
Rno-miR-130b Attenuates Lipid Accumulation Through Promoting Apoptosis and Inhibiting Differentiation in Rat Intramuscular Adipocytes


Abstract
:1. Introduction
2. Results
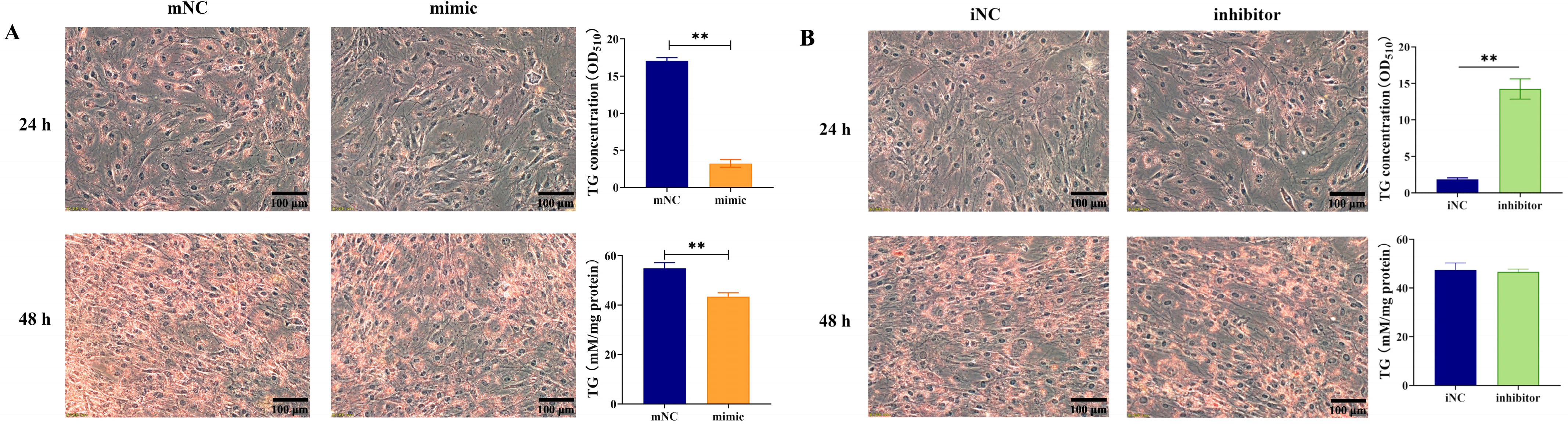
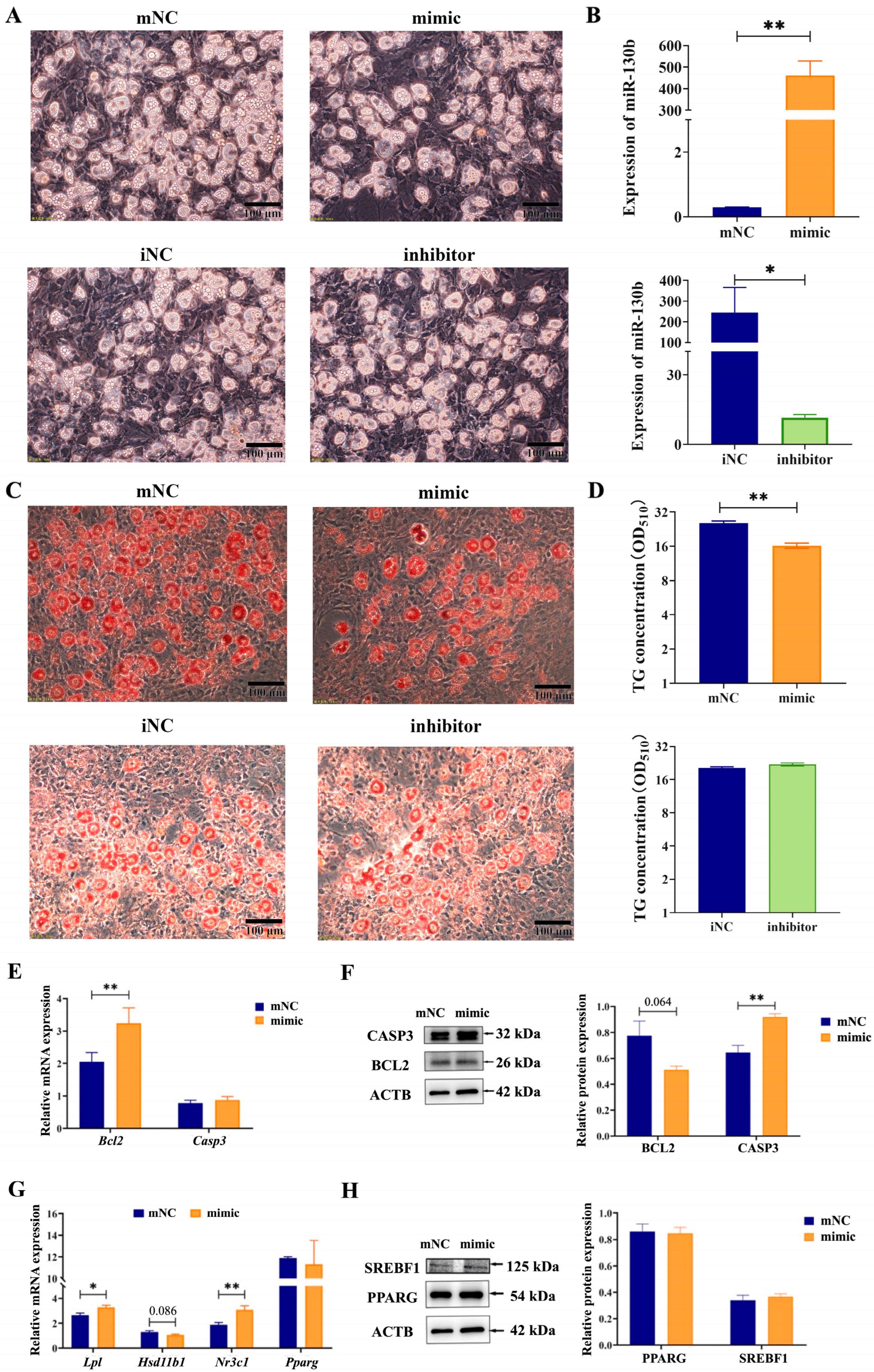
2.1. Overexpression of MiR-130b Significantly Inhibited Lipid Deposition in Rat IMAs
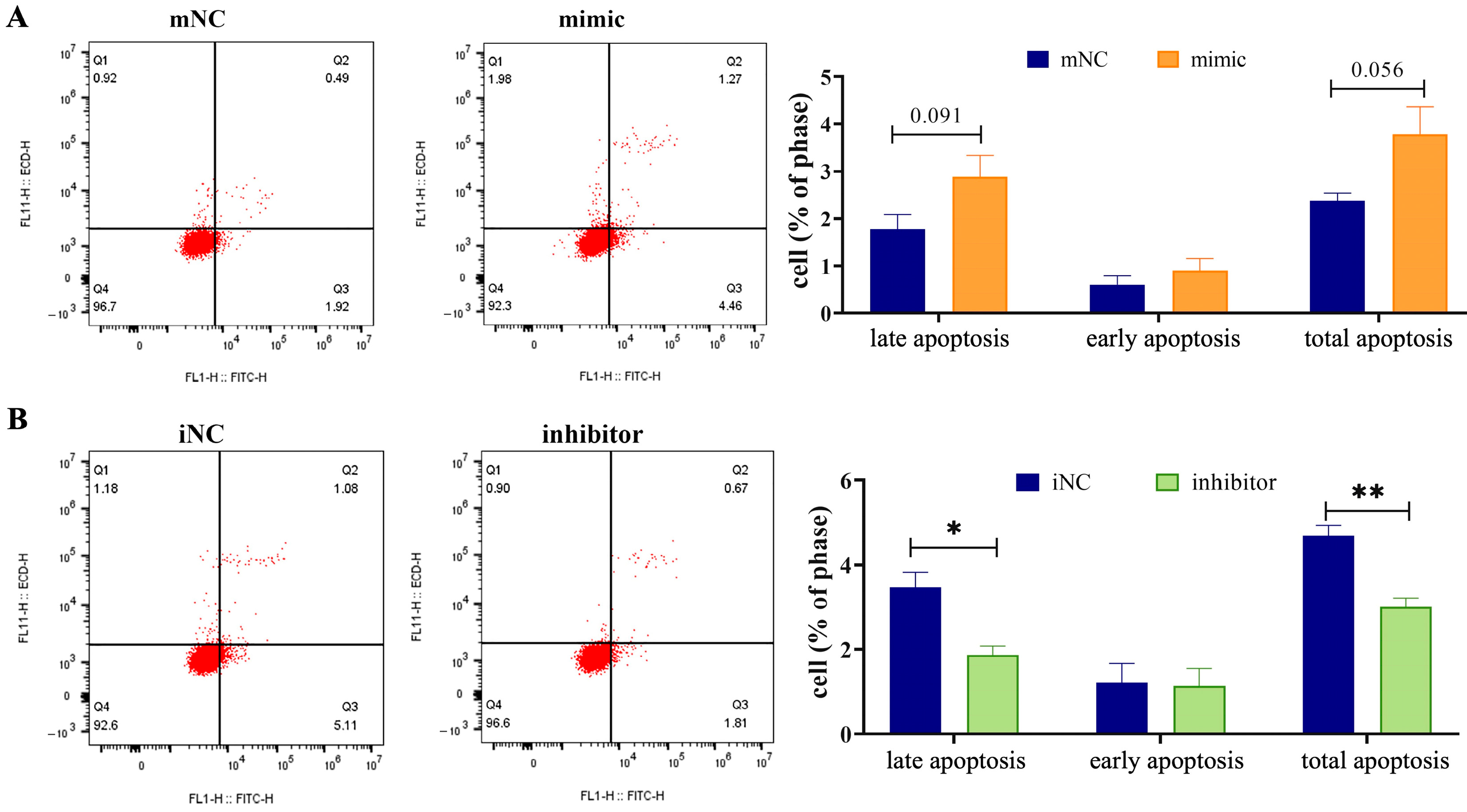
2.2. Overexpression of MiR-130b Significantly Promoted the Apoptosis of Rat IMAs
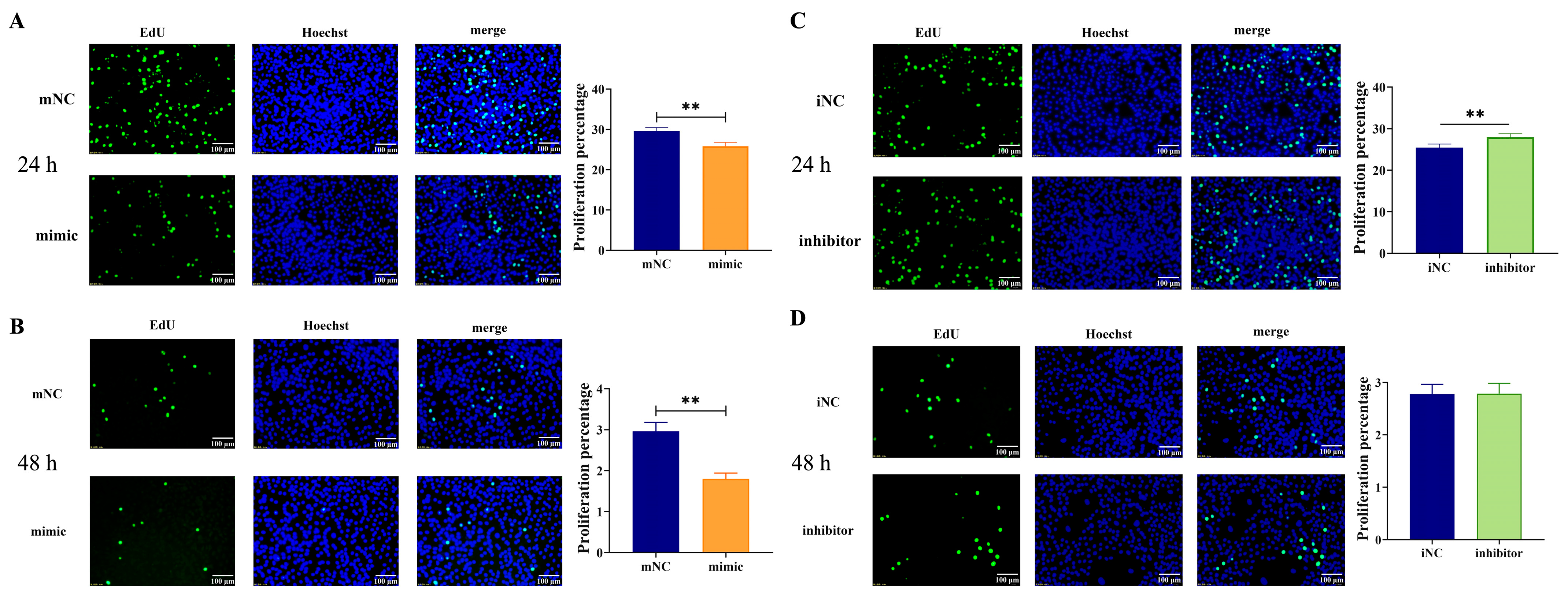
2.3. Overexpression of MiR-130b Significantly Inhibited the Proliferation of Rat IMAs
2.4. Overexpression of MiR-130b Changed Cell Cycle Phases at Different Time Points in Rat IMAs
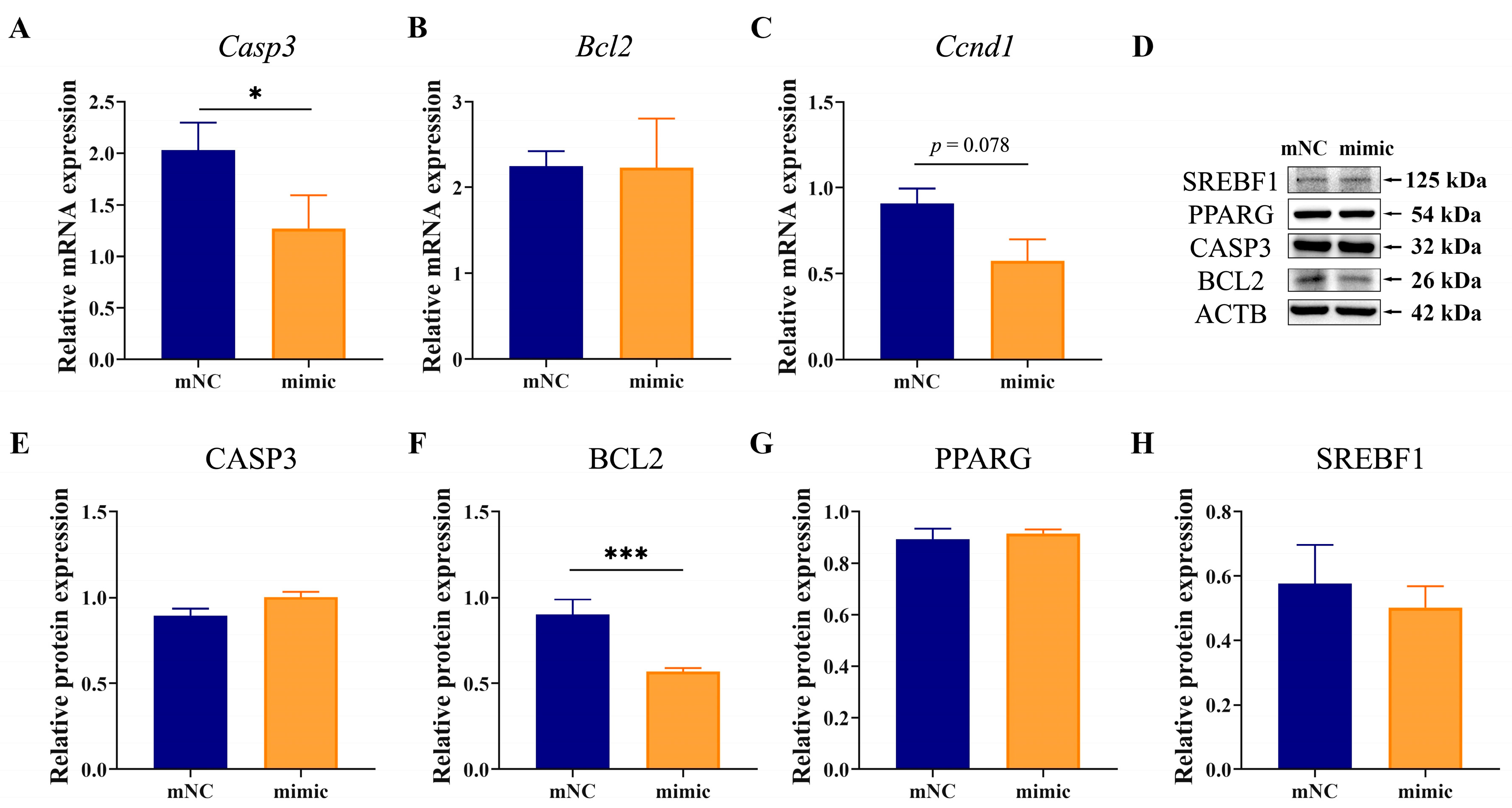
2.5. Overexpression of MiR-130b Promoted Apoptosis by Inhibiting BCL2 Expression in Rat IMAs
2.6. MiR-130b Inhibition Repressed Apoptosis by Increasing BCL2 Expression in Rat IMAs
2.7. Overexpression of MiR-130b Promoted Apoptosis by Inhibiting BCL2 and Increasing CASP3 Expression in Rat IMAs
3. Discussion
4. Materials and Methods
4.1. Animals and Ethics Statements
4.2. Cell Isolation, Culture, and Differentiation
4.3. Cell Transfection
4.4. Oil Red O Staining Extraction Assay and TG Content Determination
4.5. RNA Extraction and Quantitative Real-Time PCR
4.6. Western Blot Assay
4.7. EdU Staining
4.8. Cell Cycle Analysis
4.9. Apoptosis Detection by Annexin V-FITC Method
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BCL2 | B-cell lymphoma 2 |
| CASP3 | Caspase 3 |
| CCND1 | Cyclin D1 |
| CEBP | CCAAT enhancer binding protein |
| CRC | Colorectal cancer |
| FAP | Fibro/adipogenic progenitor |
| GC | Glucocorticoid |
| HFD | High-fat diet |
| HSD11B1 | Hydroxysteroid 11-beta dehydrogenase 1 |
| IMA | Intramuscular preadipocyte |
| IMF | Intramuscular fat |
| LPL | Lipoprotein lipase |
| MiRNAs | MicroRNAs |
| NR3C1 | Nuclear receptor subfamily 3 group C member 1 |
| PPARG | Peroxisome proliferator-activated receptor gamma |
| RT-qPCR | Quantitative real-time PCR |
| SCF | Subcutaneous fat |
| SREBF1 | Sterol regulatory element binding transcription factor 1 |
| TG | Triglyceride |
| WAT | White adipose tissue |
References
- Xu, K.; Ji, M.; Huang, X.; Peng, Y.; Wu, W.; Zhang, J. Differential Regulatory Roles of MicroRNAs in Porcine Intramuscular and Subcutaneous Adipocytes. J. Agric. Food Chem. 2020, 68, 3954–3962. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Yao, Y.; Ye, N.; Zhao, Y.; Ye, Z.; Wei, W.; Zhang, L.; Chen, J. The myokine CCL5 recruits subcutaneous preadipocytes and promotes intramuscular fat deposition in obese mice. Am. J. Physiol. Cell Physiol. 2024, 326, C1320–C1333. [Google Scholar] [CrossRef]
- Sasaki, Y.; Nagai, K.; Nagata, Y.; Doronbekov, K.; Nishimura, S.; Yoshioka, S.; Fujita, T.; Shiga, K.; Miyake, T.; Taniguchi, Y.; et al. Exploration of genes showing intramuscular fat deposition-associated expression changes in musculus longissimus muscle. Anim. Genet. 2006, 37, 40–46. [Google Scholar] [CrossRef]
- Cristancho, A.G.; Lazar, M.A. Forming functional fat: A growing understanding of adipocyte differentiation. Nat. Rev. Mol. Cell Biol. 2011, 12, 722–734. [Google Scholar] [CrossRef] [PubMed]
- Ambele, M.A.; Dhanraj, P.; Giles, R.; Pepper, M.S. Adipogenesis: A Complex Interplay of Multiple Molecular Determinants and Pathways. Int. J. Mol. Sci. 2020, 21, 4283. [Google Scholar] [CrossRef]
- Ma, F.; Zhan, Y.; Bartolomé-Cabrero, R.; Ying, W.; Asano, M.; Huang, Z.; Xiao, C.; González-Martín, A. Analysis of a miR-148a Targetome in B Cell Central Tolerance. Front. Immunol. 2022, 13, 861655. [Google Scholar] [CrossRef] [PubMed]
- Karbiener, M.; Scheideler, M. MicroRNA Functions in Brite/Brown Fat—Novel Perspectives towards Anti-Obesity Strategies. Comput. Struct. Biotechnol. J. 2014, 11, 101–105. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.C.; Wang, J.; Kong, J.; Qi, Y.; Quigg, R.J.; Li, X. miR-17-92 cluster accelerates adipocyte differentiation by negatively regulating tumor-suppressor Rb2/p130. Proc. Natl. Acad. Sci. USA 2008, 105, 2889–2894. [Google Scholar] [CrossRef]
- Wosczyna, M.N.; Carbajal, E.E.P.; Wagner, M.W.; Paredes, S.; Konishi, C.T.; Liu, L.; Wang, T.T.; Walsh, R.A.; Gan, Q.; Morrissey, C.S.; et al. Targeting microRNA-mediated gene repression limits adipogenic conversion of skeletal muscle mesenchymal stromal cells. Cell Stem Cell 2021, 28, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Dong, P.; Mai, Y.; Zhang, Z.; Mi, L.; Wu, G.; Chu, G.; Yang, G.; Sun, S. MiR-15a/b promote adipogenesis in porcine pre-adipocyte via repressing FoxO1. Acta Biochim. Biophys. Sin. 2014, 46, 565–571. [Google Scholar] [CrossRef]
- Shen, L.; Zhang, Y.; Du, J.; Chen, L.; Luo, J.; Li, X.; Li, M.; Tang, G.; Zhang, S.; Zhu, L. MicroRNA-23a regulates 3T3-L1 adipocyte differentiation. Gene 2016, 575, 761–764. [Google Scholar] [CrossRef]
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef]
- Pan, S.; Yang, X.; Jia, Y.; Li, R.; Zhao, R. Microvesicle-shuttled miR-130b reduces fat deposition in recipient primary cultured porcine adipocytes by inhibiting PPAR-γ expression. J. Cell. Physiol. 2014, 229, 631–639. [Google Scholar] [CrossRef]
- Pan, S.; Zheng, Y.; Zhao, R.; Yang, X. MicroRNA-130b and microRNA-374b mediate the effect of maternal dietary protein on offspring lipid metabolism in Meishan pigs. Br. J. Nutr. 2013, 109, 1731–1738. [Google Scholar] [CrossRef]
- Pan, S.; Yang, X.; Jia, Y.; Li, Y.; Chen, R.; Wang, M.; Cai, D.; Zhao, R. Intravenous injection of microvesicle-delivery miR-130b alleviates high-fat diet-induced obesity in C57BL/6 mice through translational repression of PPAR-γ. J. Biomed. Sci. 2015, 22, 86. [Google Scholar] [CrossRef]
- Luo, W.; Kim, Y.; Jensen, M.E.; Herlea-Pana, O.; Wang, W.; Rudolph, M.C.; Friedman, J.E.; Chernausek, S.D.; Jiang, S. miR-130b/301b Is a Negative Regulator of Beige Adipogenesis and Energy Metabolism In Vitro and In Vivo. Diabetes 2022, 71, 2360–2371. [Google Scholar] [CrossRef]
- Lai, K.W.; Koh, K.X.; Loh, M.; Tada, K.; Subramaniam, M.M.; Lim, X.Y.; Vaithilingam, A.; Salto-Tellez, M.; Iacopetta, B.; Ito, Y.; et al. MicroRNA-130b regulates the tumour suppressor RUNX3 in gastric cancer. Eur. J. Cancer 2010, 46, 1456–1463. [Google Scholar] [CrossRef]
- Song, D.; Zhang, Q.; Zhang, H.; Zhan, L.; Sun, X. MiR-130b-3p promotes colorectal cancer progression by targeting CHD9. Cell Cycle 2022, 21, 585–601. [Google Scholar] [CrossRef] [PubMed]
- Abebe, B.K.; Wang, J.; Guo, J.; Wang, H.; Li, A.; Zan, L. A review of the role of epigenetic studies for intramuscular fat deposition in beef cattle. Gene 2024, 908, 148295. [Google Scholar] [CrossRef]
- Engin, A.B. MicroRNA and Adipogenesis. In Obesity and Lipotoxicity; Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2017; Volume 960, pp. 489–509. [Google Scholar] [CrossRef]
- Engin, A.B.; Engin, A. MicroRNAs as Epigenetic Regulators of Obesity. In Obesity and Lipotoxicity; Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2024; Volume 1460, pp. 595–627. [Google Scholar] [CrossRef]
- Song, X.; Song, Y.; Zhang, J.; Hu, Y.; Zhang, L.; Huang, Z.; Raza, S.H.A.; Jiang, C.; Ma, Y.; Ma, Y.; et al. Regulatory role of exosome-derived miRNAs and other contents in adipogenesis. Exp. Cell Res. 2024, 441, 114168. [Google Scholar] [CrossRef]
- Li, H.; Liu, P.; Li, D.; Wang, Z.; Ding, Z.; Zhou, M.; Chen, X.; Miao, M.; Ding, J.; Lin, W.; et al. STAT3/miR-130b-3p/MBNL1 feedback loop regulated by mTORC1 signaling promotes angiogenesis and tumor growth. J. Exp. Clin. Cancer Res. 2022, 41, 297. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Tu, W.Z.; Guo, J.B.; Song, G.; Zhang, J.; Chen, C.C.; Chen, P.J. A Bioinformatic Analysis of MicroRNAs’ Role in Human Intervertebral Disc Degeneration. Pain Med. 2019, 20, 2459–2471. [Google Scholar] [CrossRef]
- Wu, X.; Yao, C.; Kong, J.; Tian, Y.; Fan, Y.; Zhang, Z.; Han, J.; Wu, S. Molecular mechanism underlying miR-130b-Sp1 transcriptional regulation in LPS-induced upregulation of MUC5AC in the bile duct epithelium. Mol. Med. Rep. 2021, 23, 106. [Google Scholar] [CrossRef]
- Guo, Q.; Zhu, X.; Wei, R.; Zhao, L.; Zhang, Z.; Yin, X.; Zhang, Y.; Chu, C.; Wang, B.; Li, X. miR-130b-3p regulates M1 macrophage polarization via targeting IRF1. J. Cell. Physiol. 2021, 236, 2008–2022. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wei, D.; Cheng, G.; Li, S.; Wang, L.; Wang, Y.; Wang, X.; Zhang, S.; Wang, H.; Zan, L. Bta-miR-130a/b regulates preadipocyte differentiation by targeting PPARG and CYP2U1 in beef cattle. Mol. Cell. Probes 2018, 42, 10–17. [Google Scholar] [CrossRef]
- Wang, Y.C.; Li, Y.; Wang, X.Y.; Zhang, D.; Zhang, H.; Wu, Q.; He, Y.Q.; Wang, J.Y.; Zhang, L.; Xia, H.; et al. Circulating miR-130b mediates metabolic crosstalk between fat and muscle in overweight/obesity. Diabetologia 2013, 56, 2275–2285. [Google Scholar] [CrossRef]
- Pan, S.; Cui, Y.; Dong, X.; Zhang, T.; Xing, H. MicroRNA-130b attenuates dexamethasone-induced increase of lipid accumulation in porcine preadipocytes by suppressing PPAR-γ expression. Oncotarget 2017, 8, 87928–87943. [Google Scholar] [CrossRef]
- Ahmad, I.; Mui, E.; Galbraith, L.; Patel, R.; Tan, E.H.; Salji, M.; Rust, A.G.; Repiscak, P.; Hedley, A.; Markert, E.; et al. Sleeping Beauty screen reveals Pparg activation in metastatic prostate cancer. Proc. Natl. Acad. Sci. USA 2016, 113, 8290–8295. [Google Scholar] [CrossRef]
- Galbraith, L.; Leung, H.Y.; Ahmad, I. Lipid pathway deregulation in advanced prostate cancer. Pharmacol. Res. 2018, 131, 177–184. [Google Scholar] [CrossRef]
- Kersten, S. Physiological regulation of lipoprotein lipase. Biochim. Biophys. Acta 2014, 1841, 919–933. [Google Scholar] [CrossRef]
- Anagnostis, P.; Athyros, V.G.; Tziomalos, K.; Karagiannis, A.; Mikhailidis, D.P. Clinical review: The pathogenetic role of cortisol in the metabolic syndrome: A hypothesis. J. Clin. Endocrinol. Metab. 2009, 94, 2692–2701. [Google Scholar] [CrossRef]
- Lundgren, M.; Burén, J.; Ruge, T.; Myrnäs, T.; Eriksson, J.W. Glucocorticoids down-regulate glucose uptake capacity and insulin-signaling proteins in omental but not subcutaneous human adipocytes. J. Clin. Endocrinol. Metab. 2004, 89, 2989–2997. [Google Scholar] [CrossRef]
- Sun, X.J.; Li, Z.H.; Zhang, Y.; Zhou, G.; Zhang, J.Q.; Deng, J.M.; Bai, J.; Liu, G.N.; Li, M.H.; MacNee, W.; et al. Combination of erythromycin and dexamethasone improves corticosteroid sensitivity induced by CSE through inhibiting PI3K-δ/Akt pathway and increasing GR expression. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2015, 309, L139–L146. [Google Scholar] [CrossRef]
- Chu, W.; Wei, W.; Han, H.; Gao, Y.; Liu, K.; Tian, Y.; Jiang, Z.; Zhang, L.; Chen, J. Muscle-specific downregulation of GR levels inhibits adipogenesis in porcine intramuscular adipocyte tissue. Sci. Rep. 2017, 7, 510. [Google Scholar] [CrossRef]
- Kupczyk, D.; Bilski, R.; Kozakiewicz, M.; Studzińska, R.; Kędziora-Kornatowska, K.; Kosmalski, T.; Pedrycz-Wieczorska, A.; Głowacka, M. 11β-HSD as a New Target in Pharmacotherapy of Metabolic Diseases. Int. J. Mol. Sci. 2022, 23, 8984. [Google Scholar] [CrossRef]
- Bao, D.; Li, M.; Zhou, D.; Zhuang, C.; Ge, Z.; Wei, Q.; Zhang, L. miR-130b-3p is high-expressed in polycystic ovarian syndrome and promotes granulosa cell proliferation by targeting SMAD4. J. Steroid Biochem. Mol. Biol. 2021, 209, 105844. [Google Scholar] [CrossRef]
- Kuszczak, B.; Wróbel, T.; Wicherska-Pawłowska, K.; Rybka, J. The Role of BCL-2 and PD-1/PD-L1 Pathway in Pathogenesis of Myelodysplastic Syndromes. Int. J. Mol. Sci. 2023, 24, 4708. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, X.; Zhou, K.; Ling, X.; Zhang, J.; Wu, P.; Zhang, T.; Xie, K.; Dai, G. miRNA-10a-5p Targeting the BCL6 Gene Regulates Proliferation, Differentiation and Apoptosis of Chicken Myoblasts. Int. J. Mol. Sci. 2022, 23, 9545. [Google Scholar] [CrossRef]
- Han, F.; Zhou, L.; Zhao, L.; Wang, L.; Liu, L.; Li, H.; Qiu, J.; He, J.; Liu, N. Identification of miRNA in Sheep Intramuscular Fat and the Role of miR-193a-5p in Proliferation and Differentiation of 3T3-L1. Front. Genet. 2021, 12, 633295. [Google Scholar] [CrossRef]
- Mens, M.M.J.; Ghanbari, M. Cell Cycle Regulation of Stem Cells by MicroRNAs. Stem Cell Rev. Rep. 2018, 14, 309–322. [Google Scholar] [CrossRef]
- Tong, H.; Wang, L.; Shi, J.; Jin, H.; Zhang, K.; Bao, Y.; Wu, Y.; Cheng, Y.; Liu, P.; Wang, C. Upregulated miR-322-5p regulates cell cycle and promotes cell proliferation and apoptosis by directly targeting Wee1 in mice liver injury. Cell Cycle 2022, 21, 2635–2650. [Google Scholar] [CrossRef] [PubMed]
- Sinha, P.B.; Tesfaye, D.; Rings, F.; Hossien, M.; Hoelker, M.; Held, E.; Neuhoff, C.; Tholen, E.; Schellander, K.; Salilew-Wondim, D. MicroRNA-130b is involved in bovine granulosa and cumulus cells function, oocyte maturation and blastocyst formation. J. Ovarian Res. 2017, 10, 37. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Shiina, M.; Dasgupta, P.; Kulkarni, P.; Kato, T.; Wong, R.K.; Tanaka, Y.; Shahryari, V.; Maekawa, S.; Yamamura, S.; et al. Upregulation of miR-130b Contributes to Risk of Poor Prognosis and Racial Disparity in African-American Prostate Cancer. Cancer Prev. Res. 2019, 12, 585–598. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, L.; Fan, C.; Mu, H.; Han, M.; Liu, T.; Xie, L.; Gao, Q. Promotion of Cervical Cancer Cell Proliferation by miR-130b Expression Level Changes and Inhibition of its Apoptosis by Targeting CDKN1A Gene. Curr. Cancer Drug Targets 2022, 22, 153–168. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) |
| miR-130b mimic | CAGTGCAATGATGAAAGGGCAT |
| GCCCTTTCATCATTGCACTGTT | |
| miR-130b mimic negative control | TTCTCCGAACGTGTCACGTTT |
| ACGTGACACGTTCGGAGAATT | |
| miR-130b inhibitor | ATGCCCTTTCATCATTGCACTG |
| miR-130b inhibitor negative control | CAGTACTTTTGTGTAGTACAA |
| Universal primer | TAGAGTGAGTGTAGCGAGCA |
| Poly (T) adapter | TAGAGTGAGTGTAGCGAGCACAGAATTAATACGACTCACTATAGGTTTTTTTTTTTTTTTTVN |
| Name | Primer Sequences |
|---|---|
| Pparg (NM_013124.3) | F: TTGATTTCTCCAGCATTTC |
| R: TGATCGCACTTTGGTATT | |
| Lpl (NM_012598.2) | F: TTGTCCCACTCCGTATCTG R: TATGGTTATCAAGCTCCC |
| Nr3c1 (NM_012576.2) | F: AACGTCTGCAACTGGGTC |
| R: TGCTTTGGTCTGTGGGATA | |
| Hsd11b1 (NM_017080.2) | F: GGTGTCTCGCTGCCTTGA |
| R: TTCTTCGCACAGAGTGGATA | |
| Bcl2 (NM_016993.2) | F: CGGGAGAACAGGGTATGA |
| R: CTTCATCTCCAGTATCCCAC | |
| Casp3 (NM_012922.2) | F: CTGGACTGCGGTATTGAG |
| R: GGGTGCGCTAGAGTAAGC | |
| Srebf1 (NM_001276707.1) | F: CACTTACGGTCAGCACTT |
| R: CACAACTCACTGGACTTAGA | |
| Ccnd1 (NM_171992.5) | F: GCGAGGAGCAGAAGTGCGAAGA |
| R: GGCGGATAGAGTTGTCAGTGTAGATG | |
| Gapdh (NM_017008.4) | F: CCTGGAGAAACCTGCCAAG |
| R: CACAGGAGACAACCTGGTCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Chen, Y.; Wang, L.; Cheng, J.; Du, M.; Pan, S. Rno-miR-130b Attenuates Lipid Accumulation Through Promoting Apoptosis and Inhibiting Differentiation in Rat Intramuscular Adipocytes. Int. J. Mol. Sci. 2025, 26, 1399. https://doi.org/10.3390/ijms26041399
Yu Y, Chen Y, Wang L, Cheng J, Du M, Pan S. Rno-miR-130b Attenuates Lipid Accumulation Through Promoting Apoptosis and Inhibiting Differentiation in Rat Intramuscular Adipocytes. International Journal of Molecular Sciences. 2025; 26(4):1399. https://doi.org/10.3390/ijms26041399
Chicago/Turabian StyleYu, Yichen, Yongfang Chen, Lijun Wang, Ji Cheng, Min Du, and Shifeng Pan. 2025. "Rno-miR-130b Attenuates Lipid Accumulation Through Promoting Apoptosis and Inhibiting Differentiation in Rat Intramuscular Adipocytes" International Journal of Molecular Sciences 26, no. 4: 1399. https://doi.org/10.3390/ijms26041399
APA StyleYu, Y., Chen, Y., Wang, L., Cheng, J., Du, M., & Pan, S. (2025). Rno-miR-130b Attenuates Lipid Accumulation Through Promoting Apoptosis and Inhibiting Differentiation in Rat Intramuscular Adipocytes. International Journal of Molecular Sciences, 26(4), 1399. https://doi.org/10.3390/ijms26041399