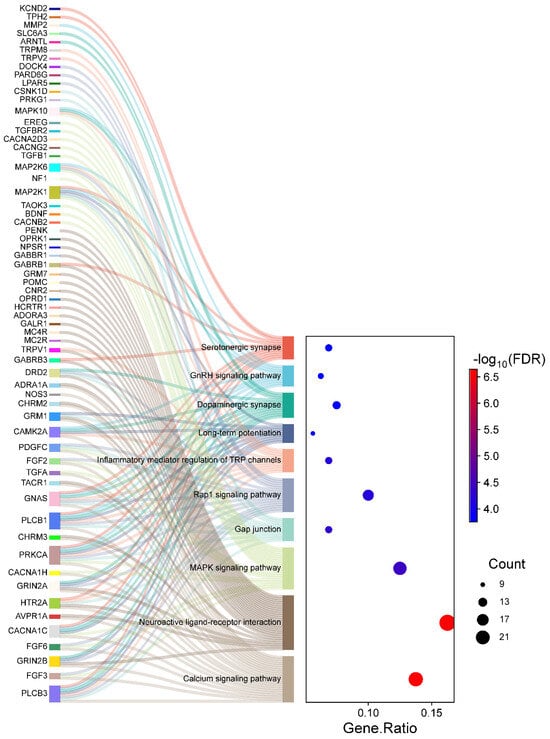
Figure 5.
Sankey diagram and dot plot depicting the mapping of genes to significantly enriched KEGG pathways in HS. The left section (Sankey diagram) illustrates the flow from individual genes to their respective pathways. The right section (dot plot) quantifies pathway enrichment, where dot sizes correspond to the number of genes associated with each pathway, and the dot colors represent the statistical significance (−log10 FDR).
We observed that pathways like Calcium signaling and Glutamatergic synapse exhibit high gene counts and strong enrichment significance, indicating their prominent role in pain sensitization and inflammatory dynamics in HS patients. Similarly, pathways like Long-term potentiation and Circadian entrainment suggest broader implications in neuronal plasticity and circadian rhythm disruptions, which may exacerbate pain perception and other neurological symptoms. These findings highlight the functional relevance of these pathways and the identified genes, offering insights into their potential as therapeutic targets for managing chronic pain in HS.
3. Discussion
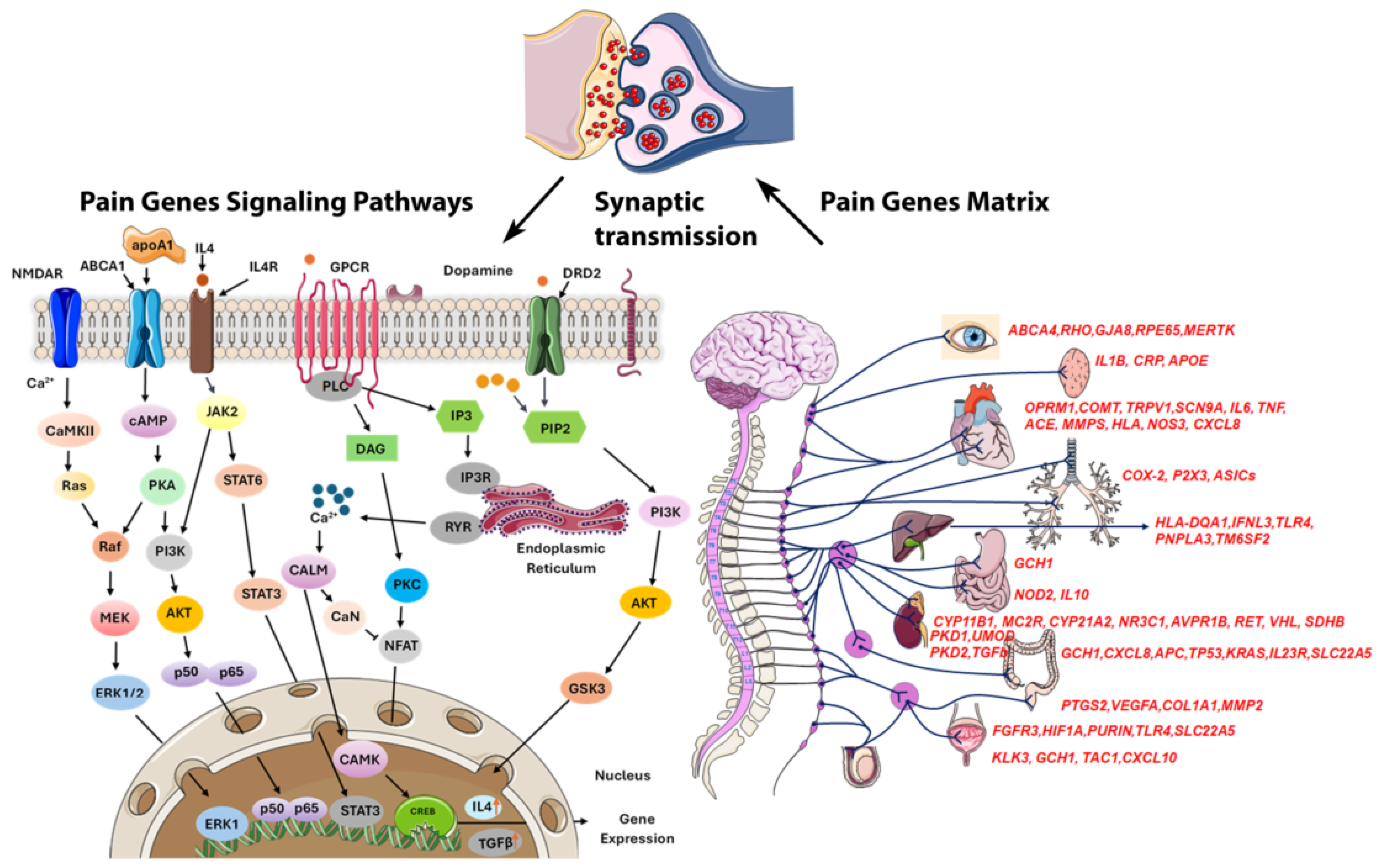
Managing pain in HS is challenging due to its chronic nature, frequent flare-ups, and complex pain mechanisms like inflammation, infection, and mechanical stress, complicating personalized treatment [13]. Epigenetics are crucial in pain perception, with future research aiming at gene therapies due to the link between DNA methylation and chronic pain [36]. We identified 253 differentially methylated pain-related genes, many of which exhibit pleiotropy, contributing to a wide range of biological functions, including circadian rhythm regulation, inflammation, telomere maintenance, glucose metabolism, autophagy, cytochrome activity, neurotransmission, oxidative stress, and obesity (Figure 6). Among these, 224 CpGs were hypomethylated, and 29 CpGs were hypermethylated. Notably, hypomethylation appears to exacerbate pain to a greater extent than hypermethylation, though the effects are highly context-dependent, varying by individual and condition [37].
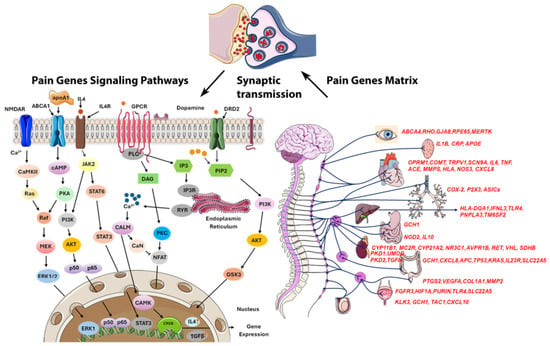
Figure 6.
Schematic representation of pain genes and their regulatory pathways showing that most pain transmission is mediated by noxious stimuli via synaptic transmission in the synaptic cleft. Several pain-related genes are commonly expressed across all organs, while different organs utilize specific genes to modulate pain perception through distinct pathways. In the brain, genes like TRPV1 and SCN9A are critical for detecting and modulating pain signals, while COMT influences pain sensitivity through neurotransmitter regulation. Inflammatory pain responses are prominent in all organs, with genes such as IL6, TNF, IL1B, IL10, and IFNL3 playing key roles. Overall, these genes modulate pain through inflammatory responses, nerve sensitivity, and tissue damage across various organs.
Autophagy and Chronic Pain: Recent evidence links dysregulated autophagy to chronic inflammation and pain [38]. We identified five key autophagy genes associated with pain: BDNF, ESR2, LMX1B, PRKCA, and ULK4. BDNF, which plays a role in brain signaling and synaptic plasticity, is proposed as a marker of nociception in chronic pain, particularly in the hippocampus, prefrontal cortex, and mesocorticolimbic system, potentially worsening chronic pain in HS by amplifying nociceptive signaling [39]. Estrogen receptor beta (ESR2) influences inflammation and pain perception, with signaling changes potentially increasing pain sensitivity in severe HS, especially in women with estrogen imbalances, highlighting the role of hormonal regulation in managing HS pain [40]. LMX1B influences skin development and inflammation, and its dysregulation could impact pain by affecting skin barrier integrity [41]. The PRKCA gene, encoding protein kinase C alpha (PKCα), plays a key role in signal transduction, inflammation, and pain modulation. It regulates pain signals, especially in inflammatory and neuropathic pain, by modulating ion channels like TRPV1 and voltage-gated calcium channels, both crucial for pain transmission [42]. ULK4 is crucial for the initiation and regulation of autophagy; disruptions in these kinases can lead to altered autophagic responses, exacerbating inflammation and pain [43].
Telomeres and Chronic Pain: Emerging studies suggest that telomere-related genes may impact pain perception and chronic pain severity [44]. We identified 31 dysregulated genes linked to telomere maintenance [44]. DNMT1, primarily known for its role in DNA methylation, shows emerging evidence linking it to pain modulation through epigenetic mechanisms [45]. Vitamin B12 deficiency can alter DNA methylation, potentially leading to DNMT1 dysregulation [46]. Our genome-wide study identified four dysregulated genes: CUBN (vitamin B12 absorption), AMN (works with cubilin for absorption), TCN1 (binds and protects vitamin B12), and SLC12A5 (cobalt carrier crucial for vitamin B12 production and various biological functions). Vitamin B12 may be a complementary treatment for pain in HS. [47]. CSNK1D is associated with pain sensitivity, influencing cellular processes through protein phosphorylation [48]. SMAD3, part of the TGF-β pathway, influences inflammation and tissue repair, impacting pain conditions [49]. DLG2, associated with synaptic function, plays a role in chronic pain by regulating neuronal pain signaling [50]. Other telomere-associated genes involved in pain regulation, such as CAMK2A, ESR2, AHR, and NOS3, are detailed elsewhere.
Circadian Rhythm and Chronic Pain: The circadian rhythm, a natural internal process regulating the sleeping–waking cycle and other physiological functions, significantly impacts pain perception, with sensitivity typically peaking in the late afternoon and being lowest in the early morning [51]. Additionally, ultradian rhythms, which occur more frequently than once per day, also influence pain sensitivity. Circadian rhythm-related genes play a key role in regulating processes such as sleep, hormone release, and inflammation. Disruption of these genes can alter pain perception, increasing sensitivity and exacerbating chronic pain by disturbing the body’s natural rhythms and affecting pain pathways and immune responses. We identified 22 dysregulated genes associated with circadian rhythms and 9 genes implicated in ultradian rhythms linked to pain regulation [52]. Disruptions, like shift work, contribute to chronic pain, which is worsened by poor sleep. Sleep disturbances and psychological stress, including depression and anxiety, can worsen pain and promote a sedentary lifestyle and overeating. This rhythm affects pain-modulating substances like melatonin. Chronic pain disrupts sleep, creating a cycle where pain and sleep issues reinforce each other, with individual variations influencing susceptibility and the effectiveness of relief.
Obesity contributes to chronic pain: Obesity is linked to chronic inflammation, which exacerbates pain and conditions like rheumatoid arthritis and fibromyalgia [52]. It also raises the risk of metabolic syndrome, cardiovascular issues, nerve damage, and neuropathic pain [53]. Obese individuals often require higher pain medication doses, complicating management. Reduced activity weakens muscles and increases the risk of chronic conditions, worsening pain. We identified 31 genes linked to both obesity and pain, including MC4R (appetite regulation and obesity), CYP19A1 (estrogen synthesis affecting body fat distribution), IRS1 (insulin signaling linked to obesity), FAAH (endocannabinoid system in appetite regulation), and DRD2 (dopamine in reward and pleasure affecting food consumption) [54], which play key roles in pain, obesity, and appetite regulation.
Glucose Metabolism and Chronic Pain: Glucose metabolism, which is vital for energy production, intricately influences pain perception through complex mechanisms [55]. We identified 19 genes related to glucose metabolism that may influence pain perception, including AHR, CACNA1C, CACNA1H, CP, CYP19A1, ENPP1, EXT2, GLIS3, GNAS, GPD2, IRS1, KLF11, MC2R, MC4R, MMP2, NOTCH3, POMC, STAT6, and TGFA. The interconnectedness of these genes with inflammatory, hormonal, and neural processes suggests a complex interplay between glucose metabolism and pain perception, emphasizing the need for further research [56]. Genes like CACNA1C (calcium regulation) and AHR (inflammation) may influence pain pathways, while NOTCH3 is involved in immune responses and stress regulation, and anxiety may contribute to pain modulation.
Chronic Pain Can Elevate Suicide Risk: Persistent pain increases suicide risk by causing distress, lowering quality of life, and triggering depression and anxiety. We identified 57 dysregulated pain-related genes linked to suicide, affecting neurotransmission, inflammation, sensory processing, stress response, and hormonal regulation. BDNF, which is crucial for neuronal survival and plasticity, is linked to mood disorders and suicide risk, while DRD2, involved in dopamine signaling, affects mood regulation and susceptibility to depression and suicidal tendencies [57]. Glutamate receptors, including GRIN2A and GRIN2B, are vital for glutamatergic neurotransmission, impacting mood and being associated with suicidal behavior [58]. Serotonin-related genes, including the serotonin receptor gene HTR2A, play a crucial role in neuropsychiatric processes by modulating serotonin signaling in the brain. TPH2 is vital for serotonin synthesis, ensuring adequate serotonin levels, while the serotonin transporter gene SLC6A4 regulates serotonin reuptake and mood stability. Disruptions in these genes are associated with mood disorders and an increased risk of suicide.
The FAAH, part of the endocannabinoid system, also contributes to mood regulation and susceptibility to mood disorders and suicidal behavior [59]. The POMC gene, which regulates stress and mood, is linked to depression and suicidal tendencies when dysregulated [60]. Genetic variations in NOS3, affecting nitric oxide production, and TGFB1, involved in inflammation and tissue repair, further illustrate the complex interactions between genetic predispositions and mental health outcomes, both potentially contributing to increased suicide risk if dysregulated [61,62].
Calcium and Chronic Pain: Calcium ions are crucial for pain management, influencing sensitivity, signal transmission, and keratinocyte function; their dysregulation can heighten pain and delay healing. [63]. We identified 23 dysregulated calcium-associated genes involved in pain in the HS cohort. The CACNA1C, CACNA2D3, CACNA1H, and CACNG2 genes encode various components and regulatory subunits of calcium channels vital for calcium signaling, affecting neurotransmitter release and excitability. CACNA1C and CACNA1H encode L-type and T-type calcium channels, respectively, which are key in pain signal transmission [64]. CACNA2D3 and CACNG2 also affect calcium signaling, and their dysfunction can exacerbate pain and emotional distress [65,66]. CAMK2A, involved in pain pathway sensitization, may perpetuate pain hypersensitivity due to persistent inflammation in HS. CHRM2 modulates pain via acetylcholine signaling, and disruptions in cholinergic signaling in HS could worsen pain perception. Additionally, studies in animals have shown that CHRM3 is essential for epidermal development, improving basal cell adhesion and inhibiting their proliferation [67]. NMDA receptor GRIN2A plays critical roles in synaptic plasticity and pain perception, with chronic pain in HS potentially associated with NMDA receptor-mediated central sensitization. GRM1 modulates pain through G-protein signaling and calcium mobilization, and its persistent activation in HS could enhance pain states, HCRTR1 influences pain sensitivity and stress-induced analgesia, and its dysregulation in HS could alter pain perception [68]. Serotonin receptor HTR2A modulates pain perception and analgesic efficacy [69].
Potassium channels: Pain perception in hidradenitis suppurativa may be significantly influenced by the dysregulation of potassium channels, including genes such as KCNMA1, KCNQ5, KCNB2, KCND2, KCND3, and KCNAB3, all of which were identified in this study. KCNMA1 encodes a large-conductance calcium-activated potassium channel crucial for regulating neuronal excitability and pain signal transmission [70]. Dysregulation of KCNMA1 can alter neuronal excitability, potentially worsening pain sensitivity in HS. KCNB2, KCND2, and KCND3 are critical for voltage-gated potassium channels, which maintain normal neuronal function [71]. KCNB2 regulates action potentials and neuronal excitability, impacting pain signal propagation. KCND2 controls action potentials and firing rates, affecting neuronal responsiveness and pain perception. KCND3 is vital for neuronal excitability and firing patterns, influencing pain processing. KCNQ5, also plays a role in voltage-gated potassium channels, further affecting neuronal activity and pain perception [72].
Sodium channels: We identified five dysregulated sodium channel genes: SCN11A, SCN3A, SCN5A, SCN8A, and SCN11A. They are mainly in peripheral neurons and nociceptors and transmit pain signals, with variations affecting pain thresholds [73]. SCN11A encodes a sodium channel essential for pain perception and neuronal excitability, with variants potentially disrupting pain signaling and exacerbating chronic pain [74]. SCN3A, upregulated in response to nerve injury, contributes to pain pathway development and hyperexcitability [75]. The SCN5A gene mainly regulates cardiac conduction but also affects sensory neurons and pain mechanisms, particularly in conditions with both cardiac and sensory symptoms [76]. SCN8A gene encodes the Naᵥ1.6 sodium channel, which is essential for action potential generation in neurons [77]. Variations in SCN8A can increase neuronal excitability, heightening pain sensitivity (hyperalgesia) or causing pain from non-painful stimuli (allodynia). The SCN11A gene encodes the Nav1.9 sodium channel, which is vital for pain perception. SCN11A variants are notably associated with familial episodic pain syndrome (FEPS), a severe pain disorder in early childhood.
Transporters and chronic pain: Transporters regulating ion and lipid balance are crucial for inflammation and pain. In HS, dysregulation of 12 key transporters, including ABCC2, ABCG1, and SLC genes, worsens chronic pain and inflammation. Genes such as ABCC2 and ABCC4 play a crucial role in the efflux of inflammatory mediators and drugs, and their dysregulation may lead to impaired drug metabolism and persistent inflammation, contributing to inadequate pain control in HS patients. Similarly, ABCG1 is pivotal in regulating lipid homeostasis and macrophage function, with its disruption potentially exacerbating inflammation and chronic pain [78]. SLC10A7 impacts inflammatory pathways and cellular homeostasis, while SLC12A5, also known as (KCC2), influences neuronal excitability, potentially leading to neuropathic pain due to the dysregulated inflammatory response in HS [79]. The SLC24A3 and SLC24A4 regulate intracellular sodium and calcium levels, with their dysregulation potentially disrupting ion balance and cellular signaling, contributing to inflammation and pain in HS. Mitochondrial function, as mediated by SLC25A3, is another critical factor, where impaired ATP production and increased oxidative stress may contribute to chronic pain and fatigue in HS [80]. Furthermore, SLC39A8 and SLC39A9 are key in zinc transport and immune regulation, with their dysregulation amplifying inflammatory responses and sustaining pain. SLC44A2 and SLC6A3 influence cholinergic and dopaminergic signaling, respectively, where alterations in these pathways may lead to heightened pain perception and chronic pain due to central sensitization in HS.
Addiction Caused by Chronic Pain: Opioids are commonly prescribed for post-surgical, cancer-related, and chronic pain management, but their use carries a substantial risk of addiction, particularly in those with chronic pain. We identified ten genes—BDNF, DRD2, CNR2, FAAH, GABBR1, SLC6A3, GRM1, OPRD1, and CHRM2—that may play crucial roles in neurotransmission and reward pathways, influencing susceptibility and response to substances. BDNF is essential for brain plasticity and the reward system, with certain variants linked to higher addiction risk [81]. DRD2 plays a key role in dopamine signaling, which influences reward, motivation, and pain perception, with variants influencing addiction and drug response [82]. CNR2, part of the endocannabinoid system, affects reward pathways and addiction [83]. FAAH modulates endocannabinoid metabolism and reward systems [84]. GABBR1 is involved in GABAergic neurotransmission, with variants potentially affecting addiction susceptibility [85]. SLC6A3 (DAT1) controls dopamine reuptake, influencing dopamine levels, mood, and pain sensitivity, with dysregulation often seen in chronic pain conditions, impacting addiction risk [82]. GRM1 and OPRD1 contribute to addiction through their roles in neurotransmission and opioid signaling, while CHRM2 influences cholinergic signaling, affecting reward and addiction pathways [86].
Immunomodulators and Chronic Pain: The immune system is crucial for maintaining homeostasis and responding to injury. Immunomodulators, including cytokines and chemokines, regulate the balance between pro-inflammatory and anti-inflammatory processes. Disruptions in these pathways can lead to chronic inflammation, a major factor in chronic pain in HS [52]. We identified 27 dysregulated genes in HS that act as immunomodulators, each playing distinct roles in regulating immune responses and pain perception. Notably, genes such as PRKCA, POMC, BDNF, and DNMT1 are involved in signaling pathways that modulate pain sensitivity and inflammatory responses [42,45,87]. SOD2, TGFB1, IFNG, and SMAD3 contribute to the management of oxidative stress and the regulation of both pro-inflammatory and anti-inflammatory cytokine production [52,88]. CX3CR1 and NCAM1 are associated with immune cell recruitment and neural activity, influencing inflammation and pain pathways [89,90]. Furthermore, RUNX1, MMP2, AHR, STAT6, TGFA, LTA, TNFRSF1B, IL1R2, and NRG1 play critical roles in immune regulation, cell survival, and inflammation, all of which impact chronic pain conditions. The involvement of EREG, FGF2, IL12B, IL19, IL18R1, DPP4, TNC, and TGFBR2 underscores the complex interplay between immune modulation and pain in HS, affecting various levels of gene expression and signaling cascades.
Endocrine Functions and Pain: Chronic pain in HS may be influenced by dysregulated hormonal and endocrine genes. We identified four genes involved in endocrine functions that could potentially play roles in pain mechanisms in HS. POMC produces peptides like ACTH and beta-endorphin, with beta-endorphins playing a key role in pain modulation [91]. Dysregulation of POMC can result in abnormal beta-endorphin levels, affecting pain sensitivity and inflammation in HS patients. TG is a glycoprotein essential for thyroid hormone synthesis. Although TG itself is not directly linked to pain, thyroid dysfunction, reflected by abnormal TG levels, can affect systemic inflammation and pain perception. Thyroid disorders linked to inflammatory skin conditions suggest that TG dysregulation may worsen pain in HS by affecting thyroid function and inflammation [92]. MC2R, a receptor for ACTH, regulates adrenal function and stress responses. MC2R signaling influences cortisol release, which in turn modulates inflammation and pain [93]. CYP19A1 (Aromatase) converts androgens to estrogens, which are known to modulate pain sensitivity [94]. Dysregulation of CYP19A1 can lead to imbalances in estrogen levels, affecting pain perception and inflammatory responses.
Metal-Linked Genes in Chronic Pain: Dysregulations in metal-associated genes could contribute to the chronic pain experienced by HS patients. Several metal-associated genes were found to be dysregulated in the study, including six zinc-related SLC39A8, SLC39A9, MIR132, MMP2, MMP3, and CBS. Zinc deficiency can lead to liver and pancreatic dysfunction, kidney impairment, chronic inflammation, delayed wound healing, and a weakened immune response, increasing vulnerability to various diseases. Dysregulation of SLC39A8 and SLC39A9 disrupts zinc balance, impairing immunity and raising oxidative stress. MIR132 heightens inflammation and pain sensitivity, while matrix metalloproteinases (MMPs), specifically MMP-2 and MMP-3, significantly impact the pathogenesis of hidradenitis suppurativa (HS). These enzymes are involved in the breakdown of extracellular matrix (ECM) components, such as collagen, which is crucial for maintaining skin integrity. The overexpression of MMP2 can lead to excessive ECM degradation, resulting in tissue destruction and the formation of sinus tracts. Elevated MMP3 levels can further exacerbate inflammation by processing pro-inflammatory cytokines, such as TNF-α and IL-1β, leading to a chronic inflammatory state. CBS dysfunction worsens oxidative stress and impairs cellular repair, all contributing to persistent pain in HS. Iron is crucial in HS by regulating oxidative stress, immune function, and inflammation. An imbalance in iron can exacerbate tissue damage, impair healing, and worsen HS symptoms. Dysregulated SOD2 increases oxidative stress and inflammation, CACNG2 and CACNA2D3 impair calcium channels and heighten pain sensitivity, BMP6 disruption hampers tissue repair and aggravates inflammation, and CP (ceruloplasmin) disruption worsens iron metabolism, further escalating oxidative stress and HS severity. As detailed elsewhere, manganese-related (SLC39A8), cobalt-linked (SLC12A5), and lithium-associated (CACNG2) were found to be dysregulated in the study. Manganese supports manganese superoxide dismutase (MnSOD), an enzyme that neutralizes oxidative stress. Since oxidative stress is central to chronic inflammation in HS, sufficient manganese levels are essential for managing oxidative damage in HS lesions. Cobalt is crucial for vitamin B12, which supports DNA synthesis and cellular metabolism. Imbalances in cobalt can disrupt these processes and potentially worsen inflammatory conditions like HS. Lithium affects mood, neuronal health, and cellular signaling, potentially influencing inflammatory and immune responses in HS, though its direct link to the condition is not well established.
Limitations: The study identifies potential pain markers for HS but has limitations, including the use of blood samples instead of lesion tissues. Blood samples were chosen for ease of access and minimal invasiveness [95], allowing longitudinal studies and capturing broad epigenetic changes linked to inflammation and immune responses. While lesion tissues might offer more localized data, blood profiles still reflect key molecular alterations associated with pain and disease severity. Future research should include tissue-specific analyses. Additionally, the study’s focus on Asian Indian HS patients limits geographic diversity, and broader populations should be included for generalizability. Although the identified differentially methylated CpG sites are promising, further validation is required to confirm their specificity for HS-related pain.
Conclusions: This study represents the first genome-wide analysis demonstrating that distinct DNA methylation patterns directly influence the pain modulatory system in HS. Our findings introduce a novel epigenetic mechanism, highlighting specific gene methylation signatures that are closely associated with pain perception and sensitivity in HS patients. These pain-related epigenetic modifications offer insights into the biological pathways underlying chronic pain in HS and hold promise as biomarkers. They could pave the way for targeted therapies, enabling personalized treatments that modulate pain pathways and improve outcomes. By linking epigenetic regulation to pain in HS, this research lays the groundwork for future studies to explore how these changes can be harnessed for clinical benefit, potentially leading to novel pain management strategies.
4. Materials and Methods
Study design: The Institutional Review Board of Beaumont Health System, located in Royal Oak, MI, USA, approved this research (HIC#: 2015-172). This study adhered to ethical standards by obtaining written informed consent from all participants and following the Helsinki Declaration principles. Rigorous matching criteria—such as age, gender, and body mass index (BMI)—were used to pair individuals with HS with healthy controls, ensuring the reliability and significance of the results.
Hidradenitis Suppurativa Sample Selection
Inclusion and exclusion criteria: HS patients were clinically evaluated for symptoms and severity by three independent board-certified dermatologists (RR, DGS, TM) at VS Hospital in Ahmedabad, India, using a questionnaire [96], based on the European Hidradenitis Suppurativa Foundation (EHSF) guidelines [97]. The diagnostic process and sample assessment followed these guidelines, with disease severity evaluated using established scoring systems, including the Hurley score [98], the HS Severity Score System (IHS4) [5], and the Autoinflammatory Disease Damage Index (ADDI) [99]. These systems were used to evaluate the severity and impact of the condition on those affected.
Inclusion criteria: Adult patients (>20 years) included in the study met the following criteria: (a) diagnosed with HS for more than 5 years, (b) had at least moderate Hurley II severity, (c) scored more than 3 points on the International Hidradenitis Suppurativa Severity Score System (IHS4), (d) scored less than 3 points on the Autoinflammatory Disease Damage Index (ADDI), (e) newly diagnosed (<3 months), or (f) untreated for the past 6 months. This ensured a comprehensive understanding by including patients with a confirmed HS diagnosis for over 5 years, representing the chronic phase of the disease, to investigate long-term molecular adaptations, including epigenetic modifications and sustained gene expression changes driven by chronic inflammation and pain. Additionally, the inclusion of patients newly diagnosed within the last three months allowed us to assess early epigenetic alterations before long-term changes occurred. These inclusion criteria encompass both chronic and early-stage disease dynamics, offering valuable insights into the spectrum of molecular changes in HS.
Exclusion criteria: The exclusion criteria were as follows: (a) syndromic HS defined by Van der Zee and Jemec clinical phenotypes, (b) smoking, (c) fasting regimens or specific diets other than an omnivore, (d) alcohol abuse (Alcohol Use Disorders Identification Test (AUDIT) > 7 points), (e) drug addiction, (f) use of concurrent medications, including contraceptives, and TRG (taste receptor gene) inducers such as grapefruits which inhibit CYP3A4 and alter drug metabolism and treatment outcomes in HS (and also affect inflammation, complicating the analysis of genetic and immune markers), (g) previous treatment for HS, (h) chronic inflammatory or infectious diseases, (i) history of cancer within the past 5 years, and (j) individuals unable to provide informed consent for any reason.
DNA extraction, bisulfite conversion: Blood samples were obtained from 24 individuals diagnosed with HS and 24 healthy controls. Genomic DNA was extracted using the Gentra Puregene® Blood Kit (Qiagen, Germany). DNA methylation patterns were analyzed after sodium bisulfite conversion of the extracted DNA, following the manufacturer’s protocol with the EZ 96-DNA Methylation Kit (Zymo Research, Irvine, CA, USA).
Illumina Infinium Methylation EPIC BeadChip: Genome-wide DNA methylation profiling was performed using the Infinium MethylationEPIC BeadChip (450K) array (Illumina Inc., San Diego, CA, USA) with bisulfite-treated genomic DNA, following the manufacturer’s protocol, as previously described in detail in a prior publication [100,101].
Statistical and bioinformatic analysis: The data were analyzed using the GenomeStudio methylation analysis software (1.9.0) from Illumina. Methylation levels (β-values) for each CpG site were calculated. Before analysis, CpG probes with missing β-values were excluded. Differential methylation was evaluated by comparing β-values for each CpG locus between HS and controls. To minimize confounding, we excluded probes linked to sex chromosomes and those containing SNPs, particularly those with dbSNP entries within 10 bp of the CpG site; as binding sites can influence interactions, only CpG targets with allele frequencies ≤ 0.05 were included for further analysis.
The most discriminating CpG sites were selected based on a preset cutoff criterion: FDR p < 0.05. For genes with multiple CpG sites, the site with the highest AUC ROC (≥0.75) was selected alongside the lowest p-value. The p-value for methylation differences between case and control groups at each locus was calculated as previously described [100,101]. Raw and FDR p-values corrected for multiple testing (Benjamini–Hochberg test) were calculated. The AUC for combinations of loci was calculated using the ‘R’ program “ROCR” package (v3.5.0).
Selection of Candidate Genes for Chronic Pain: Pain-associated genes were identified by utilizing a genetic database that consolidates data from genome-wide association studies (GWASs), gene expression profiles, and curated research focused on pain-related phenotypes [102,103]. We focused on genes with strong GWAS evidence linking them to pain perception or chronic pain and those documented in pain pathways, particularly in chronic pain management. Detailed information on these genes, including functions, pathways, and relevant SNPs or mutations, was extracted and cross-referenced with other databases to ensure accuracy and relevance, especially in the context of chronic pain and genetic diseases. Moreover, to identify genes potentially related to pain, relevant review articles were reviewed [104,105,106,107,108]. These articles offered crucial insights by compiling and analyzing existing studies, particularly those focused on genes tied to pain perception, chronic pain, and opioid use. Additionally, we utilized the Human Pain Genetics Database (https://diatchenko.lab.mcgill.ca/hpgdb/ (accessed on 25 August 2024)), a hand-curated resource compiling genetic associations with human pain phenotypes. Genes that overlapped between this database and the published literature were included only once. The complete list of genes used in these analyses is provided in Supplementary Table S2 (n = 897).
Heatmap: The heatmap was generated using the ‘ComplexHeatmap’ package (version 1.6.0) within R (version 3.2.2), designed to display the methylation patterns of CpG sites strongly associated with pain in HS. For this analysis, only genes exhibiting an AUC > 0.9 and FDR values ≤ 0.05 were included, ensuring that the displayed methylation differences were both statistically significant and highly discriminative between HS patients and controls. To assess the relationships and similarities among samples, hierarchical cluster analysis was performed. This analysis utilized Ward’s method, which minimizes the sum of squares of any two (hypothetical) clusters that can be formed at each. This method is particularly effective in identifying distinct groups by examining the methylation profiles of the CpG sites [109,110].
Principal component analysis: Principal component analysis (PCA) is widely utilized as a dimensionality reduction technique to improve data visualization and facilitate feature extraction by simplifying complex datasets. In this study, PCA was employed to pinpoint the key features responsible for the greatest variance across groups, which is particularly useful in fields such as epigenetics. The R function ‘prcomp’ was applied to calculate the principal components (PCs), and PC1, PC2, and PC3 were selected for visualization. A PCA distribution plot was created using the ‘ggplot2’ package in R”.
Protein–Protein Interaction Network: The 253 significantly differentially methylated CpG sites associated with HS were used to match corresponding genes against the STRING database (version 12.0, https://string-db.org/, accessed on 29 August 2024) to identify known and predicted protein–protein interactions. [111]. The interaction network was constructed using the full STRING network, which includes both functional and physical protein associations. Edges in the network were defined based on evidence, with line color indicating the type of interaction evidence. Interactions were considered from curated databases, experimentally determined interactions, and gene co-expression data, while sources such as text mining, neighborhood, gene fusion, and co-occurrence were excluded. A medium confidence threshold of 0.400 was applied to ensure the reliability of the interactions included in the analysis. This process resulted in 249 genes with validated interactions, which were further explored to understand their interaction dynamics.
Gene Ontology and KEGG Pathway Analysis: Gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses were performed on differently methylated genes. Both the GO and KEGG pathway analyses were conducted using the ‘clusterProfiler’ and ‘Pathview’ packages in R, respectively [112,113]. These tools facilitate the statistical analysis and visualization of functional profiles of genes, which are essential for interpreting the biological themes among gene clusters. This approach improves our understanding of genomic data and reveals altered pathways in HS.
To visually represent the KEGG pathway analysis results, we utilized the SRplot web server (http://www.bioinformatics.com.cn/SRplot (accessed on 25 August 2024)) to generate a Sankey diagram and a dot plot [114]. The Sankey diagram illustrates the flow and connection between enriched KEGG pathways and associated genes, providing an intuitive depiction of pathway relationships. The dot plot highlights the statistical significance and gene ratios of the enriched pathways, offering a concise overview of key insights from the analysis.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms26031039/s1.
Author Contributions
Conceptualization, methodology, software; validation; formal analysis; investigation; resources; data curation; writing—original draft preparation, U.R. (Uppala Radhakrishna), L.V.U., J.P. and G.D.; writing—review and editing, U.R (Uppala Radhakrishna), M.R.K., S.K.N., I.H., J.P., N.S., L.V.U., R.M.R., G.D., U.R. (Uppala Ratnamala); visualization, U.R. (Uppala Radhakrishna), G.D. and I.H.; supervision, Project administration and funding acquisition, U.R (Uppala Radhakrishna). All authors have read and agreed to the published version of the manuscript.
Funding
The authors acknowledge financial support for the research, authorship, and publication of this article. Funding for this study was provided by HS-USA (Michigan, USA) to Dr. Uppala Radhakrishna’s laboratory. The efforts of SKN were supported by the Oklahoma Medical Research Foundation (OMRF), Oklahoma City, OK 73104, USA, under grant number OMRF-9124.
Institutional Review Board Statement
Reviewed and approved by Beaumont Health System, Royal Oak, MI, USA, (HIC#: 2015-172).
Informed Consent Statement
Not applicable.
Data Availability Statement
The published article and its Supplementary Materials contain all the data generated during this study.
Acknowledgments
We thank the late Vix Kennedy for his support and encouragement for the study. We thank the patients for their cooperation in the study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Crowley, J.J.; Mekkes, J.R.; Zouboulis, C.C.; Scheinfeld, N.; Kimball, A.; Sundaram, M.; Gu, Y.; Okun, M.M.; Kerdel, F. Association of hidradenitis suppurativa disease severity with increased risk for systemic comorbidities. Br. J. Dermatol. 2014, 171, 1561–1565. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T. Hidradenitis Suppurativa as a Potential Subtype of Autoinflammatory Keratinization Disease. Front. Immunol. 2020, 11, 847. [Google Scholar] [CrossRef] [PubMed]
- Metpally, R.P.; Vishweswaraiah, S.; Krishnamurthy, S.; Saiyed, N.; Stahl, R.C.; Golden, A.; Denisenko, A.; Staples, J.; Gonzaga-Jauregui, C.; Carey, D.J.; et al. Identification of Novel Genetic Risk Variants Associated with Hidradenitis Suppurativa in an Exome Sequencing Cohort of 92,455 Individuals. Dermatology 2024, 240, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Ratnamala, U.; Jain, N.K.; Jhala, D.D.; Prasad, P.V.S.; Saiyed, N.; Nair, S.; Radhakrishna, U. An Updated Mutation Spectrum of the gamma-Secretase Complex: Novel NCSTN Gene Mutation in an Indian Family with Hidradenitis Suppurativa and Acne Conglobata. Indian. J. Dermatol. 2023, 68, 141–147. [Google Scholar] [CrossRef]
- Radhakrishna, U.; Ratnamala, U.; Jhala, D.D.; Vadsaria, N.; Patel, M.; Uppala, L.V.; Vishweswaraiah, S.; Vedangi, A.; Saiyed, N.; Damiani, G.; et al. Methylated miRNAs may serve as potential biomarkers and therapeutic targets for hidradenitis suppurativa. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 2199–2213. [Google Scholar] [CrossRef]
- Radhakrishna, U.; Ratnamala, U.; Jhala, D.D.; Uppala, L.V.; Vedangi, A.; Saiyed, N.; Patel, M.; Shah, S.R.; Rawal, R.M.; Jemec, G.B.E.; et al. Deregulated Long Non-Coding RNAs (lncRNA) as Promising Biomarkers in Hidradenitis Suppurativa. J. Clin. Med. 2024, 13, 3016. [Google Scholar] [CrossRef]
- Ratnamala, U.; Jhala, D.; Jain, N.K.; Saiyed, N.M.; Raveendrababu, M.; Rao, M.V.; Mehta, T.Y.; Al-Ali, F.M.; Raval, K.; Nair, S.; et al. Expanding the spectrum of gamma-secretase gene mutation-associated phenotypes: Two novel mutations segregating with familial hidradenitis suppurativa (acne inversa) and acne conglobata. Exp. Dermatol. 2016, 25, 314–316. [Google Scholar] [CrossRef]
- Saunte, D.M.; Boer, J.; Stratigos, A.; Szepietowski, J.C.; Hamzavi, I.; Kim, K.H.; Zarchi, K.; Antoniou, C.; Matusiak, L.; Lim, H.W.; et al. Diagnostic delay in hidradenitis suppurativa is a global problem. Br. J. Dermatol. 2015, 173, 1546–1549. [Google Scholar] [CrossRef]
- Pelekanou, A.; Kanni, T.; Savva, A.; Mouktaroudi, M.; Raftogiannis, M.; Kotsaki, A.; Giamarellos-Bourboulis, E.J. Long-term efficacy of etanercept in hidradenitis suppurativa: Results from an open-label phase II prospective trial. Exp. Dermatol. 2010, 19, 538–540. [Google Scholar] [CrossRef]
- Vinkel, C.; Thomsen, S.F. Hidradenitis Suppurativa: Causes, Features, and Current Treatments. J. Clin. Aesthet. Dermatol. 2018, 11, 17–23. [Google Scholar]
- Okun, M.M.; Flamm, A.; Werley, E.B.; Kirby, J.S. Hidradenitis Suppurativa: Diagnosis and Management in the Emergency Department. J. Emerg. Med. 2022, 63, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Alavi, A.; Farzanfar, D.; Lee, R.K.; Almutairi, D. The Contribution of Malodour in Quality of Life of Patients With Hidradenitis Suppurativa. J. Cutan. Med. Surg. 2018, 22, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Savage, K.T.; Singh, V.; Patel, Z.S.; Yannuzzi, C.A.; McKenzie-Brown, A.M.; Lowes, M.A.; Orenstein, L.A.V. Pain management in hidradenitis suppurativa and a proposed treatment algorithm. J. Am. Acad. Dermatol. 2021, 85, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Ballard, K.; Shuman, V.L. Hidradenitis Suppurativa. In StatPearls; Treasure Island (FL) Ineligible Companies: Treasure Island, FL, USA, 2023. [Google Scholar]
- Kaaz, K.; Szepietowski, J.C.; Matusiak, L. Influence of Itch and Pain on Sleep Quality in Patients with Hidradenitis Suppurativa. Acta Derm. Venereol. 2018, 98, 757–761. [Google Scholar] [CrossRef]
- Alotaibi, H.M. Incidence, Risk Factors, and Prognosis of Hidradenitis Suppurativa Across the Globe: Insights from the Literature. Clin. Cosmet. Investig. Dermatol. 2023, 16, 545–552. [Google Scholar] [CrossRef]
- Amtmann, D.; Askew, R.L.; Kim, J.; Chung, H.; Ehde, D.M.; Bombardier, C.H.; Kraft, G.H.; Jones, S.M.; Johnson, K.L. Pain affects depression through anxiety, fatigue, and sleep in multiple sclerosis. Rehabil. Psychol. 2015, 60, 81–90. [Google Scholar] [CrossRef]
- Brasure, M.; Nelson, V.A.; Scheiner, S.; Forte, M.L.; Butler, M.; Nagarkar, S.; Saha, J.; Wilt, T.J. Treatment for Acute Pain: An Evidence Map; Agency for Healthcare Research and Quality: Rockville, MD, USA, 2019.
- Patel, Z.S.; Hoffman, L.K.; Buse, D.C.; Grinberg, A.S.; Afifi, L.; Cohen, S.R.; Lowes, M.A.; Seng, E.K. Pain, Psychological Comorbidities, Disability, and Impaired Quality of Life in Hidradenitis Suppurativa [corrected]. Curr. Pain. Headache Rep. 2017, 21, 49. [Google Scholar] [CrossRef]
- Matusiak, L. Profound consequences of hidradenitis suppurativa: A review. Br. J. Dermatol. 2020, 183, e171–e177. [Google Scholar] [CrossRef]
- Moller Johansen, L.; Gerra, M.C.; Arendt-Nielsen, L. Time course of DNA methylation in pain conditions: From experimental models to humans. Eur. J. Pain. 2021, 25, 296–312. [Google Scholar] [CrossRef]
- Krajewski, P.K.; Matusiak, L.; von Stebut, E.; Schultheis, M.; Kirschner, U.; Nikolakis, G.; Szepietowski, J.C. Pain in Hidradenitis Suppurativa: A Cross-sectional Study of 1795 Patients. Acta Derm. Venereol. 2021, 101, adv00364. [Google Scholar]
- Whitley, S.K. Elucidation of Pain Mechanisms in Hidradenitis Suppurativa. JAMA Dermatol. 2023, 159, 1047–1049. [Google Scholar] [CrossRef] [PubMed]
- Horváth, B.; Janse, I.C.; Sibbald, G.R. Pain management in patients with hidradenitis suppurativa. J. Am. Acad. Dermatol. 2015, 73, S47–S51. [Google Scholar] [CrossRef] [PubMed]
- Amat-Samaranch, V.; Agut-Busquet, E.; Vilarrasa, E.; Puig, L. New perspectives on the treatment of hidradenitis suppurativa. Ther. Adv. Chronic Dis. 2021, 12, 20406223211055920. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yao, J.; Rong, M. Editorial: Role of Ion Channels in Pain. Front. Pharmacol. 2022, 13, 884665. [Google Scholar] [CrossRef]
- Alles, S.R.A.; Gomez, K.; Moutal, A.; Khanna, R. Putative roles of SLC7A5 (LAT1) transporter in pain. Neurobiol. Pain. 2020, 8, 100050. [Google Scholar] [CrossRef]
- Radhakrishna, U.; Ratnamala, U.; Jhala, D.D.; Uppala, L.V.; Vedangi, A.; Saiyed, N.; Patel, M.; Vadsaria, N.; Shah, S.R.; Rawal, R.M.; et al. Hidradenitis suppurativa associated telomere-methylome dysregulations in blood. J. Eur. Acad. Dermatol. Venereol. 2024, 38, 393–403. [Google Scholar] [CrossRef]
- Jha, M.K.; Song, G.J.; Lee, M.G.; Jeoung, N.H.; Go, Y.; Harris, R.A.; Park, D.H.; Kook, H.; Lee, I.K.; Suk, K. Metabolic Connection of Inflammatory Pain: Pivotal Role of a Pyruvate Dehydrogenase Kinase-Pyruvate Dehydrogenase-Lactic Acid Axis. J. Neurosci. 2015, 35, 14353–14369. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef]
- Radhakrishna, U.; Ratnamala, U.; Jhala, D.D.; Uppala, L.V.; Vedangi, A.; Patel, M.; Vadsaria, N.; Shah, S.; Saiyed, N.; Rawal, R.M.; et al. Hidradenitis suppurativa presents a methylome dysregulation capable to explain the pro-inflammatory microenvironment: Are these DNA methylations potential therapeutic targets? J. Eur. Acad. Dermatol. Venereol. 2023, 37, 2109–2123. [Google Scholar] [CrossRef]
- Bumgarner, J.R.; McCray, E.W.; Nelson, R.J. The disruptive relationship among circadian rhythms, pain, and opioids. Front. Neurosci. 2023, 17, 1109480. [Google Scholar] [CrossRef]
- Stavropoulou, E.; Pircalabioru, G.G.; Bezirtzoglou, E. The Role of Cytochromes P450 in Infection. Front. Immunol. 2018, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishna, U.; Ratnamala, U.; Jhala, D.D.; Vadsaria, N.; Patel, M.; Uppala, L.V.; Vedangi, A.; Saiyed, N.; Rawal, R.M.; Damiani, G.; et al. Cytochrome P450 Genes Mediated by DNA Methylation Are Involved in the Resistance to Hidradenitis Suppurativa. J. Investig. Dermatol. 2023, 143, 670–673.e19. [Google Scholar] [CrossRef] [PubMed]
- Goodin, B.R.; Overstreet, D.S.; Penn, T.M.; Bakshi, R.; Quinn, T.L.; Sims, A.; Ptacek, T.; Jackson, P.; Long, D.L.; Aroke, E.N. Epigenome-wide DNA methylation profiling of conditioned pain modulation in individuals with non-specific chronic low back pain. Clin. Epigenet. 2022, 14, 45. [Google Scholar] [CrossRef] [PubMed]
- Tajerian, M.; Alvarado, S.; Millecamps, M.; Dashwood, T.; Anderson, K.M.; Haglund, L.; Ouellet, J.; Szyf, M.; Stone, L.S. DNA methylation of SPARC and chronic low back pain. Mol. Pain. 2011, 7, 65. [Google Scholar] [CrossRef]
- Garriga, J.; Laumet, G.; Chen, S.R.; Zhang, Y.; Madzo, J.; Issa, J.J.; Pan, H.L.; Jelinek, J. Nerve Injury-Induced Chronic Pain Is Associated with Persistent DNA Methylation Reprogramming in Dorsal Root Ganglion. J. Neurosci. 2018, 38, 6090–6101. [Google Scholar] [CrossRef]
- Zheng, G.; Ren, J.; Shang, L.; Bao, Y. Role of autophagy in the pathogenesis and regulation of pain. Eur. J. Pharmacol. 2023, 955, 175859. [Google Scholar] [CrossRef]
- Merighi, A. Brain-Derived Neurotrophic Factor, Nociception, and Pain. Biomolecules 2024, 14, 539. [Google Scholar] [CrossRef]
- Kozera, E.K.; Lowes, M.A.; Hsiao, J.L.; Frew, J.W. Clinical considerations in the management of hidradenitis suppurativa in women. Int. J. Womens Dermatol. 2021, 7 5Pt B, 664–671. [Google Scholar] [CrossRef]
- Yamamoto, S.; Kashimoto, R.; Furukawa, S.; Ohashi, A.; Satoh, A. Lmx1b activation in axolotl limb regeneration. Dev. Dyn. 2022, 251, 1509–1523. [Google Scholar] [CrossRef]
- Warner, S.C.; van Meurs, J.B.; Schiphof, D.; Bierma-Zeinstra, S.M.; Hofman, A.; Uitterlinden, A.G.; Richardson, H.; Jenkins, W.; Doherty, M.; Valdes, A.M. Genome-wide association scan of neuropathic pain symptoms post total joint replacement highlights a variant in the protein-kinase C gene. Eur. J. Hum. Genet. 2017, 25, 446–451. [Google Scholar] [CrossRef]
- Lazarus, M.B.; Novotny, C.J.; Shokat, K.M. Structure of the human autophagy initiating kinase ULK1 in complex with potent inhibitors. ACS Chem. Biol. 2015, 10, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Steward, A.M.; Morgan, J.D.; Espinosa, J.P.; Turk, D.C.; Patel, K.V. Chronic Pain and Telomere Length in Community-Dwelling Adults: Findings From the 1999 to 2002 National Health and Nutrition Examination Survey. J. Pain 2017, 18, 1517–1525. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.; Monteiro, C.; Cardoso-Cruz, H.; Galhardo, V. Altered Brain Expression of DNA Methylation and Hydroxymethylation Epigenetic Enzymes in a Rat Model of Neuropathic Pain. Int. J. Mol. Sci. 2023, 24, 7305. [Google Scholar] [CrossRef]
- Coppede, F.; Stoccoro, A.; Tannorella, P.; Migliore, L. Plasma Homocysteine and Polymorphisms of Genes Involved in Folate Metabolism Correlate with DNMT1 Gene Methylation Levels. Metabolites 2019, 9, 298. [Google Scholar] [CrossRef] [PubMed]
- Buesing, S.; Costa, M.; Schilling, J.M.; Moeller-Bertram, T. Vitamin B12 as a Treatment for Pain. Pain. Physician 2019, 22, E45–E52. [Google Scholar] [CrossRef]
- Sutherland, H.G.; Albury, C.L.; Griffiths, L.R. Advances in genetics of migraine. J. Headache Pain. 2019, 20, 72. [Google Scholar] [CrossRef]
- Deng, Z.; Fan, T.; Xiao, C.; Tian, H.; Zheng, Y.; Li, C.; He, J. TGF-beta signaling in health, disease, and therapeutics. Signal Transduct. Target. Ther. 2024, 9, 61. [Google Scholar] [CrossRef]
- Griesius, S.; O’Donnell, C.; Waldron, S.; Thomas, K.L.; Dwyer, D.M.; Wilkinson, L.S.; Hall, J.; Robinson, E.S.J.; Mellor, J.R. Reduced expression of the psychiatric risk gene DLG2 (PSD93) impairs hippocampal synaptic integration and plasticity. Neuropsychopharmacology 2022, 47, 1367–1378. [Google Scholar] [CrossRef]
- Daguet, I.; Raverot, V.; Bouhassira, D.; Gronfier, C. Circadian rhythmicity of pain sensitivity in humans. Brain 2022, 145, 3225–3235. [Google Scholar] [CrossRef]
- Radhakrishna, U.; Ratnamala, U.; Jhala, D.D.; Uppala, L.V.; Vedangi, A.; Saiyed, N.; Shah, S.R.; Patel, M.; Rawal, R.M.; Mazza, T.; et al. DNA methylation patterns of circadian and ultradian genes are altered in the peripheral blood of patients with hidradenitis suppurativa. Front. Immunol. 2024, 15, 1475424. [Google Scholar] [CrossRef]
- Callaghan, B.; Feldman, E. The metabolic syndrome and neuropathy: Therapeutic challenges and opportunities. Ann. Neurol. 2013, 74, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Baik, J.H. Dopamine signaling in food addiction: Role of dopamine D2 receptors. BMB Rep. 2013, 46, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Ye, D.; Fairchild, T.J.; Vo, L.; Drummond, P.D. High Blood Glucose and Excess Body fat Enhance Pain Sensitivity and Weaken Pain Inhibition in Healthy Adults: A Single-blind Cross-over Randomized Controlled Trial. J. Pain 2023, 24, 128–144. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Suarez, V.J.; Beltran-Velasco, A.I.; Redondo-Florez, L.; Martin-Rodriguez, A.; Yanez-Sepulveda, R.; Tornero-Aguilera, J.F. Neuro-Vulnerability in Energy Metabolism Regulation: A Comprehensive Narrative Review. Nutrients 2023, 15, 3106. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.Y.; Jones, B.L.; Haas, G.L. Suicidal ideation and aggression in childhood, genetic variation and young adult depression. J. Affect. Disord. 2020, 276, 954–962. [Google Scholar] [CrossRef]
- Saez, E.; Erkoreka, L.; Moreno-Calle, T.; Berjano, B.; Gonzalez-Pinto, A.; Basterreche, N.; Arrue, A. Genetic variables of the glutamatergic system associated with treatment-resistant depression: A review of the literature. World J. Psychiatry 2022, 12, 884–896. [Google Scholar] [CrossRef]
- Rafiei, D.; Kolla, N.J. Elevated Brain Fatty Acid Amide Hydrolase Induces Depressive-Like Phenotypes in Rodent Models: A Review. Int. J. Mol. Sci. 2021, 22, 1047. [Google Scholar] [CrossRef]
- Lopez, J.F.; Palkovits, M.; Arato, M.; Mansour, A.; Akil, H.; Watson, S.J. Localization and quantification of pro-opiomelanocortin mRNA and glucocorticoid receptor mRNA in pituitaries of suicide victims. Neuroendocrinology 1992, 56, 491–501. [Google Scholar] [CrossRef]
- Gonzalez-Castro, T.B.; Genis-Mendoza, A.D.; Tovilla-Zarate, C.A.; Juarez-Rojop, I.E.; Lopez-Narvaez, M.L.; Perez-Hernandez, N.; Rodriguez-Perez, J.M.; Martinez-Magana, J.J. Association between polymorphisms of NOS1, NOS2 and NOS3 genes and suicide behavior: A systematic review and meta-analysis. Metab. Brain Dis. 2019, 34, 967–977. [Google Scholar] [CrossRef]
- Lee, H.Y.; Kim, Y.K. Transforming growth factor-beta1 and major depressive disorder with and without attempted suicide: Preliminary study. Psychiatry Res. 2010, 178, 92–96. [Google Scholar] [CrossRef]
- Bonsignore, G.; Martinotti, S.; Ranzato, E. Wound Repair and Ca2+ Signalling Interplay: The Role of Ca2+ Channels in Skin. Cells 2024, 13, 491. [Google Scholar] [CrossRef] [PubMed]
- Andrade, A.; Brennecke, A.; Mallat, S.; Brown, J.; Gomez-Rivadeneira, J.; Czepiel, N.; Londrigan, L. Genetic Associations between Voltage-Gated Calcium Channels and Psychiatric Disorders. Int. J. Mol. Sci. 2019, 20, 3537. [Google Scholar] [CrossRef] [PubMed]
- Nissenbaum, J. From mouse to humans: Discovery of the CACNG2 pain susceptibility gene. Clin. Genet. 2012, 82, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Bellessort, B.; Bachelot, A.; Grouthier, V.; De Lombares, C.; Narboux-Neme, N.; Garagnani, P.; Pirazzini, C.; Astigiano, S.; Mastracci, L.; Fontaine, A.; et al. Comparative analysis of molecular signatures suggests the use of gabapentin for the management of endometriosis-associated pain. J. Pain. Res. 2018, 11, 715–725. [Google Scholar] [CrossRef]
- Duan, J.; Grando, C.; Liu, S.; Chernyavsky, A.; Chen, J.K.; Andersen, B.; Grando, S.A. The M3 Muscarinic Acetylcholine Receptor Promotes Epidermal Differentiation. J. Investig. Dermatol. 2022, 142, 3211–3221.e2. [Google Scholar] [CrossRef]
- Kowalska, M.; Kapelusiak-Pielok, M.; Grzelak, T.; Wypasek, E.; Kozubski, W.; Dorszewska, J. The New *G29A and G1222A of HCRTR1, 5-HTTLPR of SLC6A4 Polymorphisms and Hypocretin-1, Serotonin Concentrations in Migraine Patients. Front. Mol. Neurosci. 2018, 11, 191. [Google Scholar] [CrossRef]
- Bardin, L. The complex role of serotonin and 5-HT receptors in chronic pain. Behav. Pharmacol. 2011, 22, 390–404. [Google Scholar] [CrossRef]
- Echeverria, F.; Gonzalez-Sanabria, N.; Alvarado-Sanchez, R.; Fernandez, M.; Castillo, K.; Latorre, R. Large conductance voltage-and calcium-activated K+ (BK) channel in health and disease. Front. Pharmacol. 2024, 15, 1373507. [Google Scholar] [CrossRef]
- Wulff, H.; Castle, N.A.; Pardo, L.A. Voltage-gated potassium channels as therapeutic targets. Nat. Rev. Drug Discov. 2009, 8, 982–1001. [Google Scholar] [CrossRef]
- Du, X.; Gao, H.; Jaffe, D.; Zhang, H.; Gamper, N. M-type K+ channels in peripheral nociceptive pathways. Br. J. Pharmacol. 2018, 175, 2158–2172. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Wen, J.; Yang, W.; Wang, C.; Gao, L.; Zheng, L.H.; Wang, T.; Ran, K.; Li, Y.; Li, X.; et al. Gain-of-function mutations in SCN11A cause familial episodic pain. Am. J. Hum. Genet. 2013, 93, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Leipold, E.; Hanson-Kahn, A.; Frick, M.; Gong, P.; Bernstein, J.A.; Voigt, M.; Katona, I.; Oliver Goral, R.; Altmuller, J.; Nurnberg, P.; et al. Cold-aggravated pain in humans caused by a hyperactive NaV1.9 channel mutant. Nat. Commun. 2015, 6, 10049. [Google Scholar] [CrossRef] [PubMed]
- Vanoye, C.G.; Gurnett, C.A.; Holland, K.D.; George, A.L., Jr.; Kearney, J.A. Novel SCN3A variants associated with focal epilepsy in children. Neurobiol. Dis. 2014, 62, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Veerman, C.C.; Wilde, A.A.; Lodder, E.M. The cardiac sodium channel gene SCN5A and its gene product NaV1.5: Role in physiology and pathophysiology. Gene 2015, 573, 177–187. [Google Scholar] [CrossRef]
- O’Brien, J.E.; Meisler, M.H. Sodium channel SCN8A (Nav1.6): Properties and de novo mutations in epileptic encephalopathy and intellectual disability. Front. Genet. 2013, 4, 213. [Google Scholar] [CrossRef]
- Duan, Y.; Gong, K.; Xu, S.; Zhang, F.; Meng, X.; Han, J. Regulation of cholesterol homeostasis in health and diseases: From mechanisms to targeted therapeutics. Signal Transduct. Target. Ther. 2022, 7, 265. [Google Scholar] [CrossRef]
- Tang, B.L. The Expanding Therapeutic Potential of Neuronal KCC2. Cells 2020, 9, 240. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Yue, G.; Zhao, Y. Energy metabolism disturbance in migraine: From a mitochondrial point of view. Front. Physiol. 2023, 14, 1133528. [Google Scholar] [CrossRef]
- Barker, J.M.; Taylor, J.R.; De Vries, T.J.; Peters, J. Brain-derived neurotrophic factor and addiction: Pathological versus therapeutic effects on drug seeking. Brain Res. 2015, 1628 Pt A, 68–81. [Google Scholar] [CrossRef]
- Blum, K.; Oscar-Berman, M.; Barh, D.; Giordano, J.; Gold, M. Dopamine Genetics and Function in Food and Substance Abuse. J. Genet. Syndr. Gene Ther. 2013, 4, 1000121. [Google Scholar]
- Navarrete, F.; Garcia-Gutierrez, M.S.; Gasparyan, A.; Navarro, D.; Lopez-Picon, F.; Morcuende, A.; Femenia, T.; Manzanares, J. Biomarkers of the Endocannabinoid System in Substance Use Disorders. Biomolecules 2022, 12, 396. [Google Scholar] [CrossRef] [PubMed]
- Panlilio, L.V.; Justinova, Z.; Goldberg, S.R. Inhibition of FAAH and activation of PPAR: New approaches to the treatment of cognitive dysfunction and drug addiction. Pharmacol. Ther. 2013, 138, 84–102. [Google Scholar] [CrossRef] [PubMed]
- Nudmamud-Thanoi, S.; Veerasakul, S.; Thanoi, S. Pharmacogenetics of drug dependence: Polymorphisms of genes involved in GABA neurotransmission. Neurosci. Lett. 2020, 726, 134463. [Google Scholar] [CrossRef] [PubMed]
- Popescu, A.; Marian, M.; Dragoi, A.M.; Costea, R.V. Understanding the genetics and neurobiological pathways behind addiction (Review). Exp. Ther. Med. 2021, 21, 544. [Google Scholar] [CrossRef]
- Thakkar, B.; Acevedo, E.O. BDNF as a biomarker for neuropathic pain: Consideration of mechanisms of action and associated measurement challenges. Brain Behav. 2023, 13, e2903. [Google Scholar] [CrossRef]
- Anthoni, M.; Fyhrquist-Vanni, N.; Wolff, H.; Alenius, H.; Lauerma, A. Transforming growth factor-beta/Smad3 signalling regulates inflammatory responses in a murine model of contact hypersensitivity. Br. J. Dermatol. 2008, 159, 546–554. [Google Scholar]
- Eve, M.; Gandawijaya, J.; Yang, L.; Oguro-Ando, A. Neuronal Cell Adhesion Molecules May Mediate Neuroinflammation in Autism Spectrum Disorder. Front. Psychiatry 2022, 13, 842755. [Google Scholar] [CrossRef]
- Iemmolo, M.; Ghersi, G.; Bivona, G. The Cytokine CX3CL1 and ADAMs/MMPs in Concerted Cross-Talk Influencing Neurodegenerative Diseases. Int. J. Mol. Sci. 2023, 24, 8026. [Google Scholar] [CrossRef]
- Harno, E.; Gali Ramamoorthy, T.; Coll, A.P.; White, A. POMC: The Physiological Power of Hormone Processing. Physiol. Rev. 2018, 98, 2381–2430. [Google Scholar] [CrossRef]
- Cohen, B.; Cadesky, A.; Jaggi, S. Dermatologic manifestations of thyroid disease: A literature review. Front. Endocrinol. 2023, 14, 1167890. [Google Scholar] [CrossRef]
- Lightman, S.L.; Birnie, M.T.; Conway-Campbell, B.L. Dynamics of ACTH and Cortisol Secretion and Implications for Disease. Endocrine Reviews 2020, 41, 470–490. [Google Scholar] [CrossRef] [PubMed]
- Kramer, P.R.; Rao, M.; Stinson, C.; Bellinger, L.L.; Kinchington, P.R.; Yee, M.B. Aromatase Derived Estradiol Within the Thalamus Modulates Pain Induced by Varicella Zoster Virus. Front. Integr. Neurosci. 2018, 12, 46. [Google Scholar] [CrossRef] [PubMed]
- Husby, A. On the Use of Blood Samples for Measuring DNA Methylation in Ecological Epigenetic Studies. Integr. Comp. Biol. 2020, 60, 1558–1566. [Google Scholar] [CrossRef] [PubMed]
- Cazzaniga, S.; Naldi, L.; Damiani, G.; Atzori, L.; Patta, F.; Guidarelli, G.; Bettoli, V. Validation of a visual-aided questionnaire for the self-assessment of hidradenitits suppurativa. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 1993–1998. [Google Scholar] [CrossRef]
- Lipsker, D.; Severac, F.; Freysz, M.; Sauleau, E.; Boer, J.; Emtestam, L.; Matusiak, L.; Prens, E.; Velter, C.; Lenormand, C.; et al. The ABC of Hidradenitis Suppurativa: A Validated Glossary on how to Name Lesions. Dermatology 2016, 232, 137–142. [Google Scholar] [CrossRef]
- Hurley, H.J. Axillary hyperhidrosis, apocrine bromhidrosis, hidradenitis suppurativa, and familial benign pemphigus: Surgical approach. In Roenigk and Roenigk’s Dermatologic Surgery, Principles and Practice, 2nd ed.; Roenigk, R.K., Roenigk, H.H., Jr., Eds.; Marcel Dekker: New York, NY, USA, 1996; pp. 623–645. [Google Scholar]
- Damiani, G.; Della Valle, V.; Iannone, M.; Dini, V.; Marzano, A.V. Autoinflammatory Disease Damage Index (ADDI): A possible newborn also in hidradenitis suppurativa daily practice. Ann. Rheum. Dis. 2017, 76, e25. [Google Scholar] [CrossRef]
- Radhakrishna, U.; Vishweswaraiah, S.; Uppala, L.V.; Szymanska, M.; Macknis, J.; Kumar, S.; Saleem-Rasheed, F.; Aydas, B.; Forray, A.; Muvvala, S.B.; et al. Placental DNA methylation profiles in opioid-exposed pregnancies and associations with the neonatal opioid withdrawal syndrome. Genomics 2021, 113, 1127–1135. [Google Scholar] [CrossRef]
- Radhakrishna, U.; Albayrak, S.; Zafra, R.; Baraa, A.; Vishweswaraiah, S.; Veerappa, A.M.; Mahishi, D.; Saiyed, N.; Mishra, N.K.; Guda, C.; et al. Placental epigenetics for evaluation of fetal congenital heart defects: Ventricular Septal Defect (VSD). PLoS ONE 2019, 14, e0200229. [Google Scholar] [CrossRef]
- Parisien, M.; Khoury, S.; Chabot-Dore, A.J.; Sotocinal, S.G.; Slade, G.D.; Smith, S.B.; Fillingim, R.B.; Ohrbach, R.; Greenspan, J.D.; Maixner, W.; et al. Effect of Human Genetic Variability on Gene Expression in Dorsal Root Ganglia and Association with Pain Phenotypes. Cell Rep. 2017, 19, 1940–1952. [Google Scholar] [CrossRef]
- James, S. Human pain and genetics: Some basics. Br. J. Pain. 2013, 7, 171–178. [Google Scholar] [CrossRef]
- Bai, G.; Ren, K.; Dubner, R. Epigenetic regulation of persistent pain. Transl. Res. 2015, 165, 177–199. [Google Scholar] [CrossRef] [PubMed]
- Linnstaedt, S.D.; Rueckeis, C.A.; Riker, K.D.; Pan, Y.; Wu, A.; Yu, S.; Wanstrath, B.; Gonzalez, M.; Harmon, E.; Green, P.; et al. MicroRNA-19b predicts widespread pain and posttraumatic stress symptom risk in a sex-dependent manner following trauma exposure. Pain 2020, 161, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Zorina-Lichtenwalter, K.; Meloto, C.B.; Khoury, S.; Diatchenko, L. Genetic predictors of human chronic pain conditions. Neuroscience 2016, 338, 36–62. [Google Scholar] [CrossRef] [PubMed]
- Wistrom, E.; Chase, R.; Smith, P.R.; Campbell, Z.T. A compendium of validated pain genes. WIREs Mech. Dis. 2022, 14, e1570. [Google Scholar] [CrossRef]
- Sexton, J.E.; Cox, J.J.; Zhao, J.; Wood, J.N. The Genetics of Pain: Implications for Therapeutics. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 123–142. [Google Scholar] [CrossRef]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Gu, Z. Complex heatmap visualization. Imeta 2022, 1, e43. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res 2023, 51, D638–D646. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 2013, 29, 1830–1831. [Google Scholar] [CrossRef]
- Tang, D.; Chen, M.; Huang, X.; Zhang, G.; Zeng, L.; Zhang, G.; Wu, S.; Wang, Y. SRplot: A free online platform for data visualization and graphing. PLoS ONE 2023, 18, e0294236. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).