The Relative Effects of Local and Landscape Characteristics of Hedgerows on Bats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling Design and Scale Approach
2.3. Hedgerow Characteristics on the Local and Landscape Scales
2.4. Bat Sampling
2.5. Mean Trait Community Index
2.5.1. Habitat-Specialization Index
2.5.2. Foraging-Strategy
2.6. Data Analysis
3. Results
3.1. Local Variables of Hedgerows
3.2. Landscape Variables of Hedgerows
3.3. Habitat Preferences of Species
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IUCN. The IUCN Red List of Threatened Species. 2017. Available online: http://www.iucnredlist.org/ (accessed on 1 January 2018).
- Bengtsson, J.; Ahnström, J.; Weibull, A. The effects of organic agriculture on biodiversity and abundance: A meta-analysis. J. Appl. Ecol. 2005, 42, 261–269. [Google Scholar] [CrossRef]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Estrada, A.; Coates-Estrada, R. Bats in continuous forest, forest fragments and in an agricultural mosaic habitat-island at Los Tuxtlas, Mexico. Biol. Conserv. 2002, 103, 237–245. [Google Scholar] [CrossRef]
- Gorresen, P.M.; Willig, M.R. Landscape responses of bats to habitat fragmentation in Atlantic forest of Paraguay. J. Mammal. 2004, 85, 688–697. [Google Scholar] [CrossRef]
- Kerbiriou, C.; Azam, C.; Touroult, J.; Marmet, J.; Julien, J.F.; Pellissier, V. Common bats are more abundant within Natura 2000 areas. Biol. Conserv. 2018, 217, 66–74. [Google Scholar] [CrossRef]
- Matson, P.A.; Parton, W.J.; Power, A.; Swift, M. Agricultural intensification and ecosystem properties. Science 1997, 277, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.; Gaulton, R.; Gerard, F.T.; Staley, J. The influence of hedgerow structural condition on wildlife habitat provision in farmed landscapes. Biol. Conserv. 2018, 220, 122–131. [Google Scholar] [CrossRef]
- Bellamy, C.; Scott, C.; Altringham, J. Multiscale, presence-only habitat suitability models: Fine-resolution maps for eight bat species. J. Appl. Ecol. 2013, 50, 892–901. [Google Scholar] [CrossRef]
- Lookingbill, T.R.; Elmore, A.J.; Engelhardt, K.A.; Churchill, J.B.; Gates, J.E.; Johnson, J.B. Influence of wetland networks on bat activity in mixed-use landscapes. Biol. Conserv. 2010, 143, 974–983. [Google Scholar] [CrossRef]
- Hale, J.D.; Fairbrass, A.J.; Matthews, T.J.; Sadler, J.P. Habitat composition and connectivity predicts bat presence and activity at foraging sites in a large UK conurbation. PLoS ONE 2012, 7, e33300. [Google Scholar] [CrossRef] [PubMed]
- Verboom, B.; Spoelstra, K. Effects of food abundance and wind on the use of tree lines by an insectivorous bat, Pipistrellus pipistrellus. Can. J. Zool. 1999, 77, 1393–1401. [Google Scholar] [CrossRef]
- Boughey, K.L.; Lake, I.R.; Haysom, K.A.; Dolman, P.M. Improving the biodiversity benefits of hedgerows: How physical characteristics and the proximity of foraging habitat affect the use of linear features by bats. Biol. Conserv. 2011, 144, 1790–1798. [Google Scholar] [CrossRef]
- Baudry, J.; Jouin, A. De la Haie aux Bocages. Organisation, Dynamique et Gestion; INRA: Paris, France, 2003; 435p. [Google Scholar]
- Wickramasinghe, L.P.; Harris, S.; Jones, G.; Vaughan Jennings, N. Abundance and species richness of nocturnal insects on organic and conventional farms: Effects of agricultural intensification on bat foraging. Conserv. Biol. 2004, 18, 1283–1292. [Google Scholar] [CrossRef]
- Dietz, C.; von Helversen, O.; Nill, D. L’encyclopédie des Chauves-Souris d’Europe et d’Afrique du Nord: Biologie, Caractéristiques, Protection; Delachaux et Niestlé: Paris, France, 2009; 400p. [Google Scholar]
- Froidevaux, J.S.P.; Boughey, K.L.; Barlow, K.E.; Jones, G. Factors driving population recovery of the greater horseshoe bat (Rhinolophus ferrumequinum) in the UK: Implications for conservation. Biodivers. Conserv. 2017, 26, 1–21. [Google Scholar] [CrossRef]
- Pinaud, D.; Claireau, F.; Leuchtmann, M.; Kerbiriou, C. Modelling landscape connectivity for greater horseshoe bat (Rhinolophus ferrumequinum) using an empirical quantification of resistance. J. Appl. Ecol. 2018, in press. [Google Scholar] [CrossRef]
- Walsh, A.L.; Harris, S. Factors determining the abundance of vespertilionid bats in Britain: Geographical, land class and local habitat relationships. J. Appl. Ecol. 1996, 33, 519–529. [Google Scholar] [CrossRef]
- Hein, C.D.; Castleberry, S.B.; Miller, K.V. Site-occupancy of bats in relation to forested corridors. For. Ecol. Manag. 2009, 257, 1200–1207. [Google Scholar] [CrossRef]
- Quinn, J.F.; Harrison, S.P. Effects of habitat fragmentation and isolation on species richness: Evidence from biogeographic patterns. Oecologia 1988, 75, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Boyce, M.S. Population viability analysis. Annu. Rev. Ecol. Syst. 1992, 23, 451–506. [Google Scholar] [CrossRef]
- Baudry, J.; Bunce, R.G.H.; Burel, F. Hedgerows: An international perspective on their origin, function and management. J. Environ. Manag. 2000, 60, 7–22. [Google Scholar] [CrossRef]
- Staley, J.T.; Bullock, J.M.; Baldock, K.C.R.; Redhead, J.W.; Hooftman, D.A.P.; Button, N.; Pywell, R.F. Changes in hedgerow floral diversity over 70 years in an English rural landscape, and the impacts of management. Biol. Conserv. 2013, 167, 97–105. [Google Scholar] [CrossRef]
- Morandin, L.A.; Kremen, C. Bee preference for native versus exotic plants in restored agricultural hedgerows. Restor. Ecol. 2013, 21, 26–32. [Google Scholar] [CrossRef]
- Morandin, L.A.; Long, R.F.; Kremen, C. Hedgerows enhance beneficial insects on adjacent tomato fields in an intensive agricultural landscape. Agric. Ecosyst. Environ. 2014, 189, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Sklenicka, P.; Molnarova, K.; Brabec, E.; Kumble, P.; Pittnerova, B.; Pixova, K.; Salek, M. Remnants of medieval field patterns in the Czech Republic: Analysis of driving forces behind their disappearance with special attention to the role of hedgerows. Agric. Ecosyst. Environ. 2009, 129, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Staley, J.T.; Sparks, T.H.; Croxton, P.J.; Baldock, K.C.R.; Heard, M.S.; Hulmes, S.; Hulmes, L.; Peyton, J.; Amy, S.R.; Pywell, R.F. Long-term effects of hedgerow management policies on resource provision for wildlife. Biol. Conserv. 2012, 145, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Carey, P.D.; Wallis, S.; Chamberlain, P.M.; Cooper, A.; Emmett, B.A.; Maskell, L.C.; McCann, T.; Murphy, J.; Norton, L.R.; Reynolds, B.; et al. Chapter 5, Boundary and Linear Features Broad Habitat. In Countryside Survey: UK Results from 2007; Carey, P.D., Wallis, S., Chamberlain, P.M., Cooper, A., Emmett, B.A., Maskell, L.C., McCann, T., Murphy, J., Norton, L.R., Reynolds, B., et al., Eds.; Centre for Ecology and Hydrology, Natural Environment Research Council: Swindon, UK, 2008. [Google Scholar]
- Thenail, C.; Baudry, J. Variation of farm spatial land use pattern according to the structure of the hedgerow network (bocage) landscape: A case study in northeast Brittany. Agric. Ecosyst. Environ. 2004, 101, 53–72. [Google Scholar] [CrossRef]
- Fahrig, L.; Baudry, J.; Brotons, L.; Burel, F.G.; Crist, T.O.; Fuller, R.J.; Sirami, C.; Siriwardena, G.M.; Martin, J.L. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Lett. 2011, 14, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Briers, R.A. Incorporating connectivity into reserve selection procedures. Biol. Conserv. 2002, 103, 77–83. [Google Scholar] [CrossRef]
- Lawton, J.H.; Brotherton, P.N.M.; Brown, V.K.; Elphick, C.; Fitter, A.H.; Forshaw, J.; Haddow, R.W.; Hilborne, S.; Leafe, R.N.; Mace, G.M.; et al. Making Space for Nature: A Review of England’s Wildlife Sites and Ecological Network; DEFRA: London, UK, 2010. [Google Scholar]
- Maes, J.; Barbosa, A.; Baranzelli, C.; Zulian, G.; Batista e Silva, F.; Vandecasteele, I.; Hiederer, R.; Liquete, C.; Paracchini, M.L.; Mubareka, S.; et al. More green infrastructure is required to maintain ecosystem services under current trends in land-use change in Europe. Landsc. Ecol. 2015, 30, 517–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, H.; Gourmelon, F.; Rouan, M.; Le Viol, I.; Kerbiriou, C. The contribution of agent-based simulations to conservation management on a Natura 2000 site. J. Environ. Manag. 2016, 168, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleijn, D.; Sutherland, W.J. How effective are European agri-environment schemes in conserving and promoting biodiversity? J. Appl. Ecol. 2003, 40, 947–969. [Google Scholar] [CrossRef] [Green Version]
- Kleijn, D.; Baquero, R.A.; Clough, Y.; Diaz, M.; De Estaban, J.; Fernandez, F.; Gabriel, D.; Herzog, F.; Holzschuh, A.; Jöhl, R.; et al. Mixed biodiversity benefits of agri-environment schemes in five European countries. Ecol. Lett. 2006, 9, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Wittingham, M.J. The future of agri-environment schemes: Biodiversity gains and ecosystem service delivery? J. Appl. Ecol. 2011, 48, 509–513. [Google Scholar] [CrossRef]
- Fuentes-Montemayor, E.; Goulson, D.; Park, K.J. Pipistrelle bats and their prey do not benefit from four widely applied agri-environment management prescriptions. Biol. Conserv. 2011, 144, 2233–2246. [Google Scholar] [CrossRef]
- Davidson-Watts, I.; Jones, G. Differences in foraging behaviour between Pipistrellus pipistrellus (Schreber, 1774) and Pipistrellus pygmaeus (Leach, 1825): Foraging behaviour in cryptic bat species. J. Zool. 2005, 268, 55–62. [Google Scholar] [CrossRef]
- Perez-Jorda, J.L.; Ibañez, C. Preliminary results on activity rhythms and space use obtained by radio-tracking a colony of Eptesicus serotinus. Myotis 1991, 29, 61e66. [Google Scholar]
- Frey-Ehrenbold, A.; Bontadina, F.; Arlettaz, R.; Obrist, M.K. Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matrices. J. Appl. Ecol. 2013, 50, 252–261. [Google Scholar] [CrossRef]
- Kelm, D.H.; Lenski, J.; Kelm, V.; Toelch, U.; Dziock, F. Seasonal bat activity in relation to distance to hedgerows in an agricultural landscape in central Europe and implications for wind energy development. Acta Chiropterologica 2014, 16, 65–73. [Google Scholar] [CrossRef]
- Lewis, T. The Distribution of Flying Insects near a Low Hedgerow. J. Appl. Ecol. 1969, 6, 443–452. [Google Scholar] [CrossRef]
- Barataud, M. Acoustic Ecology of European Bats, Species Identification, Study of Their Habitat and Foraging Behavior; Inventaires et biodiversité Series; Muséum National d’Histoire Naturelle: Paris, France, 2015; 352p. [Google Scholar]
- Kerbiriou, C.; Bas, Y.; Le Viol, I.; Lorrilliere, R.; Mougnot, J.; Julien, J.-F. Potentiality of the bat pass duration measure for studies dealing with bat activity. Bioacoustic 2018, in press. [Google Scholar] [CrossRef]
- Desrochers, A.; Renaud, C.; Hochachka, W.M.; Cadman, M. Area-sensitivity by forest songbirds: Theoretical and practical implications of scale-dependency. Ecography 2010, 33, 921–931. [Google Scholar] [CrossRef]
- Penone, C.; Kerbiriou, C.; Julien, J.F.; Julliard, R.; Machon, N.; Le Viol, I. Urbanisation effect on Orthoptera: Which scale matters? Insect Conserv. Divers. 2013, 6, 319–332. [Google Scholar] [CrossRef]
- Van Dijk, G. The status of semi-natural grasslands in Europe. Conserv. Low Land Dry Grassl. Birds Eur. 1991, 15–36. Available online: http://mandadb.hu/common/file-servlet/document/659220/default/doc_url/The_status_of_semi_natural_grasslands_in_europe.pdf (accessed on 15 April 2018).
- Geffray, O. Le bocage en vallée de la Loire: Mise à jour cartographique et estimation des capacités en bois-énergie. Conservatoire Régional des Rives de la Loire et de ses Affluents (CORELA), Rapport d’étude. 2010. Available online: http://www.geopal.org/accueil/geoservice (accessed on 15 April 2018).
- Stone, E.; Zeale, M.R.K.; Newson, S.E.; Browne, W.J.; Harris, S.; Jones, G. Managing Conflict between Bats and Humans: The Response of Soprano Pipistrelles (Pipistrellus pygmaeus) to Exclusion from Roosts in Houses. PLoS ONE 2015, 10, e0131825. [Google Scholar] [CrossRef] [PubMed]
- Hillen, J.; Kiefer, A.; Veith, M. Interannual Fidelity to Roosting Habitat and Flight Paths by Female Western Barbastelle Bats. Acta Chiropterol. 2010, 12, 187–195. [Google Scholar] [CrossRef]
- Flaquer, C.; Puig-Montserrat, X.; Goiti, U.; Vidal, F.; Curcó, A.; Russo, D. Habitat selection in Nathusius’ pipistrelle (Pipistrellus nathusii): the importance of wetlands. Acta Chiropterol. 2009, 11, 149–155. [Google Scholar] [CrossRef]
- Robinson, M.F.; Stebbings, R.E. Home range and habitat use by the serotine bat, Eptesicus serotinus, in England. J. Zool. 1997, 243, 117–136. [Google Scholar] [CrossRef]
- Roche, N.; Catto, C.; Langton, S.; Aughney, T.; Russ, J. Development of a Car-Based Bat Monitoring Protocol for the Republic of Ireland; Irish Wildlife Manuals, No. 19; National Parks and Wildlife Service, Department of Environment, Heritage and Local Government: Dublin, Ireland, 2005.
- Météo France. French National Meteorological Service. 2012. Available online: www.meteofrance.com (accessed on 1 August 2012).
- Burt, J. Syrinx a Software for Real Time Spectrographic Recording, Analysis and Playback of Sound. 2006. Available online: http://www.syrinxpc.com (accessed on 15 April 2018).
- Lacoeuilhe, A.; Machon, N.; Le Bocq, A.; Julien, J.F.; Kerbiriou, K. The influence of low intensities of light pollution on bat communities in a semi-natural context. PLoS ONE 2014, 9, e103042. [Google Scholar] [CrossRef] [PubMed]
- Lacoeuilhe, A.; Machon, N.; Julien, J.F.; Kerbiriou, C. Effects of hedgerows on bats and bush crickets at different spatial scales. Acta Oecol. 2016, 71, 61–72. [Google Scholar] [CrossRef]
- Julliard, R.; Clavel, J.; Devictor, V.; Jiguet, F.; Couvet, D. Spatial segregation of specialists and generalists in bird communities. Ecol. Lett. 2006, 9, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Schnitzler, H.U.; Moss, C.F.; Denzinger, A. From spatial orientation to food acquisition in echolocating bats. Trends Ecol. Evol. 2003, 18, 386–394. [Google Scholar] [CrossRef]
- Audet, D. Foraging behavior and habitat use by a gleaning bat, Myotis myotis (Chiroptera: Vespertilonidae). J. Mammal. 1990, 71, 420–427. [Google Scholar] [CrossRef]
- Swift, S.M.; Racey, P.A. Gleaning as a foraging strategy in Natterer’s bat Myotis nattereri. Behav. Ecol. Sociobiol. 2002, 52, 408–416. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 2 July 2018).
- Fournier, D.A.; Skaug, H.J.; Ancheta, J.; Ianelli, J.; Magnusson, A.; Maunder, M.; Nielsen, A.; Sibert, J. AD Model Builder: Using automatic differentiation for statistical inference of highly parameterized complex nonlinear models. Optim. Methods Softw. 2012, 27, 233–249. [Google Scholar] [CrossRef] [Green Version]
- Skaug, H.; Fournier, D.; Bolker, B.; Magnusson, A.; Nielsen, A. Generalized Linear Mixed Models Using ‘AD Model Builder’. R Package Version 0.8.3.2.2015. Available online: http://glmmadmb.r-forge.r-project.org/repos/ (accessed on 30 June 2013).
- Hayes, J.P. Temporal variation in activity of bats and the design of echolocation-monitoring studies. J. Mammal. 1997, 78, 514–524. [Google Scholar] [CrossRef]
- O’Donnell, C.F.J. Conservation status and causes of decline of the threatened New Zealand long-tailed bat Chalinolobus tuberculatus (Chiroptera: Vespertilionidae). Mamm. Rev. 2000, 30, 89–106. [Google Scholar] [CrossRef]
- Azam, C.; Kerbiriou, C.; Vernet, A.; Julien, J.F.; Bas, Y.; Plichard, L.; Maratrat, J.; Le Viol, I. Is part-night lighting an effective measure to limit the impacts of artificial lighting on bats? Glob. Chang. Boil. 2015, 21, 4333–4341. [Google Scholar] [CrossRef] [PubMed]
- Ciechanowski, M.; Zajac, T.; Bilas, A.; Dunajski, R. Spatiotemporal variation in activity of bat species differing in hunting tactics: Effects of weather moonlight, food abundance, and structural clutter. Can. J. Zool. 2007, 85, 1249–1263. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Statistics for Biology and Health; Springer: New York, NY, USA, 2009; 574p. [Google Scholar]
- Schielzeth, H. Simple means to improve the interpretability of regression coefficients. Methods Ecol. Evol. 2010, 1, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Dormann, C.F.; McPherson, J.M.; Araújo, M.B.; Bivand, R.; Bolliger, J.; Carl, G.; Davies, R.D.; Hirzel, A.; Jetz, W.; Daniel Kissling, W.; et al. Methods to account for spatial autocorrelation in the analysis of species distributional data: A review. Ecography 2007, 30, 609–628. [Google Scholar] [CrossRef]
- Bivand, R.S.; Hauke, J.; Kossowski, T. Computing the Jacobian in Gaussian spatial autoregressive models: An illustrated comparison of available methods. Geogr. Anal. 2013, 45, 150–179. [Google Scholar] [CrossRef]
- Crawley, M.J. The R Book; Wiley-Blackwell: Chicago, IL, USA, 2009. [Google Scholar]
- Fox, J.; Monette, G. Generalized collinearity diagnostics. J. Am. Stat. Assoc. 1992, 87, 178–183. [Google Scholar] [CrossRef]
- Heiberger, R.M.; Holland, B. Multiple Comparisons. In Statistical Analysis and Data Display; Springer: New York, NY, USA, 2004; pp. 155–185. [Google Scholar] [CrossRef]
- Chatterjee, S.; Hadi, A.S. Regression Analysis by Example; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Inference—A Practical Information-Theorical Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Burnham, K.P.; Anderson, D.R.; Huyvaert, K.P. AIC model selection and multimodel inference in behavioral ecology: Some background, observations, and comparisons. Behav. Ecol. Sociobiol. 2011, 65, 23–35. [Google Scholar] [CrossRef]
- Verboom, B.; Huitema, H. The importance of linear landscape elements for the pipistrelle Pipistrellus pipistrellus and the serotine bat Eptesicus serotinus. Landsc. Ecol. 1997, 12, 117–125. [Google Scholar] [CrossRef]
- Dietz, M.; Pir, J.B.; Hillen, J. Does the survival of greater horseshoe bats and Geoffroy’s bats in Western Europe depend on traditional cultural landscapes? Biodivers. Conserv. 2013, 22, 3007–3025. [Google Scholar] [CrossRef]
- Zeale, M.R.; Davidson-Watts, I.; Jones, G. Home range use and habitat selection by barbastelle bats (Barbastella barbastellus): Implications for conservation. J. Mammal. 2012, 93, 1110–1118. [Google Scholar] [CrossRef]
- Arthur, L.; Lemaire, M.; Dufrêne, L.; Le Viol, I.; Julien, J.F.; Kerbiriou, C. Understanding bat-habitat associations and the effects of monitoring on long-term roost success using a volunteer dataset. Acta Chiropterol. 2014, 16, 397–411. [Google Scholar] [CrossRef]
- Racey, P.A.; Swift, S.M. Feeding Ecology of Pipistrellus pipistrellus (Chiroptera: Vespertilionidae) during Pregnancy and Lactation. I. Foraging Behaviour. J. Anim. Ecol. 1985, 54, 205–215. [Google Scholar] [CrossRef]
- Razgour, O.; Hanmer, J.; Jones, G. Using multi-scale modelling to predict habitat suitability for species of conservation concern: The grey long-eared bat as a case study. Biol. Conserv. 2011, 144, 2922–2930. [Google Scholar] [CrossRef] [Green Version]
- Tillon, L. Utilisation des gîtes et des terrains de chasse par les Chiroptères forestiers, propositions de gestion conservatoire. Ph.D. Thesis, Université Toulouse III Paul Sabatier, Toulouse, France, 2015. [Google Scholar]
- Larrieu, L.; Cabanettes, A. Species, live status, and diameter are important tree features for diversity and abundance of tree microhabitats in subnatural montane beech-fir forest. Can. J. For. Res. 2012, 42, 1433–1445. [Google Scholar] [CrossRef]
- Lewis, T. Patterns of distribution of insects near a windbreak of tall trees. Ann. Appl. Biol. 1970, 65, 213–220. [Google Scholar] [CrossRef]
- Lewis, T.; Dibley, G.C. Air movement near windbreaks and a hypothesis of the mechanism of the accumulation of airborne insects. Ann. Appl. Biol. 1970, 66, 477–484. [Google Scholar] [CrossRef]
- Pasek, J.E. Influence of wind and windbreaks on local dispersal of insects. Agric. Ecosyst. Environ. 1988, 22–23, 539–554. [Google Scholar] [CrossRef]
- Holderied, M.W.; von Helversen, O. Echolocation range and wingbeat period match in aerial-hawking bats. Proc. Biol. Sci. 2003, 270, 2293–2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merckx, T.; Feber, R.; Mclaughlan, C.; Bourn, N.A.D.; Parsons, M.S.; Townsend, M.C.; Riordan, P.; Macdonald, D.W. Shelter benefits less mobile moth species: The field-scale effect of hedgerow trees. Agric. Ecosyst. Environ. 2010, 138, 147–151. [Google Scholar] [CrossRef]
- Arlettaz, R. Feeding behaviour and foraging strategy of free-living mouse-eared bats, Myotis myotis and Myotis blythii. Anim. Behav. 1996, 51, 1–11. [Google Scholar] [CrossRef]
- Zahn, A.; Rottenwallner, A.; Güttinger, R. Population density of the greater mouse-eared bat (Myotis myotis), local diet composition and availability of foraging habitats. J. Zool. 2006, 269, 486–493. [Google Scholar] [CrossRef]
- Pereira, M.J.; Rebelo, H.; Rainho, A.; Palmeirim, J.M. Prey selection by Myotis myotis (Vespertilionidae) in a Mediterranean region. Acta Chiropterol. 2002, 4, 183–193. [Google Scholar] [CrossRef]
- Arlettaz, R.; Godat, S.; Meyer, H. Competition for food by expanding pipistrelle bat populations (Pipistrellus pipistrellus) might contribute to the decline of lesser horseshoe bats (Rhinolophus hipposideros). Biol. Conserv. 2000, 93, 55–60. [Google Scholar] [CrossRef]
- Sierro, A.; Arlettaz, R. Barbastelle bats (Barbastella spp.) specialize in the predation of mooths: Implications for foraging tactics and conservation. Acta Oecol. 1997, 18, 91–106. [Google Scholar] [CrossRef]
- Beck, A. Fecal analysis of European bat species. Myotis 1995, 32–33, 109–119. [Google Scholar]
- Barr, C.J.; Parr, T.W. Hedgerows: Linking ecological research and countryside policy. In Hedgerow Management and Nature Conservation; Watt, T.A., Buckley, G.P., Eds.; Wye College Press, Wye College, University of London: London, UK, 1994; pp. 119–136. [Google Scholar]
- Department for Environment Food and Rural Affairs (DEFRA). Trends, Long Term Survival and Ecological Values of Hedgerow Trees: Development of Populations Models to Inform Strategy; Report to the UK Government Department for the Environment, Food and Rural Affairs; Forest Research, Forestry Commission: London, UK, 2010.
- Pointereau, P. Les haies évolution du linéaire en France depuis quarante ans. Courrier de l’environnement de l’INRA 2002, 46, 69–73. [Google Scholar]
- Charrier, S.; Petit, S.; Burel, F. Movements of Abax parallelepipedus (Coleoptera, Carabidae) in woody habitats of a hedgerow network landscape: A radiotracing study. Agric. Ecosyst. Environ. 1997, 61, 133–144. [Google Scholar] [CrossRef]
- Staley, J.T.; Botham, M.S.; Chapman, R.E.; Amy, S.R.; Heard, M.S.; Hulmes, L.; Savage, J.; Pywell, R.F. Little and late: How reduced hedgerow cutting can benefit Lepidoptera. Agric. Ecosyst. Environ. 2016, 224, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Riffell, S.; Verschuyl, J.; Miller, D.; Wigley, T.B. Biofuel harvests, coarse woody debris, and biodiversity—A meta-analysis. For. Ecol. Manag. 2011, 261, 878–887. [Google Scholar] [CrossRef]


{kind=link}
| Bat Species | Total Number of Calls | Average Number of Calls per Sample (±SE) | Habitat Specialization Index (SSI) | Foraging Type |
|---|---|---|---|---|
| Barbastella barbastellus | 304 | 2.980 ± 2.036 | 3.6 | NA |
| Eptesicus serotinus | 817 | 8.010 ± 2.336 | 1.5 | Hawking |
| Myotis spp. (mainly daubentonii) | 179 | 1.755 ± 0.763 | 3.2 | Gleaning |
| Nyctalus leisleri | 327 | 3.206 ± 1.118 | 0.9 | Hawking |
| Nyctalus noctula | 274 | 2.686 ± 1.054 | 1.6 | Hawking |
| Pipistrellus kuhlii | 8600 | 84.314 ± 15.202 | 0.7 | Hawking |
| Pipistrellus nathusii | 7464 | 73.176 ± 40.149 | 3.1 | Hawking |
| Pipistrellus pipistrellus | 17,090 | 167.549 ± 75.354 | 0.5 | Hawking |
| Plecotus spp. (mainly austriacus) | 208 | 2.039 ± 0.968 | 1.4 | Gleaning |
| Species and Communities | Distribution in the Model | Foraging Strategy *** | Local Hedgerow Variables | Landscape Hedgerow Variables | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Length50m | Shrub50m | Wooded50m | 3strat50m | Dist_hedgerow | Density1000m | Woodproduct1000m | Diversity1000m | |||||||||||
| Estimated ± se | P-Value | Estimated ± se | P-Value | Estimated ± se | P-Value | Estimated ± se | P-Value | Estimated ± se | P-Value | Estimated ± se | P-Value | Estimated ± se | P-Value | Estimated ± se | P-Value | |||
| B. barbastellus | Binomial | NA | 1.04 ± 0.50 | 0.04 * | −1.28 ± 0.90 | 0.16 | 1.65 ± 0.86 | 0.05 | ||||||||||
| E. serotinus | NegBin | Hawking | 0.26 ± 0.12 | 0.03 * | −0.68 ± 0.33 | 0.04 * | −1.08 ± 0.42 | 0.01 ** | 1.81 ± 0.44 | 4.3 × 10−5 *** | ||||||||
| P. kuhlii | NegBin | Hawking | 0.19 ± 0.10 | 0.07 | −0.34 ± 0.17 | 0.05 * | −0.52 ± 0.16 | 0.1 × 10−2 ** | ||||||||||
| P. nathusii | NegBin | Hawking | 0.27 ± 0.16 | 0.09 | −0.51 ± 0.19 | 0.01 ** | −0.29 ± 0.16 | 0.07 | 0.32 ± 0.16 | 0.04 * | −0.26 ± 0.16 | 0.10 | ||||||
| P. pipistrellus | NegBin | Hawking | 0.44 ± 0.08 | 2.1 × 10−7 *** | 0.37 ± 0.09 | 3.7 × 10−5 *** | −0.27 ± 0.11 | 0.02 * | −0.54 ± 0.12 | 1.0 × 10−5 *** | ||||||||
| N. leisleri | NegBin | Hawking | −1.23 ± 0.36 | 0.001 *** | −0.80 ± 0.40 | 0.04 * | −0.41 ± 0.20 | 0.04 * | −0.90 ± 0.42 | 0.03 * | 0.96 ± 0.34 | 0.0052 ** | ||||||
| N. noctula | Binomial | Hawking | 0.68 ± 0.37 | 0.07 | ||||||||||||||
| Myotis spp. * | Binomial | Gleaning | / | |||||||||||||||
| Plecotus spp. ** | Binomial | Gleaning | 1.03 ± 0.59 | 0.08 | 1.20 ± 0.45 | 0.01 ** | −6.90 ± 6.51 | 0.29 | −0.98 ± 0.65 | 0.13 | ||||||||
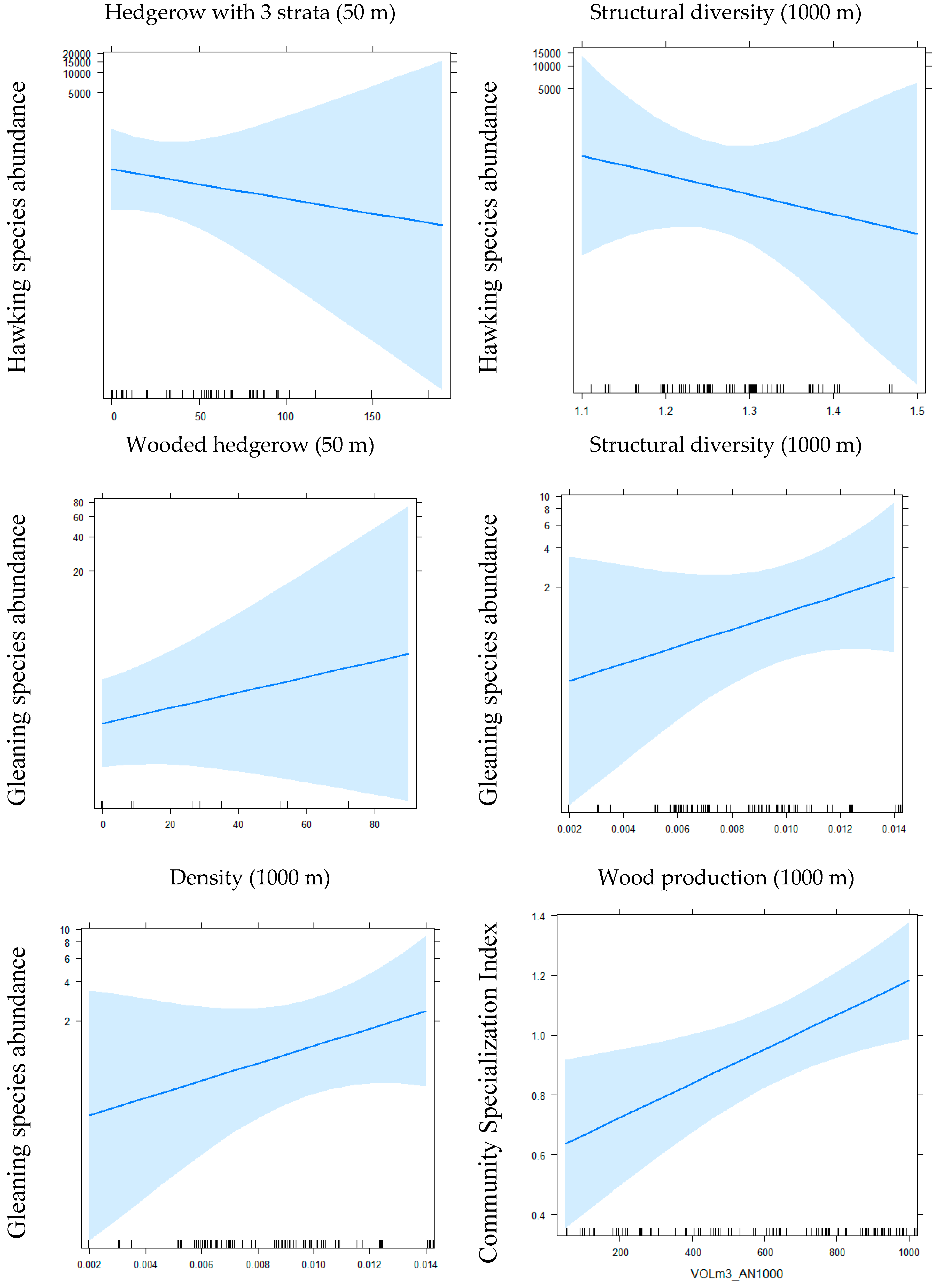
| Hawking species | NegBin | / | 0.32 ± 0.11 | 0.3 × 10−2 ** | 0.15 ± 0.07 | 0.04 * | −0.34 ± 0.12 | 0.004 ** | −0.40 ± 0.10 | 4.3 × 10−5 *** | ||||||||
| Gleaning species | NegBin | / | 0.59 ± 0.22 | 0.01 ** | −4.29 ± 1.94 | 0.03 * | −0.36 ± 0.16 | 0.02 * | 4.56 ± 2.64 | 0.08 | −11.87 ± 6.18 | 0.06 | 7.86 ± 3.71 | 0.034 * | 7.61 ± 3.91 | 0.05 | 1.12 ± 0.18 | 1.3 × 10−9 *** |
| CSI | Gaussian | / | 0.18 ± 0.05 | 0.004 | ||||||||||||||
| Species richness | NegBin | / | 0.12 ± 0.06 | 0.06 | −0.16 ± 0.075 | 0.03 * | −0.18 ± 0.07 | 0.0140 * | ||||||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lacoeuilhe, A.; Machon, N.; Julien, J.-F.; Kerbiriou, C. The Relative Effects of Local and Landscape Characteristics of Hedgerows on Bats. Diversity 2018, 10, 72. https://doi.org/10.3390/d10030072
Lacoeuilhe A, Machon N, Julien J-F, Kerbiriou C. The Relative Effects of Local and Landscape Characteristics of Hedgerows on Bats. Diversity. 2018; 10(3):72. https://doi.org/10.3390/d10030072
Chicago/Turabian StyleLacoeuilhe, Aurélie, Nathalie Machon, Jean-François Julien, and Christian Kerbiriou. 2018. "The Relative Effects of Local and Landscape Characteristics of Hedgerows on Bats" Diversity 10, no. 3: 72. https://doi.org/10.3390/d10030072
APA StyleLacoeuilhe, A., Machon, N., Julien, J.-F., & Kerbiriou, C. (2018). The Relative Effects of Local and Landscape Characteristics of Hedgerows on Bats. Diversity, 10(3), 72. https://doi.org/10.3390/d10030072