
Proteomic Dynamics in the Interaction of Susceptible and Resistant Tomato Cultivars and Potato Cyst Nematodes
 , ,
, ,
Abstract
1. Introduction
2. Results
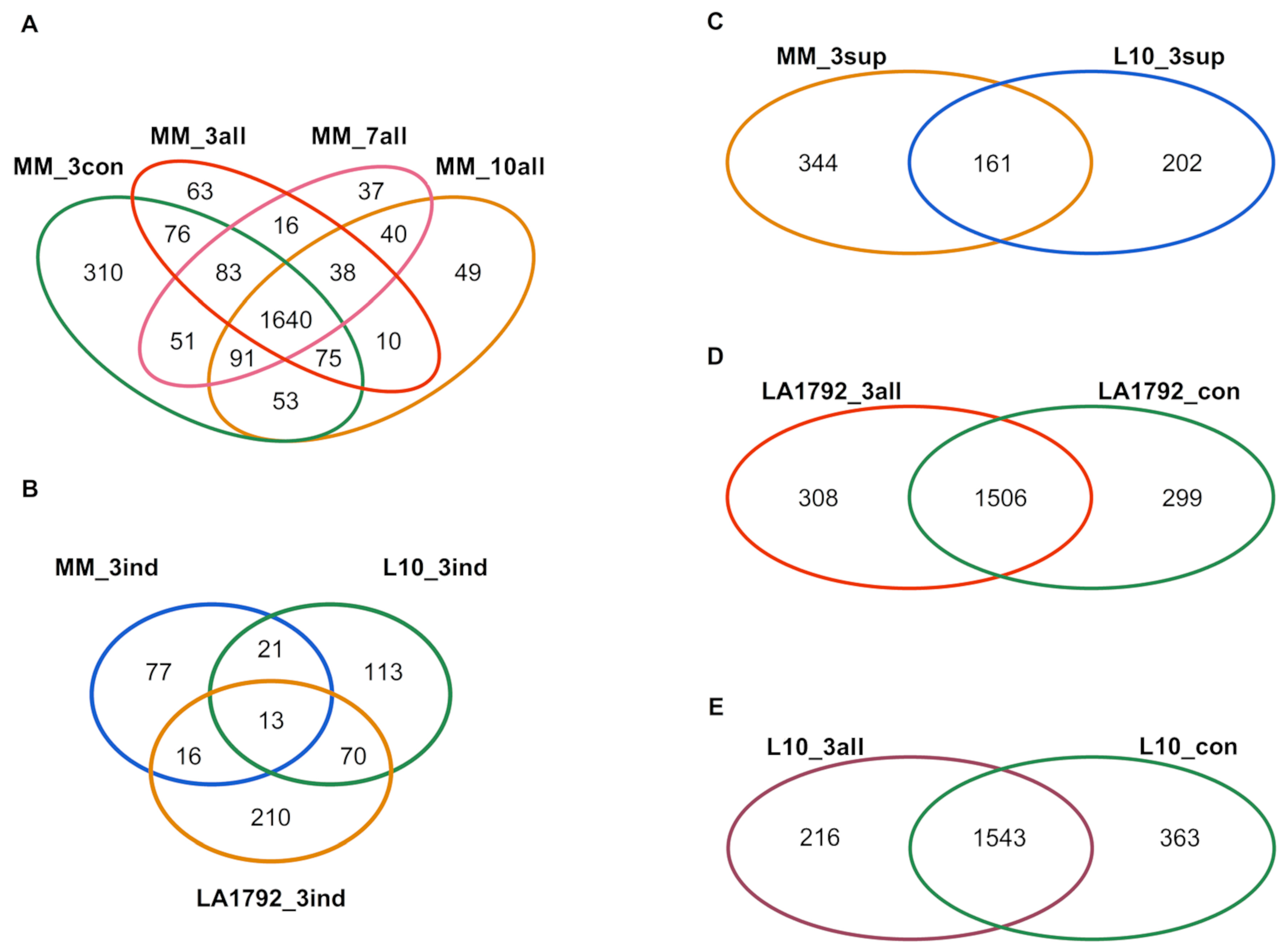
2.1. Protein Identification by Mass Spectrometry
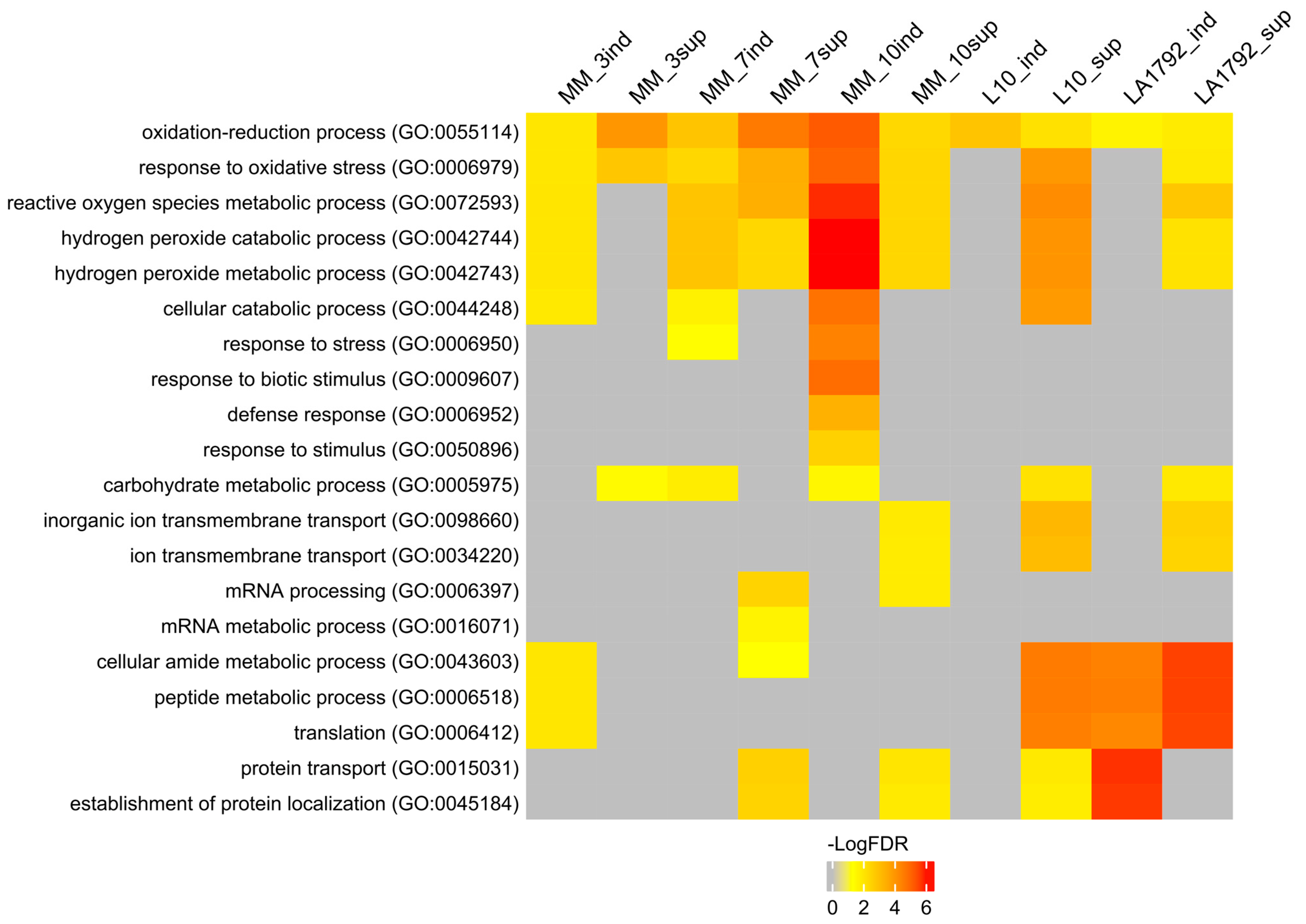
2.2. Study of the Tomato Orthologs in Arabidopsis
3. Discussion
3.1. Tomato Proteome Analysis
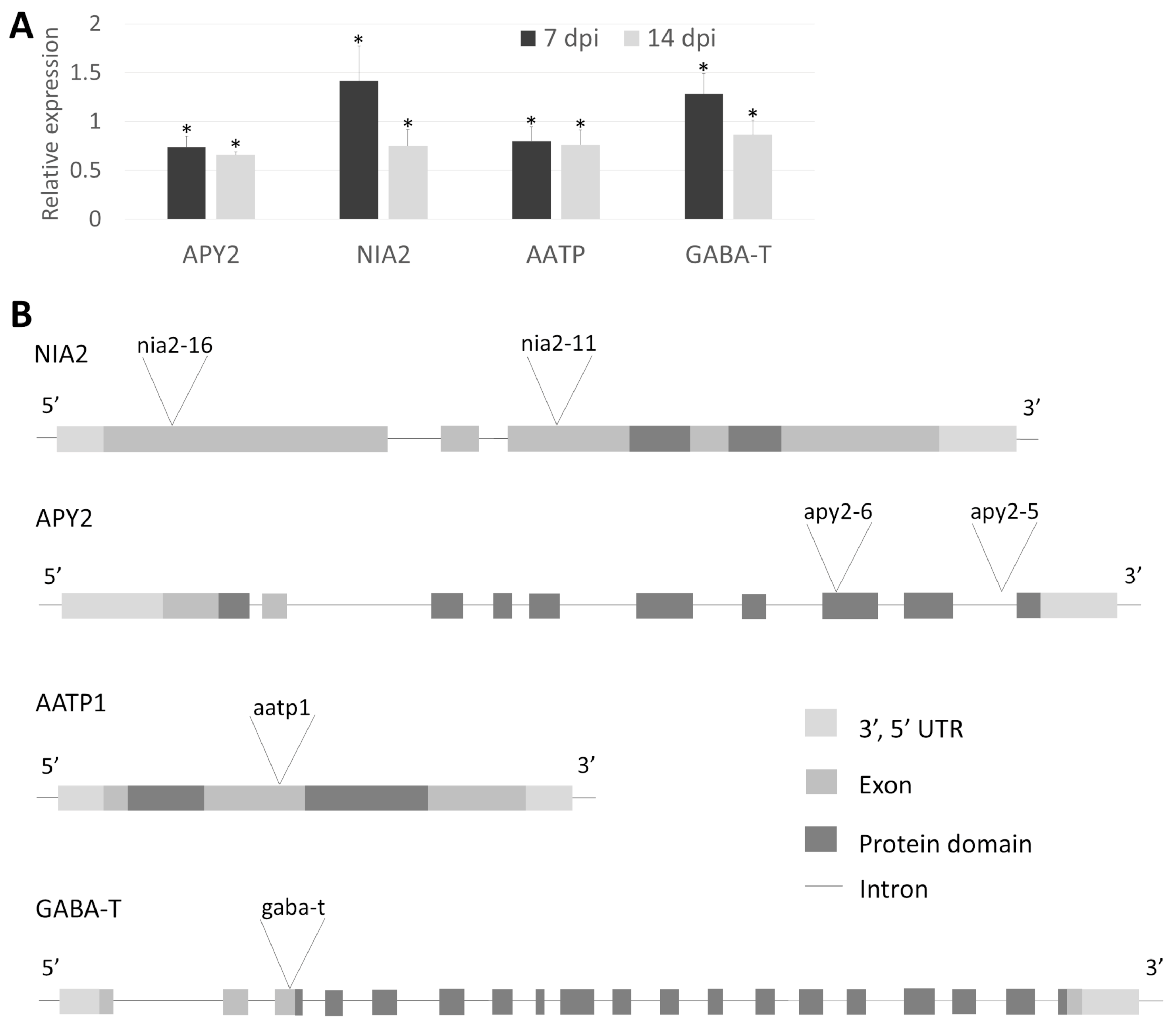
3.2. Selected Mutant Genes Analysis
4. Materials and Methods
4.1. Plant Material
4.2. Tomato Roots Infestation with G. rostochiensis Larvae
4.3. Protein Isolation and Mass Spectrometry
4.4. Protein Identification and Quantitation
4.5. RNA Isolation and RT-qPCR
4.6. Tomato Orthologues Analysis in Arabidopsis
4.7. Functional Protein Analysis
4.8. Nematode Infection Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McCarter, J.P. Molecular Approaches Toward Resistance to Plant-Parasitic Nematodes. In Cell Biology of Plant Nematode Parasitism; Berg, R.H., Taylor, C.G., Eds.; Springer: Berlin, Heidelberg, 2009; pp. 239–267. ISBN 978-3-540-85215-5. [Google Scholar]
- Sasser, J.N.; Freckman, D.W. A World Perspective on Nematology: The Role of the Society; Society of nematologists: Hyattsville, MD, USA, 1987. [Google Scholar]
- Abd-Elgawad, M.M.M.; Askary, T.H. Impact of Phytonematodes on Agriculture Economy. In Biocontrol Agents of Phytonematodes; CABI: Wallingford, UK, 2015; pp. 3–49. [Google Scholar] [CrossRef]
- Sobczak, M.; Golinowski, W.; Grundler, F.M.W. Changes in the Structure of Arabidopsis Thaliana Roots Induced during Development of Males of the Plant Parasitic Nematode Heterodera schachtii. Eur. J. Plant Pathol. 1997, 103, 113–124. [Google Scholar]
- Mitchum, M.G.; Hussey, R.S.; Baum, T.J.; Wang, X.; Elling, A.A.; Wubben, M.; Davis, E.L. Nematode Effector Proteins: An Emerging Paradigm of Parasitism. New Phytol. 2013, 199, 879–894. [Google Scholar] [PubMed]
- Grundler, F.M.W.; Sobczak, M.; Golinowski, W. Formation of Wall Openings in Root Cells of Arabidopsis thaliana Following Infection by the Plant-Parasitic Nematode Heterodera schachtii. Eur. J. Plant Pathol. 1998, 104, 545–551. [Google Scholar]
- Ohtsu, M.; Sato, Y.; Kurihara, D.; Suzaki, T.; Kawaguchi, M.; Maruyama, D.; Higashiyama, T. Spatiotemporal Deep Imaging of Syncytium Induced by the Soybean Cyst Nematode Heterodera Glycines. Protoplasma 2017, 254, 2107–2115. [Google Scholar]
- Böckenhoff, A.; Grundler, F.M.W. Studies on the Nutrient Uptake by the Beet Cyst Nematode Heterodera schachtii by in Situ Microinjection of Fluorescent Probes into the Feeding Structures in Arabidopsis thaliana. Parasitology 1994, 109, 249–254. [Google Scholar]
- Todd, J.N.A.; Carreón-Anguiano, K.G.; Islas-Flores, I.; Canto-Canché, B. Microbial Effectors: Key Determinants in Plant Health and Disease. Microorganisms 2022, 10, 1980. [Google Scholar] [CrossRef]
- Lee, C.; Chronis, D.; Kenning, C.; Peret, B.; Hewezi, T.; Davis, E.L.; Baum, T.J.; Hussey, R.; Bennett, M.; Mitchum, M.G. The Novel Cyst Nematode Effector Protein 19C07 Interacts with the Arabidopsis Auxin Influx Transporter LAX3 to Control Feeding Site Development. Plant Physiol. 2011, 155, 866–880. [Google Scholar]
- Wang, J.; Replogle, A.; Hussey, R.; Baum, T.; Wang, X.; Davis, E.L.; Mitchum, M.G. Identification of Potential Host Plant Mimics of CLAVATA3/ESR (CLE)-like Peptides from the Plant-Parasitic Nematode Heterodera schachtii. Mol. Plant Pathol. 2011, 12, 177–186. [Google Scholar]
- Tohge, T.; Watanabe, M.; Hoefgen, R.; Fernie, A.R. Shikimate and Phenylalanine Biosynthesis in the Green Lineage. Front. Plant Sci. 2013, 4, 62. [Google Scholar]
- Verma, A.; Lee, C.; Morriss, S.; Odu, F.; Kenning, C.; Rizzo, N.; Spollen, W.G.; Lin, M.; Mcrae, A.G.; Givan, S.A.; et al. The Novel Cyst Nematode Effector Protein 30D08 Targets Host Nuclear Functions to Alter Gene Expression in Feeding Sites. New Phytol. 2018, 219, 697–713. [Google Scholar]
- Rehman, S.; Postma, W.; Tytgat, T.; Prins, P.; Qin, L.; Overmars, H.; Vossen, J.; Spiridon, L.N.; Petrescu, A.J.; Goverse, A.; et al. A Secreted SPRY Domain-Containing Protein (SPRYSEC) from the Plant-Parasitic Nematode Globodera Rostochiensis Interacts with a CC-NB-LRR Protein from a Susceptible Tomato. Mol. Plant Microbe Interact. 2009, 22, 330–340. [Google Scholar] [PubMed]
- Lozano-Torres, J.L.; Wilbers, R.H.P.; Gawronski, P.; Boshoven, J.C.; Finkers-Tomczak, A.; Cordewener, J.H.G.; America, A.H.P.; Overmars, H.A.; Van ‘t Klooster, J.W.; Baranowski, L.; et al. Dual Disease Resistance Mediated by the Immune Receptor Cf-2 in Tomato Requires a Common Virulence Target of a Fungus and a Nematode. Proc. Natl. Acad. Sci. USA 2012, 109, 10119–10124. [Google Scholar]
- van Steenbrugge, J.J.M.; van den Elsen, S.; Holterman, M.; Lozano-Torres, J.L.; Putker, V.; Thorpe, P.; Goverse, A.; Sterken, M.G.; Smant, G.; Helder, J. Comparative Genomics among Cyst Nematodes Reveals Distinct Evolutionary Histories among Effector Families and an Irregular Distribution of Effector-Associated Promoter Motifs. Mol. Ecol. 2023, 32, 1515–1529. [Google Scholar] [PubMed]
- Swiecicka, M.; Filipecki, M.; Lont, D.; Van Vliet, J.; Qin, L.; Goverse, A.; Bakker, J.; Helder, J. Dynamics in the Tomato Root Transcriptome on Infection with the Potato Cyst Nematode Globodera rostochiensis. Mol. Plant Pathol. 2009, 10, 487–500. [Google Scholar]
- Szakasits, D.; Heinen, P.; Wieczorek, K.; Hofmann, J.; Wagner, F.; Kreil, D.P.; Sykacek, P.; Grundler, F.M.; Bohlmann, H. The Transcriptome of Syncytia Induced by the Cyst Nematode Heterodera schachtii in Arabidopsis Roots. Plant J. 2009, 57, 771–784. [Google Scholar]
- Matuszkiewicz, M.; Sobczak, M.; Cabrera, J.; Escobar, C.; Karpiński, S.; Filipecki, M. The Role of Programmed Cell Death Regulator. Front. Plant Sci. 2018, 9, 314. [Google Scholar]
- Jaubert-Possamai, S.; Noureddine, Y.; Favery, B. MicroRNAs, New Players in the Plant-Nematode Interaction. Front. Plant Sci. 2019, 10, 1180. [Google Scholar] [CrossRef]
- Koter, M.D.; Święcicka, M.; Matuszkiewicz, M.; Pacak, A.; Derebecka, N.; Filipecki, M. The miRNAome Dynamics during Developmental and Metabolic Reprogramming of Tomato Root Infected with Potato Cyst Nematode. Plant Sci. 2018, 268, 18–29. [Google Scholar] [CrossRef]
- Labudda, M.; Różańska, E.; Szewińska, J.; Sobczak, M.; Dzik, J.M. Protease Activity and Phytocystatin Expression in Arabidopsis thaliana upon Heterodera schachtii Infection. Plant Physiol. Biochem. 2016, 109, 416–429. [Google Scholar]
- Filipecki, M.; Żurczak, M.; Matuszkiewicz, M.; Święcicka, M.; Kurek, W.; Olszewski, J.; Koter, M.D.; Lamont, D.; Sobczak, M. Profiling the Proteome of Cyst Nematode-Induced Syncytia on Tomato Roots. Int. J. Mol. Sci. 2021, 22, 12147. [Google Scholar] [CrossRef]
- Sobczak, M.; Avrova, A.; Jupowicz, J.; Phillips, M.S.; Ernst, K.; Kumar, A. Characterization of Susceptibility and Resistance Responses to Potato Cyst Nematode (Globodera Spp.) Infection of Tomato Lines in the Absence and Presence of the Broad-Spectrum Nematode Resistance Hero Gene. Mol. Plant Microbe Interact. 2005, 18, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska-Bronk, J.; Czarny, M.; Wiśniewska, A.; Fudali, S.; Baranowski, Ł.; Sobczak, M.; Święcicka, M.; Matuszkiewicz, M.; Brzyżek, G.; Wroblewski, T.; et al. Suppression of NGB and NAB/ERabp1 in Tomato Modifies Root Responses to Potato Cyst Nematode Infestation. Mol. Plant Pathol. 2015, 16, 334–348. [Google Scholar] [CrossRef] [PubMed]
- Święcicka, M.; Skowron, W.; Cieszyński, P.; Dąbrowska-Bronk, J.; Matuszkiewicz, M.; Filipecki, M.; Koter, M.D. The Suppression of Tomato Defence Response Genes upon Potato Cyst Nematode Infection Indicates a Key Regulatory Role of miRNAs. Plant Physiol. Biochem. 2017, 113, 51–55. [Google Scholar] [CrossRef]
- Du, Z.; Zhou, X.; Ling, Y.; Zhang, Z.; Su, Z. agriGO: A GO Analysis Toolkit for the Agricultural Community. Nucleic Acids Res. 2010, 38, W64–W70. [Google Scholar] [CrossRef]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO Analysis Toolkit for the Agricultural Community, 2017 Update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, H.; Lin, H.; Roy, S.; Shaler, T.A.; Hill, L.R.; Norton, S.; Kumar, P.; Anderle, M.; Becker, C.H. Quantification of Proteins and Metabolites by Mass Spectrometry without Isotopic Labeling or Spiked Standards. Anal. Chem. 2003, 75, 4818–4826. [Google Scholar] [CrossRef]
- Clark, G.; Brown, K.A.; Tripathy, M.K.; Roux, S.J. Recent Advances Clarifying the Structure and Function of Plant Apyrases (Nucleoside Triphosphate Diphosphohydrolases). Int. J. Mol. Sci. 2021, 22, 3283. [Google Scholar] [CrossRef]
- Machchhu, F.; Wany, A. Protein S-Nitrosylation in Plants under Biotic Stress. Theor. Exp. Plant Physiol. 2023, 35, 331–339. [Google Scholar] [CrossRef]
- Li, L.; Dou, N.; Zhang, H.; Wu, C. The Versatile GABA in Plants. Plant Signal. Behav. 2021, 16, 1862565. [Google Scholar] [CrossRef]
- Liu, X.; Inoue, H.; Tang, X.; Tan, Y.; Xu, X.; Wang, C.; Jiang, C.J. Rice OsAAA-ATPase1 is Induced during Blast Infection in a Salicylic Acid-Dependent Manner, and Promotes Blast Fungus Resistance. Int. J. Mol. Sci. 2020, 21, 1443. [Google Scholar] [CrossRef]
- Foss, E.J.; Radulovic, D.; Shaffer, S.A.; Ruderfer, D.M.; Bedalov, A.; Goodlett, D.R.; Kruglyak, L. Genetic Basis of Proteome Variation in Yeast. Nat. Genet. 2007, 39, 1369–1375. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Keurentjes, J.J.; Bouwmeester, H.; America, T.; Verstappen, F.W.; Ward, J.L.; Beale, M.H.; de Vos, R.C.; Dijkstra, M.; Scheltema, R.A.; et al. System-Wide Molecular Evidence for Phenotypic Buffering in Arabidopsis. Nat. Genet. 2009, 41, 166–167. [Google Scholar] [CrossRef] [PubMed]
- Stare, T.; Stare, K.; Weckwerth, W.; Wienkoop, S.; Gruden, K. Comparison between Proteome and Transcriptome Response in Potato (Solanum tuberosum L.) Leaves Following Potato Virus Y (PVY) Infection. Proteomes 2017, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Ernst, K.; Kumar, A.; Kriseleit, D.; Kloos, D.U.; Phillips, M.S.; Ganal, M.W. The Broad-Spectrum Potato Cyst Nematode Resistance Gene (Hero) from Tomato Is the Only Member of a Large Gene Family of NBS-LRR Genes with an Unusual Amino Acid Repeat in the LRR Region. Plant J. 2002, 31, 127–136. [Google Scholar] [CrossRef]
- Attieh, J.; Sparace, S.A.; Saini, H.S. Purification and Properties of Multiple Isoforms of a Novel Thiol Methyltransferase Involved in the Production of Volatile Sulfur Compounds from Brassica Oleracea. Arch. Biochem. Biophys. 2000, 380, 257–266. [Google Scholar] [CrossRef]
- Hilleary, R. Battle of the Bulge: The ARP2/3 Complex Form(in)s an Actin Phalanx to Thwart Fungal Infection. Plant Cell 2021, 33, 2910–2911. [Google Scholar] [CrossRef]
- Labudda, M.; Tokarz, K.; Tokarz, B.; Muszyńska, E.; Gietler, M.; Górecka, M.; Różańska, E.; Rybarczyk-Płońska, A.; Fidler, J.; Prabucka, B.; et al. Reactive Oxygen Species Metabolism and Photosynthetic Performance in Leaves of Hordeum vulgare Plants Co-Infested with Heterodera filipjevi and Aceria tosichella. Plant Cell Rep. 2020, 39, 1719–1741. [Google Scholar] [CrossRef]
- Labudda, M.; Różańska, E.; Gietler, M.; Fidler, J.; Muszyńska, E.; Prabucka, B.; Morkunas, I. Cyst Nematode Infection Elicits Alteration in the Level of Reactive Nitrogen Species, Protein S-Nitrosylation and Nitration, and Nitrosoglutathione Reductase in Arabidopsis thaliana Roots. Antioxidants 2020, 9, 795. [Google Scholar] [CrossRef]
- Vanholme, R.; Storme, V.; Vanholme, B.; Sundin, L.; Christensen, J.H.; Goeminne, G.; Halpin, C.; Rohde, A.; Morreel, K.; Boerjan, W. A Systems Biology View of Responses to Lignin Biosynthesis Perturbations in Arabidopsis. Plant Cell 2012, 24, 3506–3529. [Google Scholar] [CrossRef]
- Bhuiyan, N.H.; Selvaraj, G.; Wei, Y.; King, J. Role of Lignification in Plant Defense. Plant Signal. Behav. 2009, 4, 158–159. [Google Scholar] [CrossRef]
- Holbein, J.; Franke, R.B.; Marhavý, P.; Fujita, S.; Górecka, M.; Sobczak, M.; Geldner, N.; Schreiber, L.; Grundler, F.M.W.; Siddique, S. Root Endodermal Barrier System Contributes to Defence against Plant-Parasitic Cyst and Root-Knot Nematodes. Plant J. 2019, 100, 221–236. [Google Scholar] [PubMed]
- Golinowski, W.; Grundler, F.M.W.; Sobczak, M. Changes in the Structure of Arabidopsis thaliana during Female Development of the Plant-Parasitic nematode Heterodera schachtii. Protoplasma 1996, 194, 103–116. [Google Scholar]
- Bohlmann, H.; Sobczak, M. The Plant Cell Wall in the Feeding Sites of Cyst Nematodes. Front. Plant Sci. 2014, 5, 89. [Google Scholar]
- Karczmarek, A.; Fudali, S.; Lichocka, M.; Sobczak, M.; Kurek, W.; Janakowski, S.; Roosien, J.; Golinowski, W.; Bakker, J.; Goverse, A.; et al. Expression of Two Functionally Distinct Plant Endo-Beta-1,4-Glucanases Is Essential for the Compatible Interaction between Potato Cyst Nematode and Its Hosts. Mol. Plant Microbe Interact. 2008, 21, 791–798. [Google Scholar]
- Morant, A.V.; Jørgensen, K.; Jørgensen, C.; Paquette, S.M.; Sánchez-Pérez, R.; Møller, B.L.; Bak, S. Beta-Glucosidases as Detonators of Plant Chemical Defense. Phytochemistry 2008, 69, 1795–1813. [Google Scholar]
- Liu, W.; Ni, J.; Shah, F.A.; Ye, K.; Hu, H.; Wang, Q.; Wang, D.; Yao, Y.; Huang, S.; Hou, J.; et al. Genome-Wide Identification, Characterization and Expression Pattern Analysis of APYRASE Family Members in Response to Abiotic and Biotic Stresses in Wheat. PeerJ 2019, 7, e7622. [Google Scholar]
- Lim, M.H.; Wu, J.; Yao, J.; Gallardo, I.F.; Dugger, J.W.; Webb, L.J.; Huang, J.; Salmi, M.L.; Song, J.; Clark, G.; et al. Apyrase Suppression Raises Extracellular ATP Levels and Induces Gene Expression and Cell Wall Changes Characteristic of Stress Responses. Plant Physiol. 2014, 164, 2054–2067. [Google Scholar]
- Fu, Y.-F.; Zhang, Z.-W.; Yuan, S. Putative Connections Between Nitrate Reductase S-Nitrosylation and NO Synthesis Under Pathogen Attacks and Abiotic Stresses. Front. Plant Sci. 2018, 9, 474. [Google Scholar]
- Campbell, W.H. Nitrate Reductase Structure, Function and Regulation: Bridging the Gap between Biochemistry and Physiology. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 277–303. [Google Scholar]
- Labudda, M.; Różańska, E.; Muszyńska, E.; Marecka, D.; Głowienka, M.; Roliński, P.; Prabucka, B. Heterodera schachtii Infection Affects Nitrogen Metabolism in Arabidopsis thaliana. Plant Pathol. 2020, 69, 794–803. [Google Scholar]
- Baranowski, Ł.; Różańska, E.; Sańko-Sawczenko, I.; Matuszkiewicz, M.; Znojek, E.; Filipecki, M.; Grundler, F.M.W.; Sobczak, M. Arabidopsis Tonoplast Intrinsic Protein and Vacuolar H+-Adenosinetriphosphatase Reflect Vacuole Dynamics during Development of Syncytia Induced by the Beet Cyst Nematode Heterodera schachtii. Protoplasma 2019, 256, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Li, B.; Iwakawa, H.O.; Pan, Y.; Tang, X.; Ling-Hu, Q.; Liu, Y.; Sheng, S.; Feng, L.; Zhang, H.; et al. Plant 22-Nt siRNAs Mediate Translational Repression and Stress Adaptation. Nature 2020, 581, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Palanivelu, R.; Brass, L.; Edlund, A.F.; Preuss, D. Pollen Tube Growth and Guidance Is Regulated by POP2, an Arabidopsis Gene That Controls GABA Levels. Cell 2003, 114, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Seifi, H.S.; Curvers, K.; De Vleesschauwer, D.; Delaere, I.; Aziz, A.; Höfte, M. Concurrent Overactivation of the Cytosolic Glutamine Synthetase and the GABA Shunt in the ABA-Deficient sitiens Mutant of Tomato Leads to Resistance against Botrytis cinerea. New Phytol. 2013, 199, 490–504. [Google Scholar] [CrossRef]
- Wang, G.; Kong, J.; Cui, D.; Zhao, H.; Niu, Y.; Xu, M.; Jiang, G.; Zhao, Y.; Wang, W. Resistance against Ralstonia Solanacearum in Tomato Depends on the Methionine Cycle and the γ-Aminobutyric Acid Metabolic Pathway. Plant J. 2019, 97, 1032–1047. [Google Scholar] [CrossRef]
- Shelp, B.J.; Aghdam, M.S.; Flaherty, E.J. γ-Aminobutyrate (GABA) Regulated Plant Defense: Mechanisms and Opportunities. Plants (Basel) 2021, 10. [Google Scholar] [CrossRef]
- Reed, H.M.; Han, Z.; Schroeder, N.E. GABA Immunoreactivity and Pharmacological Effects Vary Among Stylet-Bearing Nematodes. J. Nematol. 2023, 55, 20230049. [Google Scholar] [CrossRef]
- Snider, J.; Thibault, G.; Houry, W.A. The AAA+ Superfamily of Functionally Diverse Proteins. Genome Biol. 2008, 9, 216. [Google Scholar] [CrossRef]
- Zhu, X.; Yin, J.; Liang, S.; Liang, R.; Zhou, X.; Chen, Z.; Zhao, W.; Wang, J.; Li, W.; He, M.; et al. The Multivesicular Bodies (MVBs)-Localized AAA ATPase LRD6-6 Inhibits Immunity and Cell Death Likely through Regulating MVBs-Mediated Vesicular Trafficking in Rice. PLoS Genet. 2016, 12, e1006311. [Google Scholar] [CrossRef]
- Zhang, B.; Van Aken, O.; Thatcher, L.; De Clercq, I.; Duncan, O.; Law, S.R.; Murcha, M.W.; van der Merwe, M.; Seifi, H.S.; Carrie, C.; et al. The Mitochondrial Outer Membrane AAA ATPase AtOM66 Affects Cell Death and Pathogen Resistance in Arabidopsis thaliana. Plant J. 2014, 80, 709–727. [Google Scholar] [CrossRef]
- Ogura, T.; Wilkinson, A.J. AAA+ Superfamily ATPases: Common Structure–Diverse Function. Genes. Cells 2001, 6, 575–597. [Google Scholar] [PubMed]
- Evans, K. Hatching of Potato Cyst Nematodes in Root Diffusates Collected from Twenty-Five Potato Cultivars. Crop Prot. 1983, 2, 97–103. [Google Scholar]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A Peptide Search Engine Integrated into the MaxQuant Environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef]
- Pant, B.D.; Musialak-Lange, M.; Nuc, P.; May, P.; Buhtz, A.; Kehr, J.; Walther, D.; Scheible, W.R. Identification of Nutrient-Responsive Arabidopsis and Rapeseed microRNAs by Comprehensive Real-Time Polymerase Chain Reaction Profiling and Small RNA Sequencing. Plant Physiol. 2009, 150, 1541–1555. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative Expression Software Tool (REST) for Group-Wise Comparison and Statistical Analysis of Relative Expression Results in Real-Time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Sijmons, P.C.; Grundler, F.M.W.; von Mende, N.; Burrows, P.R.; Wyss, U. Arabidopsis thaliana as a New Model Host for Plant-Parasitic Nematodes. Plant J. 1991, 1, 245–254. [Google Scholar]
- Fucile, G.; Di Biase, D.; Nahal, H.; La, G.; Khodabandeh, S.; Chen, Y.; Easley, K.; Christendat, D.; Kelley, L.; Provart, N.J. ePlant and the 3D Data Display Initiative: Integrative Systems Biology on the World Wide Web. PLoS One 2011, 6, e15237. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Tomato Gene ID | Arabidopsis Gene ID |
|---|---|---|
| Apyrase2 (Apy2) | Solyc12g098540.2.1 | AT5G18280 |
| Nitrate reductase 2 (NIA2) | Solyc11g013810.2.1 | AT1G37130 |
| AAA-ATPase 1 (AATP1) | Solyc05g007470.4.1 | AT5G40010 |
| Pyridoxal phosphate (PLP)-dependent transferases superfamily protein (POP) (GABA-T) | Solyc12g006470.2.1 | AT3G40010 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koter, M.D.; Żurczak, M.; Matuszkiewicz, M.; Święcicka, M.; Kotliński, M.; Barczak-Brzyżek, A.; Filipecki, M. Proteomic Dynamics in the Interaction of Susceptible and Resistant Tomato Cultivars and Potato Cyst Nematodes. Int. J. Mol. Sci. 2025, 26, 2823. https://doi.org/10.3390/ijms26062823
Koter MD, Żurczak M, Matuszkiewicz M, Święcicka M, Kotliński M, Barczak-Brzyżek A, Filipecki M. Proteomic Dynamics in the Interaction of Susceptible and Resistant Tomato Cultivars and Potato Cyst Nematodes. International Journal of Molecular Sciences. 2025; 26(6):2823. https://doi.org/10.3390/ijms26062823
Chicago/Turabian StyleKoter, Marek D., Marek Żurczak, Mateusz Matuszkiewicz, Magdalena Święcicka, Maciej Kotliński, Anna Barczak-Brzyżek, and Marcin Filipecki. 2025. "Proteomic Dynamics in the Interaction of Susceptible and Resistant Tomato Cultivars and Potato Cyst Nematodes" International Journal of Molecular Sciences 26, no. 6: 2823. https://doi.org/10.3390/ijms26062823
APA StyleKoter, M. D., Żurczak, M., Matuszkiewicz, M., Święcicka, M., Kotliński, M., Barczak-Brzyżek, A., & Filipecki, M. (2025). Proteomic Dynamics in the Interaction of Susceptible and Resistant Tomato Cultivars and Potato Cyst Nematodes. International Journal of Molecular Sciences, 26(6), 2823. https://doi.org/10.3390/ijms26062823