

Tryptophan Hydroxylase 1 Regulates Tryptophan and Its Metabolites


Abstract
1. Introduction
2. Results
2.1. Plasma Tryptophan and Its Metabolites in Tph1 Mutants and Wild-Type Mice
2.2. Brain Tryptophan and Its Metabolites in Tph1 Mutants and Wild-Type Mice
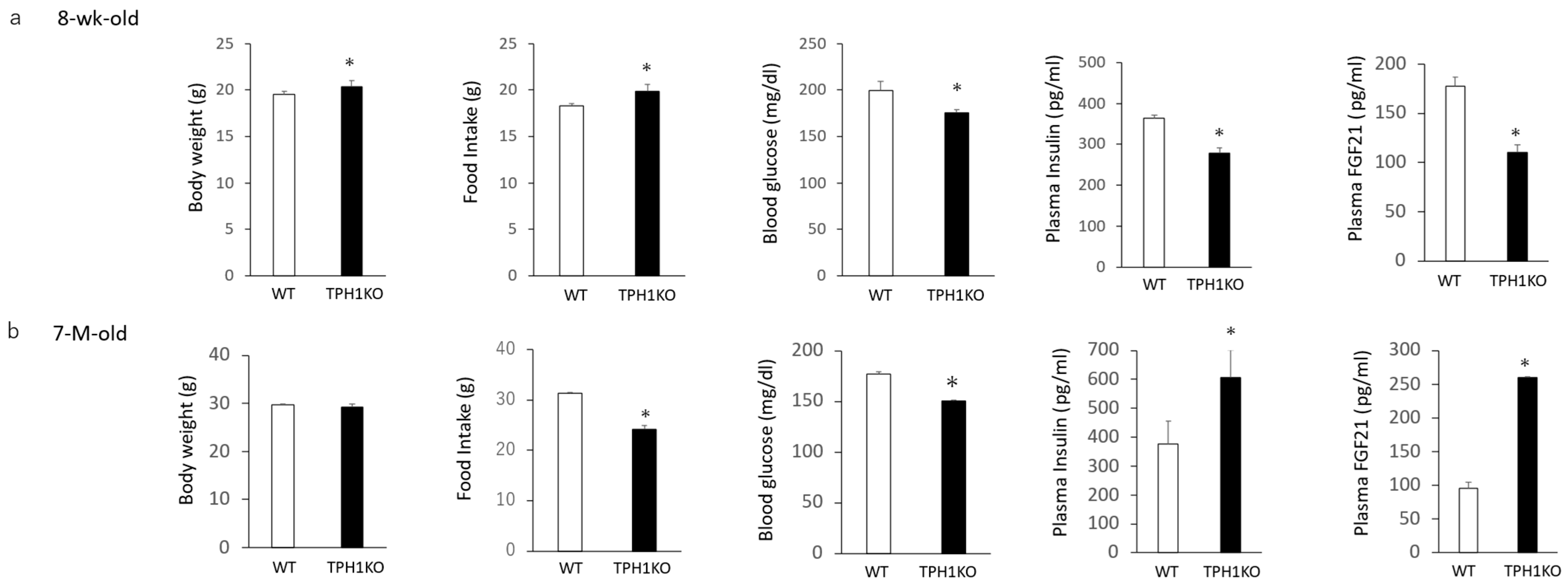
2.3. Body Weight, Food Intake, Blood Glucose and Plasma FGF21 and Insulin Levels in Tph1 Mutants and Wild Type Mice
3. Discussion
4. Materials and Methods
4.1. Tph1 Mutant Mice
4.2. Blood Chemistry
4.3. Tryptophan and Its Metabolites Analysis
4.4. Statistical Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Walther, D.J.; Peter, J.U.; Bashammakh, S.; Hörtnagl, H.; Voits, M.; Fink, H.; Bader, M. Synthesis of serotonin by a second tryptophan hydroxylase isoform. Science 2003, 299, 76. [Google Scholar] [CrossRef] [PubMed]
- Gutknecht, L.; Kriegebaum, C.; Waider, J.; Schmitt, A.; Lesch, K.P. Spatio-temporal expression of tryptophan hydroxylase isoforms in murine and human brain: Convergent data from Tph2 knockout mice. Eur. Neuropsychopharmacol. 2009, 19, 266–282. [Google Scholar] [CrossRef] [PubMed]
- Amireault, P.; Sibon, D.; Côté, F. Life without peripheral serotonin: Insights from tryptophan hydroxylase 1 knockout mice reveal the existence of paracrine/autocrine serotonergic networks. ACS Chem. Neurosci. 2013, 4, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, Y.; Lee, J.; Lee, J.Y.; Kim, H.; Lee, S.; Oh, C.-M. A Systems Biology Approach to Investigating the Interaction between Serotonin Synthesis by Tryptophan Hydroxylase and the Metabolic Homeostasis. Int. J. Mol. Sci. 2021, 22, 2452. [Google Scholar] [CrossRef] [PubMed]
- Paulmann, N.; Grohmann, M.; Voigt, J.P.; Bert, B.; Vowinckel, J.; Bader, M.; Skelin, M.; Jevsek, M.; Fink, H.; Rupnik, M.; et al. Intracellular serotonin modulates insulin secretion from pancreatic beta-cells by protein serotonylation. PLoS Biol. 2009, 7, e1000229. [Google Scholar]
- Martin, A.M.; Yabut, J.M.; Choo, J.M.; Page, A.J.; Sun, E.W.; Jessup, C.F.; Wesselingh, S.L.; Khan, W.I.; Rogers, G.B.; Steinberg, G.R.; et al. The gut microbiome regulates host glucose homeostasis via peripheral serotonin. Proc. Natl. Acad. Sci. USA 2019, 116, 19802–19804. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Namkung, J.; Hwang, I.; Kim, H.; Lim, A.; Park, H.J.; Lee, H.W.; Han, K.H.; Park, S.; Jeong, J.S.; et al. Serotonin signals through a gut-liver axis to regulate hepatic steatosis. Nat. Commun. 2018, 9, 4824. [Google Scholar] [CrossRef] [PubMed]
- Crane, J.D.; Palanivel, R.; Mottillo, E.P.; Bujak, A.L.; Wang, H.; Ford, R.J.; Collins, A.; Blümer, R.M.; Fullerton, M.D.; Yabut, J.M.; et al. Inhibiting peripheral serotonin synthesis reduces obesity and metabolic dysfunction by promoting brown adipose tissue thermogenesis. Nat. Med. 2015, 21, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Yabut, J.M.; Crane, J.D.; Green, A.E.; Keating, D.J.; Khan, W.I.; Steinberg, G.R. Emerging Roles for Serotonin in Regulating Metabolism: New Implications for an Ancient Molecule. Endocr. Rev. 2019, 40, 1092–1107. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.H.; Park, B.L.; Kim, H.; German, M.S.; Go, M.J.; Jung, H.S.; Koo, B.K.; Cho, Y.M.; Choi, S.H.; Cho, Y.S.; et al. Association of variations in TPH1 and HTR2B with gestational weight gain and measures of obesity. Obesity 2012, 20, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Genis-Mendoza, A.D.; Hernández-Díaz, Y.; González-Castro, T.B.; Tovilla-Zárate, C.A.; Castillo-Avila, R.G.; López-Narváez, M.L.; Ramos-Méndez, M.Á.; Nicolini, H. Association between TPH1 polymorphisms and the risk of suicide behavior: An updated meta-analysis of 18,398 individuals. Front. Psychiatry 2022, 13, 932135. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.C.; Bagade, S.; McQueen, M.B.; Ioannidis, J.P.; Kavvoura, F.K.; Khoury, M.J.; Tanzi, R.E.; Bertram, L. Systematic meta-analyses and field synopsis of genetic association studies in schizophrenia: The SzGene database. Nat. Genet. 2008, 40, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, K.; Kaji, T. Whey protein isolate inhibits hepatic FGF21 production, which precedes weight gain, hyperinsulinemia and hyperglycemia in mice fed a high-fat diet. Sci. Rep. 2020, 10, 15784. [Google Scholar] [CrossRef] [PubMed]
- Hajishafiee, M.; Ullrich, S.S.; Fitzgerald, P.C.; Horowitz, M.; Lange, K.; Poppitt, S.D.; Feinle-Bisset, C. Suppression of Energy Intake by Intragastric l-Tryptophan in Lean and Obese Men: Relations with Appetite Perceptions and Circulating Cholecystokinin and Tryptophan. J. Nutr. 2021, 151, 2932–2941. [Google Scholar] [CrossRef] [PubMed]
- Markan, K.R.; Naber, M.C.; Ameka, M.K.; Anderegg, M.D.; Mangelsdorf, D.J.; Kliewer, S.A.; Mohammadi, M.; Potthoff, M.J. Circulating FGF21 is liver derived and enhances glucose uptake during refeeding and overfeeding. Diabetes 2014, 63, 4057–4063. [Google Scholar] [CrossRef] [PubMed]
- Kharitonenkov, A.; DiMarchi, R. FGF21 revolutions: Recent advances illuminating FGF21 biology and medicinal properties. Trends Endocrinol. Metab. 2015, 26, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, S.; Kharitonenkov, A. FGF19 and FGF21: In NASH we trust. Mol. Metab. 2021, 46, 101152. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, K.; Kaji, T. The GLP-1 Receptor agonist liraglutide decreases primary bile acids and serotonin in the colon independently of feeding in mice. Int. J. Mol. Sci. 2024, 25, 7784. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| (a) | |||
| Plasma | 8-Week-Old WT | 8-Week-Old TPH1KO | % of WT |
| Trp | 21,133.3 ± 2168.9 | 12,025 ± 663.79 * | 57% |
| 5-HT | 256.66 ± 33.2 | 6.42 ± 0.46 * | 3% |
| 5-HIAA | 29.93 ± 1.22 | 15.45 ± 1.9 * | 52% |
| KYN | 115.36 ± 14.54 | 88.55 ± 7.47 * | 77% |
| XA | 155.4 ± 12.7 | 90.9 ± 4.2 * | 58% |
| IPA | 853.5 ± 35.5 | 665.75 ± 43.48 * | 78% |
| IAA | 160.33 ± 16.65 | 93.07 ± 17.66 * | 58% |
| KYNA | 9.13 ± 0.95 | 5.1 ± 0.39 * | 56% |
| Plasma | 7-Month-Old WT | 7-Month-Old TPH1KO | % of WT |
| Trp | 18,100 ± 2056.1 | 16,400 ± 1675.7 | 91% |
| 5-HT | 498 ± 141.2 | 3.54 ± 0.625 * | 1% |
| 5-HIAA | 21.3 ± 0.86 | 10.19 ± 0.26 * | 48% |
| KYN | 73.8 ± 2.26 | 76.78 ± 7.47 | 104% |
| XA | 113.9 ± 11.3 | 121.2 ± 17.95 | 106% |
| IPA | 753.3 ± 99.4 | 869.3 ± 114.08 | 115% |
| IAA | 71.1 ± 6.58 | 96.66 ± 7.38 * | 136% |
| KYNA | 4.54 ± 0.65 | 5.53 ± 1.05 | 122% |
| (b) | |||
| Brain | 8-Week-Old WT | 8-Week-Old TPH1KO | % of WT |
| Trp | 3710 ± 44.8 | 2755 ± 117 * | 74% |
| 5-HT | 444 ± 15.5 | 378 ± 9 * | 85% |
| 5-HIAA | 370 ± 13.8 | 316 ± 7 * | 85% |
| KYN | 28.77 ± 2.8 | 24.48 ± 1.2 | 85% |
| XA | 60.0 ± 1.3 | 46.0 ± 2.0 * | 77% |
| IPA | 7.96 ± 0.56 | 5.36 ± 0.07 * | 67% |
| IAA | 13 ± 0.3 | 11 ± 0.1 * | 85% |
| Brain | 7-Month-Old WT | 7-Month-Old TPH1KO | % of WT |
| Trp | 2613 ± 102 | 2530 ± 131.8 * | 97% |
| 5-HT | 423 ± 13.2 | 375 ± 10.3 * | 89% |
| 5-HIAA | 346 ± 3.4 | 306 ± 15.1 * | 88% |
| KYN | 15.53 ± 0.31 | 14.8 ± 0.04 * | 95% |
| XA | 45.0 ± 1.6 | 47.0 ± 2.2 | 104% |
| IPA | 8.52 ± 0.42 | 7.20 ± 1.22 * | 85% |
| IAA | 11 ± 0.3 | 11 ± 0.2 | 100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nonogaki, K.; Kaji, T. Tryptophan Hydroxylase 1 Regulates Tryptophan and Its Metabolites. Int. J. Mol. Sci. 2025, 26, 3978. https://doi.org/10.3390/ijms26093978
Nonogaki K, Kaji T. Tryptophan Hydroxylase 1 Regulates Tryptophan and Its Metabolites. International Journal of Molecular Sciences. 2025; 26(9):3978. https://doi.org/10.3390/ijms26093978
Chicago/Turabian StyleNonogaki, Katsunori, and Takao Kaji. 2025. "Tryptophan Hydroxylase 1 Regulates Tryptophan and Its Metabolites" International Journal of Molecular Sciences 26, no. 9: 3978. https://doi.org/10.3390/ijms26093978
APA StyleNonogaki, K., & Kaji, T. (2025). Tryptophan Hydroxylase 1 Regulates Tryptophan and Its Metabolites. International Journal of Molecular Sciences, 26(9), 3978. https://doi.org/10.3390/ijms26093978